ВЛИЯНИЕ ИММОБИЛИЗАЦИИ И ФИЗИЧЕСКИХ УСЛОВИЙ КУЛЬТИВИРОВАНИЯ НА СОДЕРЖАНИЕ ГИДРОКСИКОРИЧНЫХ КИСЛОТ В КЛЕТОЧНЫХ КУЛЬТУРАХ
- ВЛИЯНИЕ ИММОБИЛИЗАЦИИ И ФИЗИЧЕСКИХ УСЛОВИЙ КУЛЬТИВИРОВАНИЯ НА СОДЕРЖАНИЕ ГИДРОКСИКОРИЧНЫХ КИСЛОТ В КЛЕТОЧНЫХ КУЛЬТУРАХ

Содержание
- 2. Содержание Актуальность Цель работы Объекты исследований Результаты Выводы
- 3. Актуальность Растения являются уникальными источниками многих биологически активных соединений. Несмотря на успехи в области применения синтетических
- 4. В связи с этим большой практический интерес представляют приемы получения фитомассы, основанные на культивировании в искусственных
- 5. Цель работы Исследование характера влияния иммобилизации и физических условий культивирования (температуры, света, слабого электрического тока) на
- 6. Объекты исследований каллусная и суспензионная культуры эхинацеи пурпурной
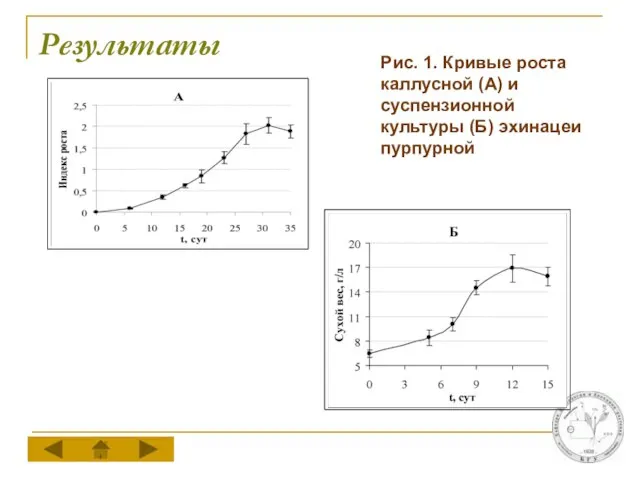
- 7. Результаты Рис. 1. Кривые роста каллусной (А) и суспензионной культуры (Б) эхинацеи пурпурной
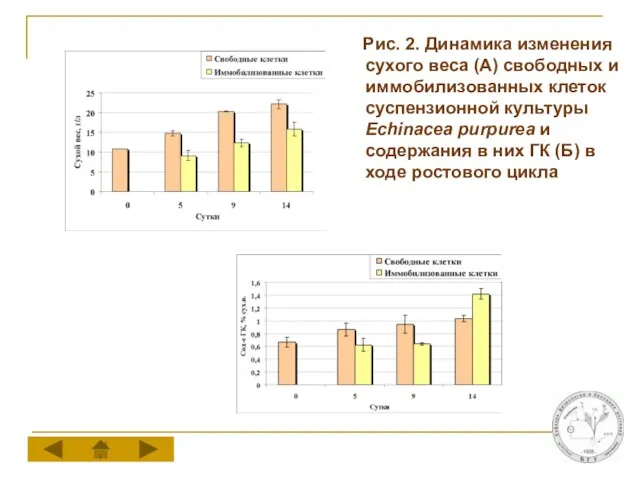
- 8. Рис. 2. Динамика изменения сухого веса (А) свободных и иммобилизованных клеток суспензионной культуры Echinacea purpurea и
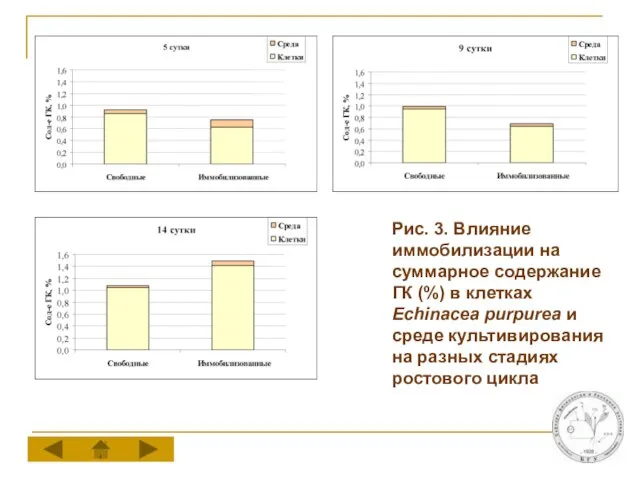
- 9. Рис. 3. Влияние иммобилизации на суммарное содержание ГК (%) в клетках Echinacea purpurea и среде культивирования
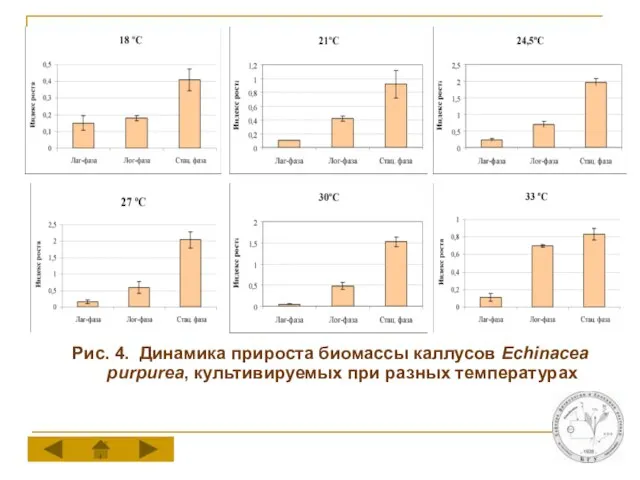
- 10. Рис. 4. Динамика прироста биомассы каллусов Echinacea purpurea, культивируемых при разных температурах
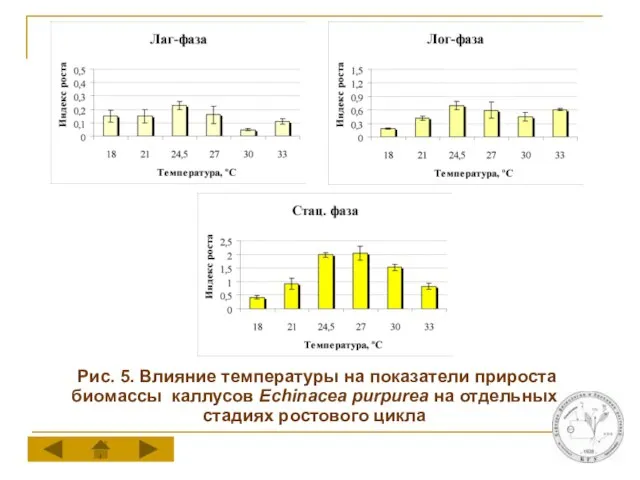
