- Структура и функции липопротеинов

Содержание
- 2. доц. Болотникова О.И. * Пространственная организация липопротеинов плазмы крови
- 3. доц. Болотникова О.И. * Структура и функция ЛП липопротетны ЛПНП ЛПВП Белки- 2% Фосфолипиды- 3% ТАГ-
- 4. доц. Болотникова О.И. * Сравнительный анализ ЛП
- 5. доц. Болотникова О.И. * Апобелки липопротеинов Интегральные: В-48 В-100 Периферические: рецепторы - апоЕ (богат аргини-ном), взаимодействует
- 6. доц. Болотникова О.И. * Метаболизм ХМ мишени ХМ – мышцы и жировая ткань ТАГ- липиды расщепляет
- 7. доц. Болотникова О.И. * Функция ЛПОНП белки, синтезируемые в шеро-ховатом эндоплазматическом ретикулуме [1] и аппарате Голь-джи
- 8. доц. Болотникова О.И. * Транспорт холестерола в составе ЛПНП осуществляется к тканям и клеткам -мишеням, обладающим
- 9. доц. Болотникова О.И. * Функционирование рецепторов ЛПНП при взаимодействии рецептора и ЛПНП формируется ямка, выстланная белком
- 10. доц. Болотникова О.И. * Регуляция биосинтеза рецепторов ЛПНП в клетках-мишенях клетки тканей-мишеней содержат огромное количество рецепторов
- 11. доц. Болотникова О.И. * Биологическая функция ЛПВП поставляют апопротеины (апо-СII, апо-А-I и апо-Е) ХМ и ЛПОНП
- 12. доц. Болотникова О.И. * Метаболизм ЛПВП гепатоцит клетка ткани-мишени ЛП-липаза плазма крови ЛХАТ депонирование ТАГ-липидов синтез
- 13. доц. Болотникова О.И. * Общая характеристика дислипопротеинемий - изменения обмена ЛП крови, при-водящие к нарушениям обмена
- 14. НАРУШЕНИЯ ОБМЕНА ЛИПИДОВ
- 15. доц. Болотникова О.И. * Катаболизм гликосфинголипидов
- 16. доц. Болотникова О.И. * Сфинголипидозы (лизосомные болезни) нарушения катаболизма гликосфинголипидов обусловлены генетическими дефектами лизосомных ферментов приводят
- 17. доц. Болотникова О.И. * Болезнь Гоше наиболее типичными проявлениями служат увеличение размеров селезенки и печени (гепатоспленомегалия),
- 18. доц. Болотникова О.И. * Причины болезни Гоше накопление клеток, перегруженных «шлаками», и нарушение их функ-ции увеличение
- 19. доц. Болотникова О.И. * Причины ожирения алиментарный дисбаланс - избыточная калорийность питания по сравнению с расходом
- 20. доц. Болотникова О.И. * Гиперхолестеролемия - Превышение нормальной концентра-ции холестерола в крови (200 50г/дл или 5,2
- 21. доц. Болотникова О.И. * Молекулярные основы атеросклероза процесс начинается с повреждения эндотелия сосудов за счет измененной
- 22. доц. Болотникова О.И. * Последствия атеросклероза накопление «пенистых клеток» обуславливает повреждение эндотелия сосудов активируется процесс агрегации
- 24. Скачать презентацию
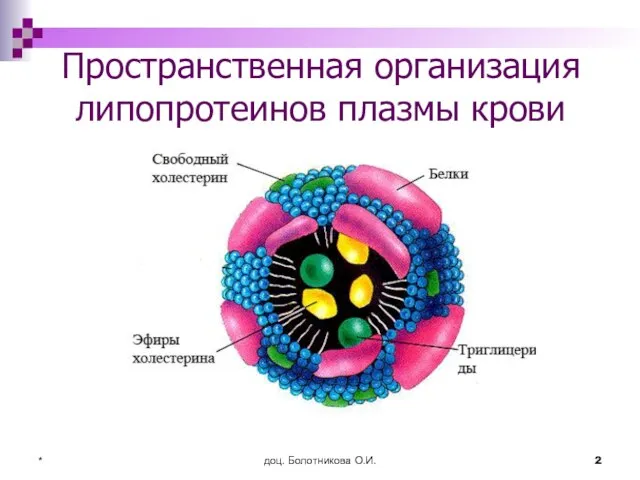
Слайд 3доц. Болотникова О.И.
