- Взаимодействие генов. Сцепленное наследование

Содержание
- 2. ВЗАИМОДЕЙСТВИЕ ГЕНОВ Отклонение от законов Менделя вызывают и различные виды взаимодействия генов (за исключением полного доминирования).
- 3. Взаимодействие аллельных генов Взаимодействие аллельных генов называется внутриаллельным. Выделяют следующие его виды: полное доминирование, неполное доминирование,
- 4. Взаимодействие аллельных генов При полном доминировании один ген полностью подавляет проявление другого гена (выполняются законы Менделя),
- 5. Взаимодействие аллельных генов При неполном доминировании (промежуточном наследовании) доминантный ген не полностью подавляет проявление действия рецессивного
- 6. Взаимодействие аллельных генов При сверхдоминировании доминантный ген в гетерозиготном состоянии проявляет себя сильнее, чем в гомозиготном.
- 7. Взаимодействие аллельных генов При кодоминировании гены одной аллельной пары равнозначны, ни один из них не подавляет
- 8. Взаимодействие аллельных генов Своеобразные внутриаллельные взаимодействия наблюдаются в случаях множественных аллелей. Множественными называются аллели, которые представлены
- 9. Взаимодействие неаллельных генов Взаимодействие неаллельных генов называется межаллельным. Различают следующие его виды: комплементарность, эпистаз полимерию.
- 10. Взаимодействие неаллельных генов При комплементарности присутствие в одном генотипе двух доминантных (рецессивных) генов из разных аллельных
- 11. Взаимодействие неаллельных генов При эпистазе доминантный (рецессивный) ген из одной аллельной пары подавляет действие доминантного (рецессивного)
- 12. Взаимодействие неаллельных генов При полимерии доминантные гены из разных аллельных пар влияют на степень проявления одного
- 13. Взаимодействие неаллельных генов У человека может наблюдаться предрасположенность к различным заболеваниям: гипертонической болезни, ожирению, сахарному диабету,
- 14. 3. Сцепленное наследование У. Сэттон и Р. Пеннет в 1908 г. обнаружили отклонения от свободного комбинирования
- 15. Сцепленное наследование Для выяснения генотипа гибридов I поколения Морган провел анализирующее скрещивание. Он взял рецессивную гомозиготную
- 16. Схема свободного комбинирования генов Р B b V v B B b b V V v
- 17. Схема образования гамет при полном сцеплении P G V b B v B b V v
- 18. Сцепленное наследование Если взять дигетерозиготную самку мухи дрозофилы и скрестить ее с рецессивным самцом, то получается
- 19. Схема образования гамет при кроссинговере Каждая из образовавшихся хроматид попадает в отдельную гамету. Образуется четыре типа
- 20. Гаметы, в которые попали хроматиды не претерпевшие кроссинговер, называются некроссоверными, их обычно больше. Гаметы, в которые
- 21. Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну группу сцепления. Так как в
- 22. 4. Основные положения хромосомной теории наследственности (Т.Морган и соавт., 1911г.) 1. Гены расположены в хромосомах в
- 23. Цитологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором отмечается последовательность расположения генов.
- 25. Скачать презентацию
Слайд 2ВЗАИМОДЕЙСТВИЕ ГЕНОВ
Отклонение от законов Менделя вызывают и различные виды взаимодействия генов (за
ВЗАИМОДЕЙСТВИЕ ГЕНОВ
Отклонение от законов Менделя вызывают и различные виды взаимодействия генов (за

Слайд 3Взаимодействие аллельных генов
Взаимодействие аллельных генов называется внутриаллельным. Выделяют следующие его виды:
полное
Взаимодействие аллельных генов
Взаимодействие аллельных генов называется внутриаллельным. Выделяют следующие его виды:
полное

Слайд 4Взаимодействие аллельных генов
При полном доминировании один ген полностью подавляет проявление другого гена
Взаимодействие аллельных генов
При полном доминировании один ген полностью подавляет проявление другого гена

Слайд 5Взаимодействие аллельных генов
При неполном доминировании (промежуточном наследовании) доминантный ген не полностью подавляет
Взаимодействие аллельных генов
При неполном доминировании (промежуточном наследовании) доминантный ген не полностью подавляет