- 11. Рис. 5. Влияние температуры на показатели прироста биомассы каллусов Echinacea purpurea на отдельных стадиях ростового цикла
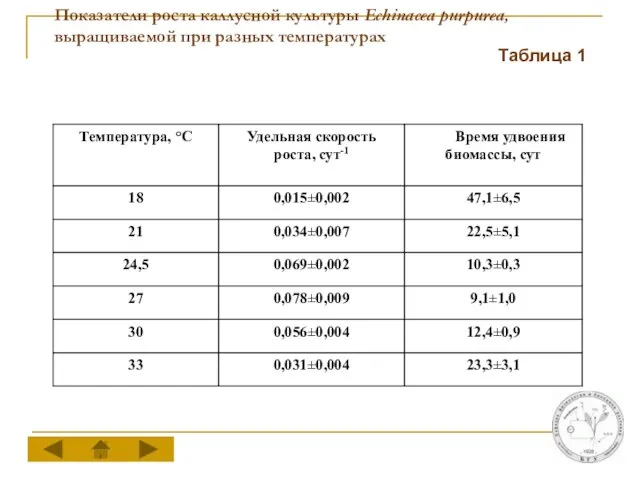
- 12. Показатели роста каллусной культуры Echinacea purpurea, выращиваемой при разных температурах Таблица 1
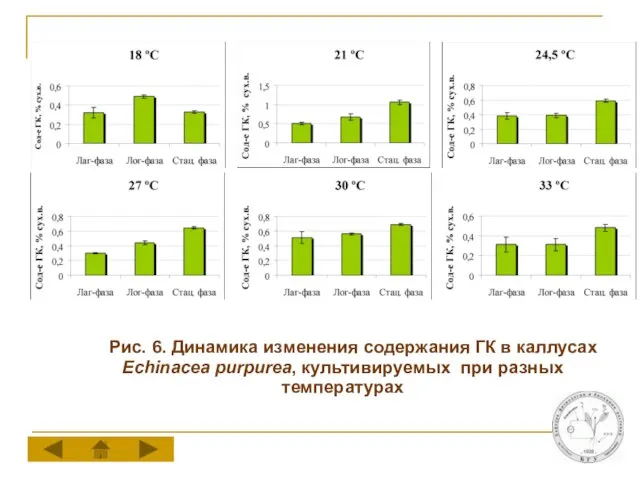
- 13. Рис. 6. Динамика изменения содержания ГК в каллусах Echinacea purpurea, культивируемых при разных температурах
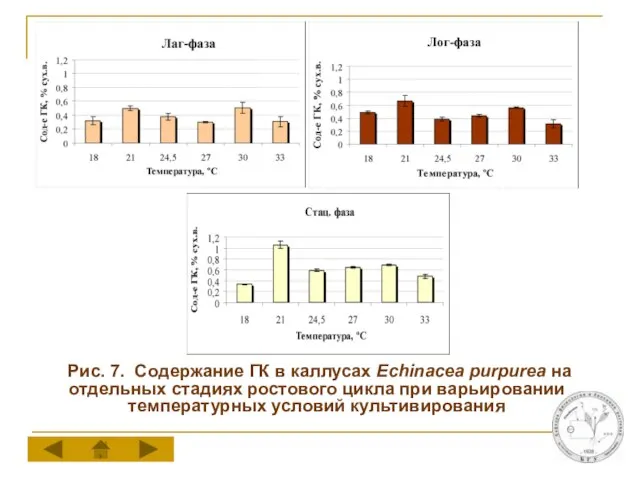
- 14. Рис. 7. Содержание ГК в каллусах Echinacea purpurea на отдельных стадиях ростового цикла при варьировании температурных
- 15. Рис. 8. Температурная зависимость продукции ГК каллусной культурой Echinacea purpurea
- 16. Рис. 9. Влияние условий освещения на индекс роста каллусов Echinacea purpurea (А) и содержание в них
- 17. Рис. 10. Влияние света на продукцию ГК каллусной культурой Echinacea purpurea
- 18. Рис. 11. Динамика изменения содержания фотосинтетических пигментов в каллусной культуре Echinacea purpurea в ходе ростового цикла
- 19. Рис. 12. Влияние слабого электрического тока (1,5 мкА) на содержание ГК в каллусах Echinacea purpurea, культивируемых
- 20. Выводы Включение клеток суспензионной культуры эхинацеи пурпурной в полисахаридный носитель (Са-альгинатый гель) вызывает стимуляцию накопления ГК
- 21. Высокая скорость прироста биомассы клеток при температуре 27°С компенсирует более низкие уровни накопления ГК, и максимальная
- 23. Скачать презентацию

Слайд 3Актуальность