*
Структура и функция ЛП
липопротетны
ЛПНП
ЛПВП
Белки- 2%
Фосфолипиды- 3%
ТАГ- 85%
транспорт липидов из кишечника
доц. Болотникова О.И.
*
Структура и функция ЛП
липопротетны
ЛПНП
ЛПВП
Белки- 2%
Фосфолипиды- 3%
ТАГ- 85%
транспорт липидов из кишечника
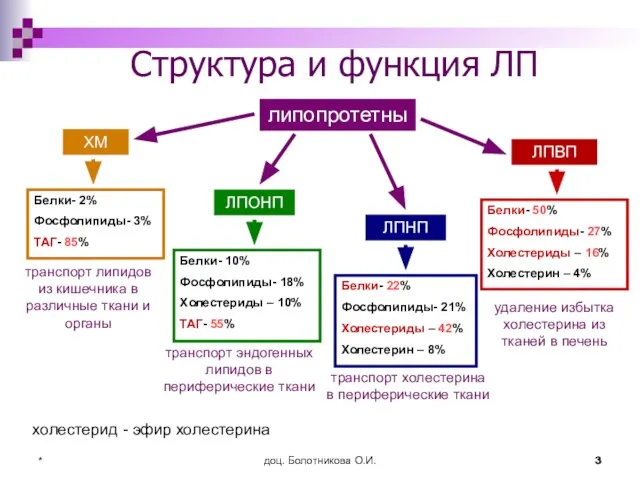
Слайд 4доц. Болотникова О.И.
*
Сравнительный анализ ЛП
доц. Болотникова О.И.
*
Сравнительный анализ ЛП
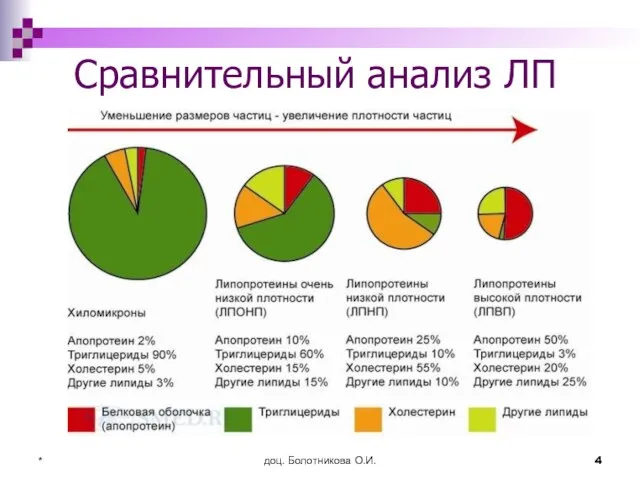
Слайд 5доц. Болотникова О.И.
*
Апобелки липопротеинов
Интегральные:
В-48
В-100
Периферические:
рецепторы - апоЕ (богат аргини-ном), взаимодействует с ЛПНП
активаторы ферментов:
доц. Болотникова О.И.