Слайд 6Взаимодействие аллельных генов
При сверхдоминировании доминантный ген в гетерозиготном состоянии проявляет себя сильнее,
Взаимодействие аллельных генов
При сверхдоминировании доминантный ген в гетерозиготном состоянии проявляет себя сильнее,

Слайд 7Взаимодействие аллельных генов
При кодоминировании гены одной аллельной пары равнозначны, ни один из
Взаимодействие аллельных генов
При кодоминировании гены одной аллельной пары равнозначны, ни один из

Слайд 8Взаимодействие аллельных генов
Своеобразные внутриаллельные взаимодействия наблюдаются в случаях множественных аллелей. Множественными называются
Взаимодействие аллельных генов
Своеобразные внутриаллельные взаимодействия наблюдаются в случаях множественных аллелей. Множественными называются

Слайд 9Взаимодействие неаллельных генов
Взаимодействие неаллельных генов называется межаллельным. Различают следующие его виды:
комплементарность,
Взаимодействие неаллельных генов
Взаимодействие неаллельных генов называется межаллельным. Различают следующие его виды:
комплементарность,

Слайд 10Взаимодействие неаллельных генов
При комплементарности присутствие в одном генотипе двух доминантных (рецессивных) генов
Взаимодействие неаллельных генов
При комплементарности присутствие в одном генотипе двух доминантных (рецессивных) генов

Слайд 11Взаимодействие неаллельных генов
При эпистазе доминантный (рецессивный) ген из одной аллельной пары подавляет
Взаимодействие неаллельных генов
При эпистазе доминантный (рецессивный) ген из одной аллельной пары подавляет

Слайд 12Взаимодействие неаллельных генов
При полимерии доминантные гены из разных аллельных пар влияют на
Взаимодействие неаллельных генов
При полимерии доминантные гены из разных аллельных пар влияют на

Слайд 13Взаимодействие неаллельных генов
У человека может наблюдаться предрасположенность к различным заболеваниям: гипертонической болезни,
Взаимодействие неаллельных генов
У человека может наблюдаться предрасположенность к различным заболеваниям: гипертонической болезни,

Слайд 143. Сцепленное наследование
У. Сэттон и Р. Пеннет в 1908 г. обнаружили отклонения
3. Сцепленное наследование
У. Сэттон и Р. Пеннет в 1908 г. обнаружили отклонения

Слайд 15Сцепленное наследование
Для выяснения генотипа гибридов I поколения Морган провел анализирующее скрещивание.
Сцепленное наследование
Для выяснения генотипа гибридов I поколения Морган провел анализирующее скрещивание.

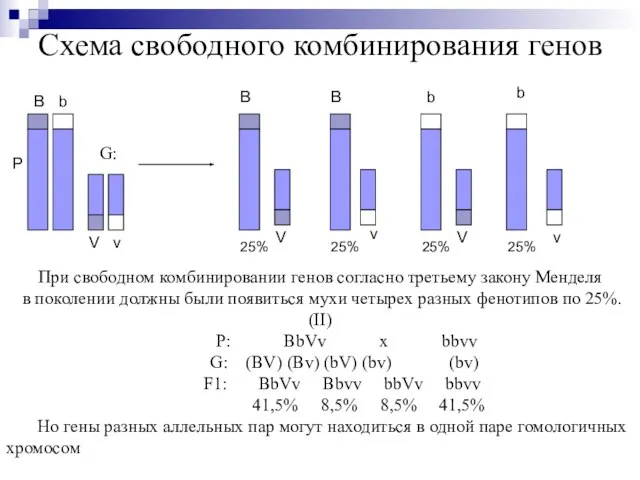
Слайд 16Схема свободного комбинирования генов
Р
B
b
V
v
B
B
b
b
V
V
v
v
При свободном комбинировании генов согласно третьему закону Менделя
в
Схема свободного комбинирования генов
Р
B
b
V
v
B
B
b
b
V
V
v
v
При свободном комбинировании генов согласно третьему закону Менделя
в
Слайд 17Схема образования гамет при полном сцеплении
P
G
V
b
B
v
B
b
V
v
Одна из пары гомологичных хромосом содержит
2
Схема образования гамет при полном сцеплении
P
G
V
b
B
v
B
b
V
v
Одна из пары гомологичных хромосом содержит
2

Слайд 18Сцепленное наследование
Если взять дигетерозиготную самку мухи дрозофилы и скрестить ее с рецессивным
Сцепленное наследование
Если взять дигетерозиготную самку мухи дрозофилы и скрестить ее с рецессивным

Слайд 19Схема образования гамет при кроссинговере
Каждая из образовавшихся хроматид попадает в отдельную гамету.
Схема образования гамет при кроссинговере
Каждая из образовавшихся хроматид попадает в отдельную гамету.