Растения являются уникальными источниками многих биологически активных соединений. Несмотря на успехи
Актуальность
Растения являются уникальными источниками многих биологически активных соединений. Несмотря на успехи

Слайд 4 В связи с этим большой практический интерес представляют приемы получения фитомассы,
В связи с этим большой практический интерес представляют приемы получения фитомассы,

Слайд 5Цель работы
Исследование характера влияния иммобилизации и физических условий культивирования (температуры, света,
Цель работы
Исследование характера влияния иммобилизации и физических условий культивирования (температуры, света,

Слайд 6Объекты исследований
каллусная и суспензионная культуры эхинацеи пурпурной
Объекты исследований
каллусная и суспензионная культуры эхинацеи пурпурной

Слайд 7Результаты
Рис. 1. Кривые роста каллусной (А) и суспензионной культуры (Б) эхинацеи
Результаты
Рис. 1. Кривые роста каллусной (А) и суспензионной культуры (Б) эхинацеи
Слайд 8 Рис. 2. Динамика изменения сухого веса (А) свободных и иммобилизованных клеток
Рис. 2. Динамика изменения сухого веса (А) свободных и иммобилизованных клеток
Слайд 9Рис. 3. Влияние иммобилизации на суммарное содержание ГК (%) в клетках Echinacea
Рис. 3. Влияние иммобилизации на суммарное содержание ГК (%) в клетках Echinacea
Слайд 10Рис. 4. Динамика прироста биомассы каллусов Echinacea purpurea, культивируемых при разных температурах
Рис. 4. Динамика прироста биомассы каллусов Echinacea purpurea, культивируемых при разных температурах