*
Апобелки липопротеинов
Интегральные:
В-48
В-100
Периферические:
рецепторы - апоЕ (богат аргини-ном), взаимодействует с ЛПНП
активаторы ферментов:
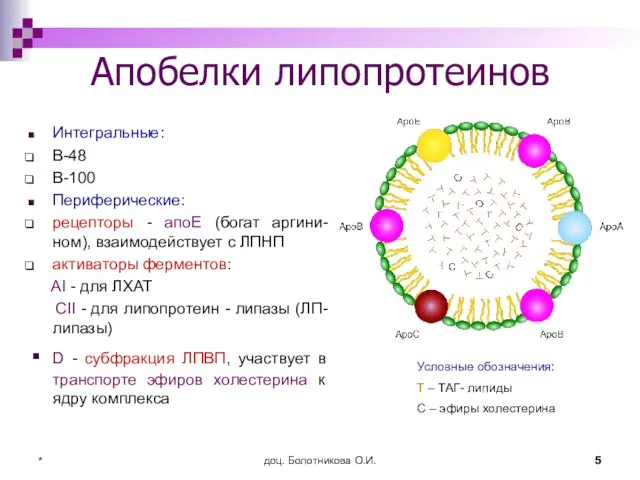
Слайд 6доц. Болотникова О.И.
*
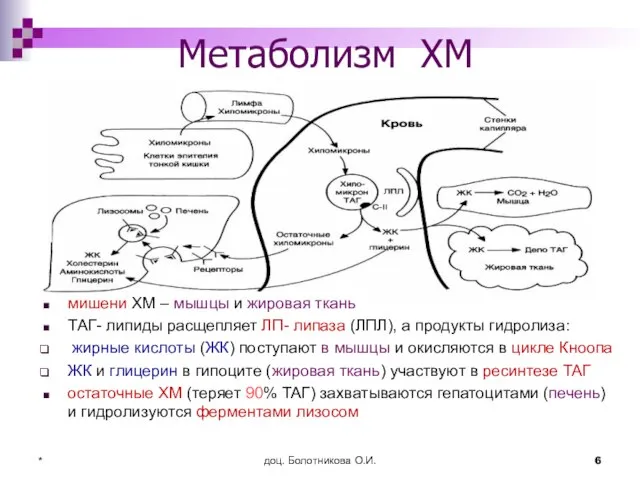
Метаболизм ХМ
мишени ХМ – мышцы и жировая ткань
ТАГ- липиды расщепляет
доц. Болотникова О.И.
*
Метаболизм ХМ
мишени ХМ – мышцы и жировая ткань
ТАГ- липиды расщепляет
Слайд 7доц. Болотникова О.И.
*
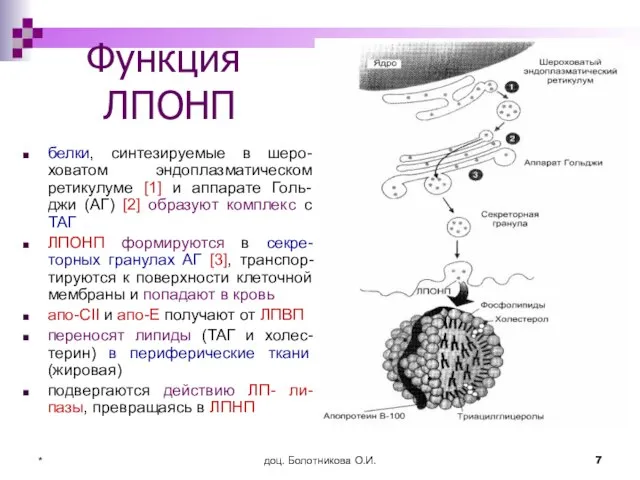
Функция
ЛПОНП
белки, синтезируемые в шеро-ховатом эндоплазматическом ретикулуме [1] и аппарате
доц. Болотникова О.И.
*
Функция
ЛПОНП
белки, синтезируемые в шеро-ховатом эндоплазматическом ретикулуме [1] и аппарате
Слайд 8доц. Болотникова О.И.