Слайд 20Гаметы, в которые попали хроматиды не претерпевшие кроссинговер, называются некроссоверными, их обычно
Гаметы, в которые попали хроматиды не претерпевшие кроссинговер, называются некроссоверными, их обычно

Слайд 21Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну группу
Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну группу

Слайд 224. Основные положения хромосомной теории наследственности (Т.Морган и соавт., 1911г.)
1. Гены расположены
4. Основные положения хромосомной теории наследственности (Т.Морган и соавт., 1911г.)
1. Гены расположены

Слайд 23Цитологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором
Цитологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором

 Музей Люфтваффе
Музей Люфтваффе 20170327_prezentatsiya_mirovye_resursy
20170327_prezentatsiya_mirovye_resursy SDL Trados Studio 2009 SP3 SDL Trados Team SDL MultiTerm Team
SDL Trados Studio 2009 SP3 SDL Trados Team SDL MultiTerm Team характеристика
характеристика Презентация на тему КУЛЬТУРА Культура в переводе с латинского означает возделывание, взращивание
Презентация на тему КУЛЬТУРА Культура в переводе с латинского означает возделывание, взращивание  Общие принципы диспансерного наблюдения
Общие принципы диспансерного наблюдения Верификацияавтоматных программ
Верификацияавтоматных программ Программирование на Бейсик
Программирование на Бейсик Интеллектуальное оповещение дежурно-диспетчерского персонала ДЦС-7
Интеллектуальное оповещение дежурно-диспетчерского персонала ДЦС-7 УЧИСЬ В ГОЛЛАНДИИ
УЧИСЬ В ГОЛЛАНДИИ  Неделя английского языка2011-2012
Неделя английского языка2011-2012 Презентация Microsoft PowerPoint
Презентация Microsoft PowerPoint 12 апреля –День космонавтики
12 апреля –День космонавтики Водоросли
Водоросли Корейская культура. Рисунок - музыка - поэзия
Корейская культура. Рисунок - музыка - поэзия Сессия стратегического планирования
Сессия стратегического планирования ФОРМИРОВАНИЕ МОТИВАЦИИ УЧЕБНОЙ ДЕЯТЕЛЬНОСТИ У СТУДЕНТОВ
ФОРМИРОВАНИЕ МОТИВАЦИИ УЧЕБНОЙ ДЕЯТЕЛЬНОСТИ У СТУДЕНТОВ Робот-манипулятор. Разработка конструкции в 3D, с применением CAD системы SolidWorks
Робот-манипулятор. Разработка конструкции в 3D, с применением CAD системы SolidWorks Проведение экспертизы раздела проектной документации «Мероприятия по обеспечению пожарной безопасности». Типовые несоответств
Проведение экспертизы раздела проектной документации «Мероприятия по обеспечению пожарной безопасности». Типовые несоответств Политическое сознание и политическое поведение
Политическое сознание и политическое поведение  Проект «Численность населения села Байдары»
Проект «Численность населения села Байдары» Давайте повторим:
Давайте повторим: Групповой проект
Групповой проект Налог на имущество
Налог на имущество Роль сметы в системе управления строительно-подрядным предприятием
Роль сметы в системе управления строительно-подрядным предприятием Человек, изменивший искусство. К 140-летию со дня рождения Пабло Пикассо
Человек, изменивший искусство. К 140-летию со дня рождения Пабло Пикассо Презентациябизнес-планаОбщие рекомендации
Презентациябизнес-планаОбщие рекомендации МГТУ ГА ФИЗИКА
МГТУ ГА ФИЗИКА