Слайд 11 Рис. 5. Влияние температуры на показатели прироста биомассы каллусов Echinacea purpurea
Рис. 5. Влияние температуры на показатели прироста биомассы каллусов Echinacea purpurea
Слайд 12
Показатели роста каллусной культуры Echinacea purpurea, выращиваемой при разных температурах
Таблица 1
Показатели роста каллусной культуры Echinacea purpurea, выращиваемой при разных температурах
Таблица 1
Слайд 13 Рис. 6. Динамика изменения содержания ГК в каллусах Echinacea purpurea, культивируемых
Рис. 6. Динамика изменения содержания ГК в каллусах Echinacea purpurea, культивируемых
Слайд 14 Рис. 7. Содержание ГК в каллусах Echinacea purpurea на отдельных стадиях
Рис. 7. Содержание ГК в каллусах Echinacea purpurea на отдельных стадиях
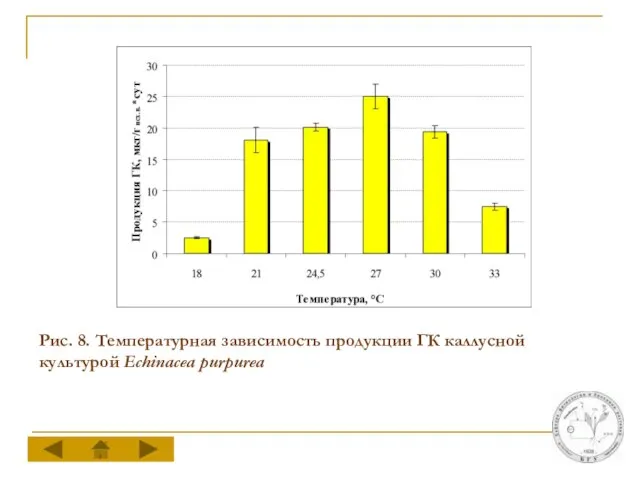
Слайд 15Рис. 8. Температурная зависимость продукции ГК каллусной культурой Echinacea purpurea
Рис. 8. Температурная зависимость продукции ГК каллусной культурой Echinacea purpurea
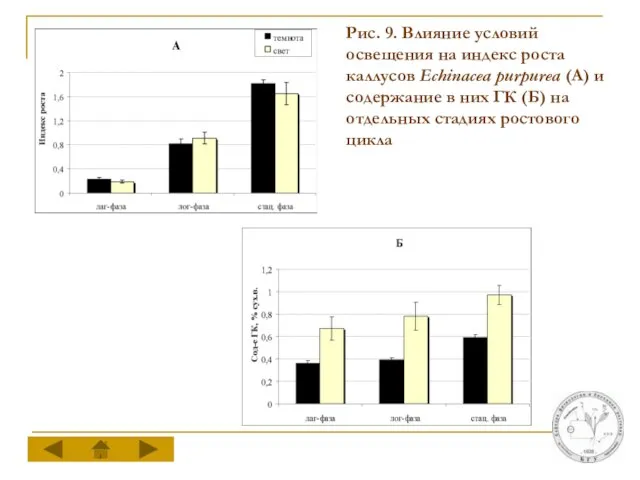
Слайд 16Рис. 9. Влияние условий освещения на индекс роста каллусов Echinacea purpurea (А)
Рис. 9. Влияние условий освещения на индекс роста каллусов Echinacea purpurea (А)
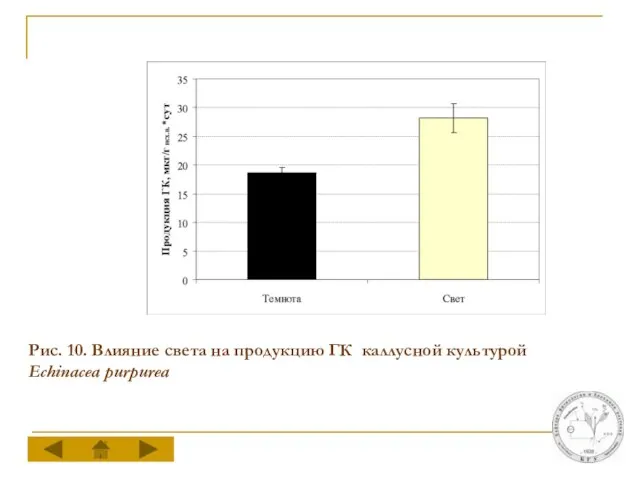
Слайд 17Рис. 10. Влияние света на продукцию ГК каллусной культурой Echinacea purpurea
Рис. 10. Влияние света на продукцию ГК каллусной культурой Echinacea purpurea
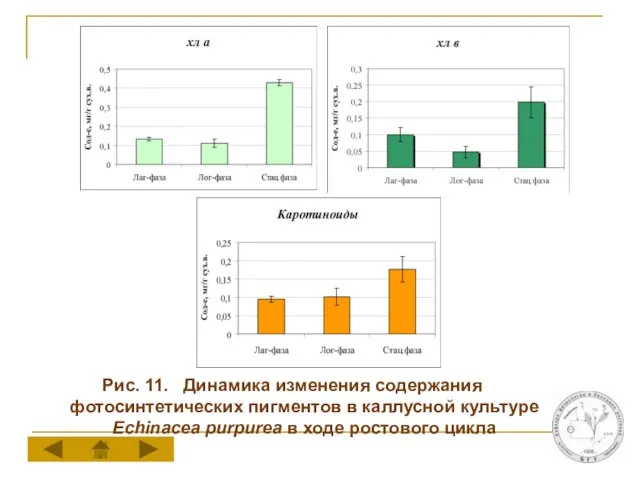