*
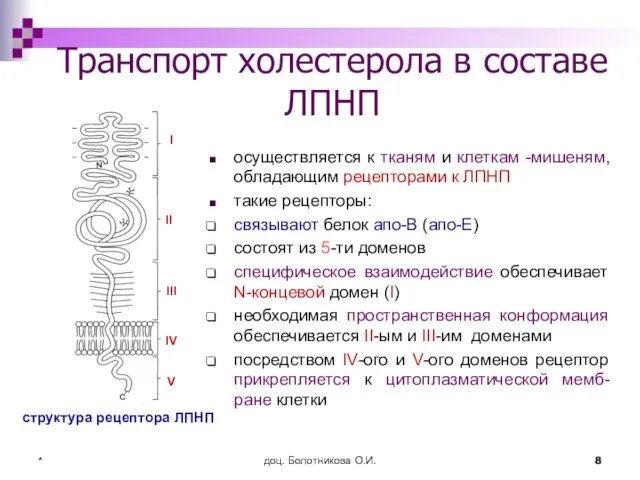
Транспорт холестерола в составе ЛПНП
осуществляется к тканям и клеткам -мишеням,
доц. Болотникова О.И.
*
Транспорт холестерола в составе ЛПНП
осуществляется к тканям и клеткам -мишеням,
Слайд 9доц. Болотникова О.И.
*
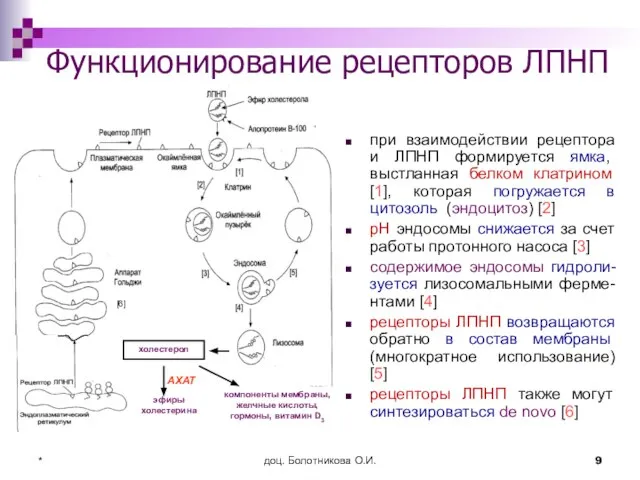
Функционирование рецепторов ЛПНП
при взаимодействии рецептора и ЛПНП формируется ямка, выстланная
доц. Болотникова О.И.
*
Функционирование рецепторов ЛПНП
при взаимодействии рецептора и ЛПНП формируется ямка, выстланная
Слайд 10доц. Болотникова О.И.
*
Регуляция биосинтеза рецепторов ЛПНП в клетках-мишенях
клетки тканей-мишеней содержат огромное количество
доц. Болотникова О.И.
*
Регуляция биосинтеза рецепторов ЛПНП в клетках-мишенях
клетки тканей-мишеней содержат огромное количество

Слайд 11доц. Болотникова О.И.
*
Биологическая функция ЛПВП
поставляют апопротеины (апо-СII, апо-А-I и апо-Е) ХМ и
доц. Болотникова О.И.
*
Биологическая функция ЛПВП
поставляют апопротеины (апо-СII, апо-А-I и апо-Е) ХМ и

Слайд 12доц. Болотникова О.И.
*
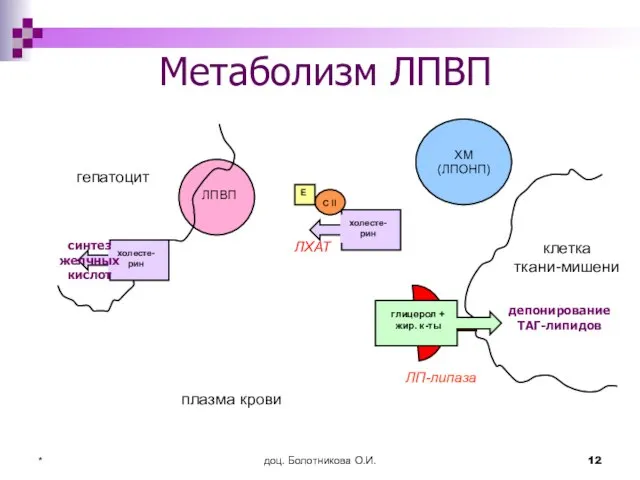
Метаболизм ЛПВП
гепатоцит
клетка ткани-мишени
ЛП-липаза
плазма крови
ЛХАТ
депонирование ТАГ-липидов
синтез желчных кислот
доц. Болотникова О.И.