Слайд 18Рис. 11. Динамика изменения содержания фотосинтетических пигментов в каллусной культуре Echinacea purpurea
Рис. 11. Динамика изменения содержания фотосинтетических пигментов в каллусной культуре Echinacea purpurea
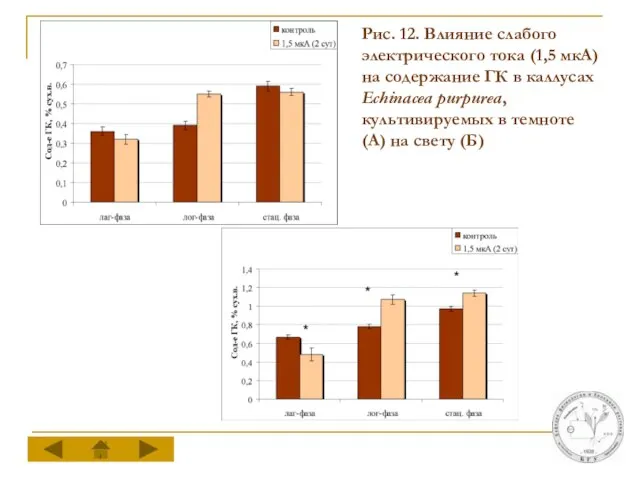
Слайд 19Рис. 12. Влияние слабого электрического тока (1,5 мкА) на содержание ГК в
Рис. 12. Влияние слабого электрического тока (1,5 мкА) на содержание ГК в
Слайд 20Выводы
Включение клеток суспензионной культуры эхинацеи пурпурной в полисахаридный носитель (Са-альгинатый гель) вызывает
Выводы
Включение клеток суспензионной культуры эхинацеи пурпурной в полисахаридный носитель (Са-альгинатый гель) вызывает

Слайд 21Высокая скорость прироста биомассы клеток при температуре 27°С компенсирует более низкие уровни
Высокая скорость прироста биомассы клеток при температуре 27°С компенсирует более низкие уровни

 Товар и деньги (8 класс)
Товар и деньги (8 класс) Welcome to Mexica
Welcome to Mexica V Астанинский экономический форум (22-24 мая 2012 года, г. Астана)
V Астанинский экономический форум (22-24 мая 2012 года, г. Астана) Юрий Гагарин первый космонавт в мире
Юрий Гагарин первый космонавт в мире Приглашаем вас в наш конный клуб
Приглашаем вас в наш конный клуб Изложение текста с рассуждением
Изложение текста с рассуждением Ряд Крокодили
Ряд Крокодили Презентация на тему Свет и цвет в природе
Презентация на тему Свет и цвет в природе  ТОиРChesla обслуживания дорог
ТОиРChesla обслуживания дорог Ринок туристичних услуг
Ринок туристичних услуг БИРЖЕВЫЕ НОВОСТИ –КАКИМИ ИМ БЫТЬ?На примере распространения информации Казахстанской фондовой биржей (KASE)
БИРЖЕВЫЕ НОВОСТИ –КАКИМИ ИМ БЫТЬ?На примере распространения информации Казахстанской фондовой биржей (KASE) Моя малая Родина – Питерский район
Моя малая Родина – Питерский район Щёлочноземельные металлы
Щёлочноземельные металлы Филиппины(характеристика)
Филиппины(характеристика) Интеграция новых ГХК с действующими агрохимическими предприятиями – перспективное направление создания газохимической отрасли
Интеграция новых ГХК с действующими агрохимическими предприятиями – перспективное направление создания газохимической отрасли  Презентация на тему "Мышонок Пик" 3 класс
Презентация на тему "Мышонок Пик" 3 класс Презентация на тему Владимир Михайлович Ануфриев
Презентация на тему Владимир Михайлович Ануфриев Информационные технологии в управлении
Информационные технологии в управлении Украина и украинцы
Украина и украинцы Verbs 4
Verbs 4 Презентация на тему Ответственные родители
Презентация на тему Ответственные родители Системы охлаждение персонального компьютера
Системы охлаждение персонального компьютера Prezentatsia_ZhKKh
Prezentatsia_ZhKKh «Моя математика» 1 класс
«Моя математика» 1 класс Презентация о звёздах и созвездиях
Презентация о звёздах и созвездиях Баскетбол. Правила
Баскетбол. Правила Россия в системе мирового рынка и международных союзов
Россия в системе мирового рынка и международных союзов Лот 6, г. Хабаровск, ул. Сысоева, 21, кв. 16
Лот 6, г. Хабаровск, ул. Сысоева, 21, кв. 16