*
Метаболизм ЛПВП
гепатоцит
клетка ткани-мишени
ЛП-липаза
плазма крови
ЛХАТ
депонирование ТАГ-липидов
синтез желчных кислот
Слайд 13доц. Болотникова О.И.
*
Общая характеристика дислипопротеинемий
- изменения обмена ЛП крови, при-водящие к нарушениям
доц. Болотникова О.И.
*
Общая характеристика дислипопротеинемий
- изменения обмена ЛП крови, при-водящие к нарушениям

Слайд 14НАРУШЕНИЯ ОБМЕНА ЛИПИДОВ
НАРУШЕНИЯ ОБМЕНА ЛИПИДОВ

Слайд 15доц. Болотникова О.И.
*
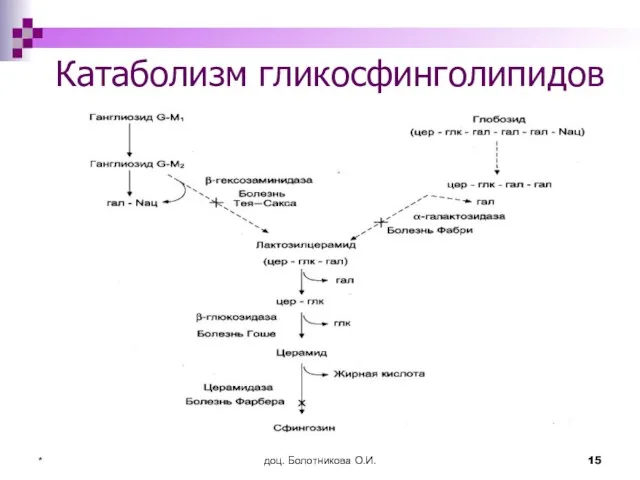
Катаболизм гликосфинголипидов
доц. Болотникова О.И.
*
Катаболизм гликосфинголипидов
Слайд 16доц. Болотникова О.И.
*
Сфинголипидозы (лизосомные болезни)
нарушения катаболизма гликосфинголипидов
обусловлены генетическими дефектами лизосомных ферментов
приводят
доц. Болотникова О.И.
*
Сфинголипидозы (лизосомные болезни)
нарушения катаболизма гликосфинголипидов
обусловлены генетическими дефектами лизосомных ферментов
приводят

Слайд 17доц. Болотникова О.И.
*
Болезнь Гоше
наиболее типичными проявлениями служат увеличение размеров селезенки и печени
доц. Болотникова О.И.
*
Болезнь Гоше
наиболее типичными проявлениями служат увеличение размеров селезенки и печени

Слайд 18доц. Болотникова О.И.
*
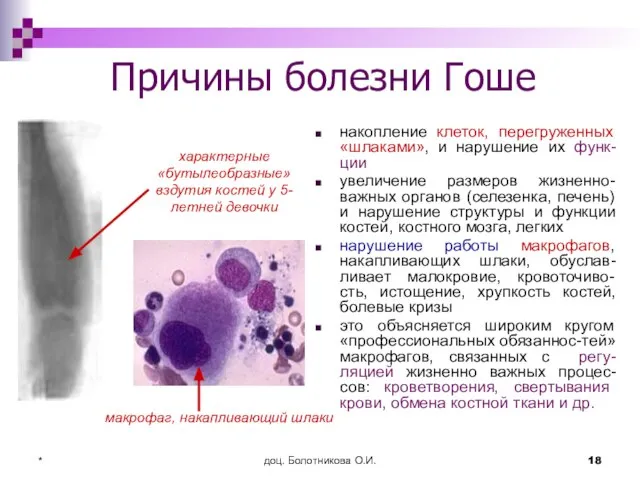
Причины болезни Гоше
накопление клеток, перегруженных «шлаками», и нарушение их
доц. Болотникова О.И.
*
Причины болезни Гоше
накопление клеток, перегруженных «шлаками», и нарушение их
Слайд 19доц. Болотникова О.И.
*
Причины ожирения
алиментарный дисбаланс - избыточная калорийность питания по сравнению с
доц. Болотникова О.И.
*
Причины ожирения
алиментарный дисбаланс - избыточная калорийность питания по сравнению с

Слайд 20доц. Болотникова О.И.
*
Гиперхолестеролемия
- Превышение нормальной концентра-ции холестерола в крови (200 50г/дл
доц. Болотникова О.И.
*
Гиперхолестеролемия
- Превышение нормальной концентра-ции холестерола в крови (200 50г/дл

Слайд 21доц. Болотникова О.И.
*
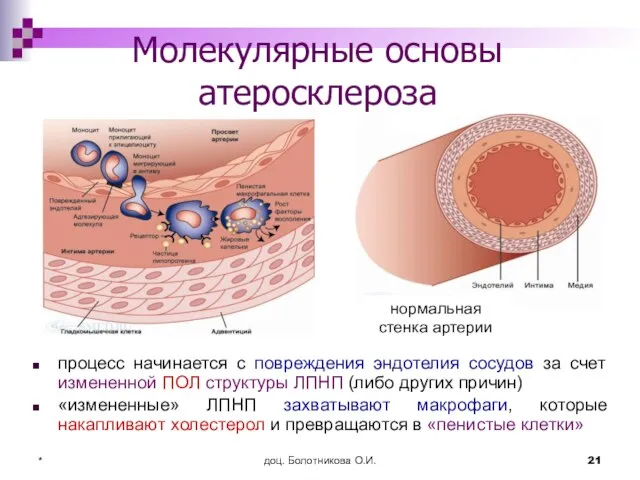
Молекулярные основы атеросклероза
процесс начинается с повреждения эндотелия сосудов за счет
доц. Болотникова О.И.
*
Молекулярные основы атеросклероза
процесс начинается с повреждения эндотелия сосудов за счет
Слайд 22доц. Болотникова О.И.
*
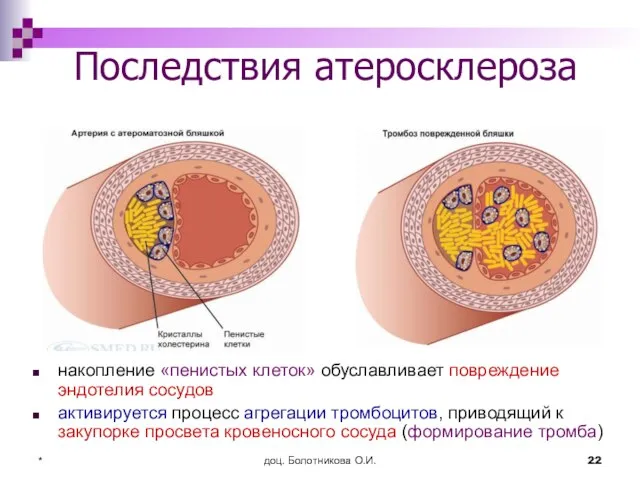
Последствия атеросклероза
накопление «пенистых клеток» обуславливает повреждение эндотелия сосудов
активируется процесс агрегации
доц. Болотникова О.И.
*
Последствия атеросклероза
накопление «пенистых клеток» обуславливает повреждение эндотелия сосудов
активируется процесс агрегации
 Black English (Афроамериканский английский )
Black English (Афроамериканский английский ) Презентация на тему Корень. Родственные слова. Два признака родственных слов
Презентация на тему Корень. Родственные слова. Два признака родственных слов Презентация на тему Русские народные танцы (8 класс)
Презентация на тему Русские народные танцы (8 класс) Первые блюда. Супы
Первые блюда. Супы «Загрязнение атмосферного воздуха посёлка «Магистральный» выбросами автотранспорта и влияние отработанных газовна здоровье
«Загрязнение атмосферного воздуха посёлка «Магистральный» выбросами автотранспорта и влияние отработанных газовна здоровье  Создание рабочих мест
Создание рабочих мест С:ЕНКО MEN
С:ЕНКО MEN Традиции модернизма в искусстве 2-й пол. ХХ века
Традиции модернизма в искусстве 2-й пол. ХХ века Многогранники в живой природе
Многогранники в живой природе Sale 30% Bestia
Sale 30% Bestia ВЫСТУПЛЕНИЕ ПЕРВОГО ВИЦЕ-ПРЕЗИДЕНТА ОАО «РЖД»В.Н.МОРОЗОВА на VII Международной конференции «Рынок транспортных услуг: взаимодейс
ВЫСТУПЛЕНИЕ ПЕРВОГО ВИЦЕ-ПРЕЗИДЕНТА ОАО «РЖД»В.Н.МОРОЗОВА на VII Международной конференции «Рынок транспортных услуг: взаимодейс Эффективный нападающий удар в волейболе
Эффективный нападающий удар в волейболе Print from the folder
Print from the folder Шаровая Молния.
Шаровая Молния. МОУ Голицынская СОШ №1 Презентация на тему : «Александр Невский – Патриот Земли Русской»
МОУ Голицынская СОШ №1 Презентация на тему : «Александр Невский – Патриот Земли Русской» Отделение дополнительного образования детей ГОУ школы №20
Отделение дополнительного образования детей ГОУ школы №20 Это мы, здравствуйте! 6 «Б» класс
Это мы, здравствуйте! 6 «Б» класс Потребности и способности
Потребности и способности a4c02c41e02991ba9d4f7fdd7cfefa1c
a4c02c41e02991ba9d4f7fdd7cfefa1c Собрание родителей и обучающихся 11 классов
Собрание родителей и обучающихся 11 классов Анализ и оценка организационных структур управления
Анализ и оценка организационных структур управления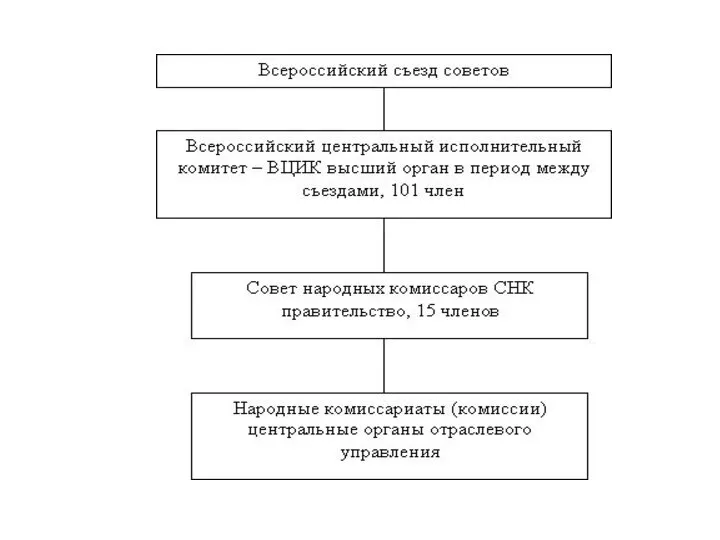 Органы государственной власти по Конституции 1918 г
Органы государственной власти по Конституции 1918 г Азбука потребителя
Азбука потребителя Совещание с грузоотправителями Калининградской железной дороги
Совещание с грузоотправителями Калининградской железной дороги Орудия труда
Орудия труда Полупроводниковые приборы
Полупроводниковые приборы Презентация на тему Гора Рашмор
Презентация на тему Гора Рашмор Программы государственной поддержки малого предпринимательства на селе в Республике Карелия
Программы государственной поддержки малого предпринимательства на селе в Республике Карелия