- Биомембраны. Функции и общая характеристика строения биомембран. Виды мембранного транспорта

Содержание
- 2. В числе тех, кто намеревался возглавить кафедру физиологии был Иван Петрович Павлов. В прошении министру народного
- 3. Владимир Николаевич Великий Основателем и первым заведующим кафедрой физиологии был профессор Владимир Николаевич Великий. В 1874
- 4. Алексей Александрович Кулябко Профессор Алексей Александрович Кулябко – выдающийся отечественный физиолог, возглавлявший кафедру с 1903 –
- 5. А.А. Кулябко внес значительный вклад в развитие мировой науки. 3 августа 1902 г. он впервые в
- 6. Николай Александрович Попов Профессор Николай Александрович Попов заведовал кафедрой в 1925-28 гг. Н.А. Попов в 1925
- 7. Борис Иванович Баяндуров Профессор Борис Иванович Баяндуров – ученик профессора Н.А. Попова. Первый заведующий кафедрой нормальной
- 8. В 1946 году за монографию «Трофическая функция головного мозга» он был удостоен Сталинской (Государственной) премии второй
- 9. Евгений Федорович Ларин Профессор Евгений Федорович Ларин заведовал кафедрой с 1948 по 1975 г. В этот
- 10. Михаил Андреевич Медведев С 1975 и по настоящее время кафедру нормальной физиологии ТМИ – СибГМУ возглавляет
- 12. Основная литература Нормальная физиология [Текст] : учебник для студентов медицинских вузов по специальностям "Лечебное дело" и
- 13. Дополнительная литература: 1. Практикум по физиологии для студентов врачебных факультетов [Электронный ресурс] : учебное пособие для
- 14. БИОМЕМБРАНЫ ПЛАН ЛЕКЦИИ: ИСТОРИЯ ИЗУЧЕНЯ. ФУНКЦИИ И ОБЩАЯ ХАРАК-ТЕРИСТИКА СТРОЕНИЯ БИОМЕМБРАН. ВИДЫ МЕМБРАННОГО ТАНСПОРТА.
- 15. Цель лекции: Дать характеристику строения и функций биомембран и её структурных компонентов. Разобрать виды мембранного транспорта.
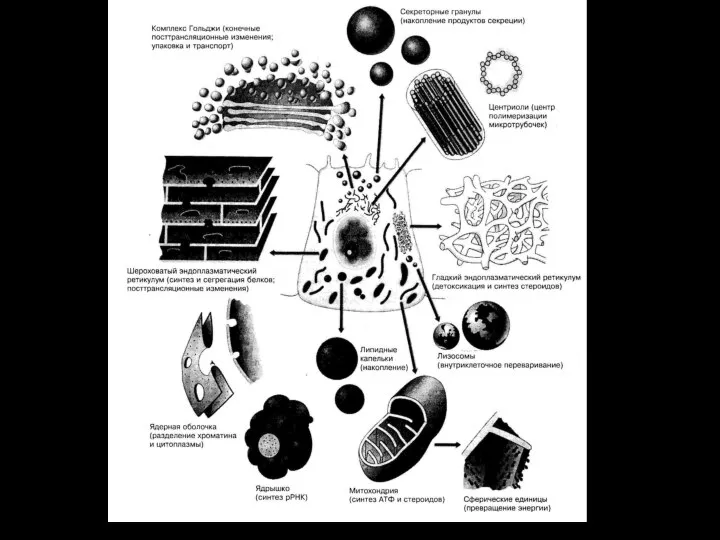
- 16. Биологическими мембранами (от лат. membrana – перепонка) Называют функциональные струк-туры клетки, ограничивающие цито-плазму и внутриклеточные структуры.
- 17. Половина объема клетки занята органеллами. Общая площадь поверхности мембран внутриклеточных органелл в 10 раз! превышает поверхность
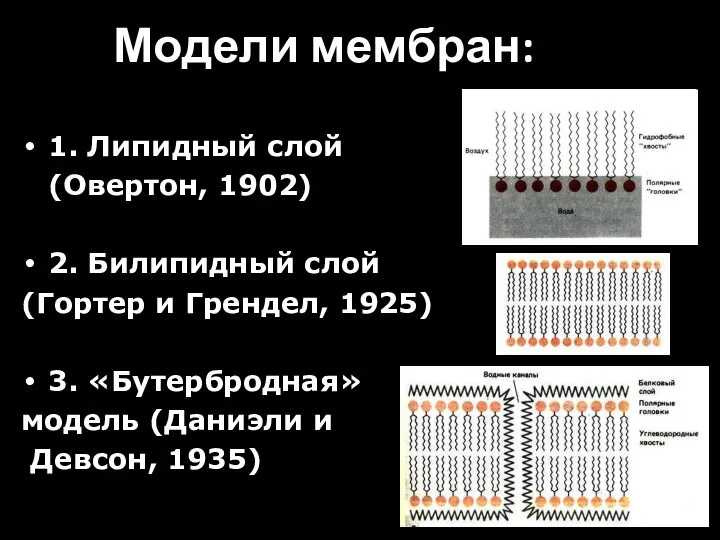
- 19. Модели мембран: 1. Липидный слой (Овертон, 1902) 2. Билипидный слой (Гортер и Грендел, 1925) 3. «Бутербродная»
- 20. В 1925 г. Гортер и Грендель провели ацетоновую экстракцию теней эритроцитов (мембраны лопнувших в гипотоническом растворе
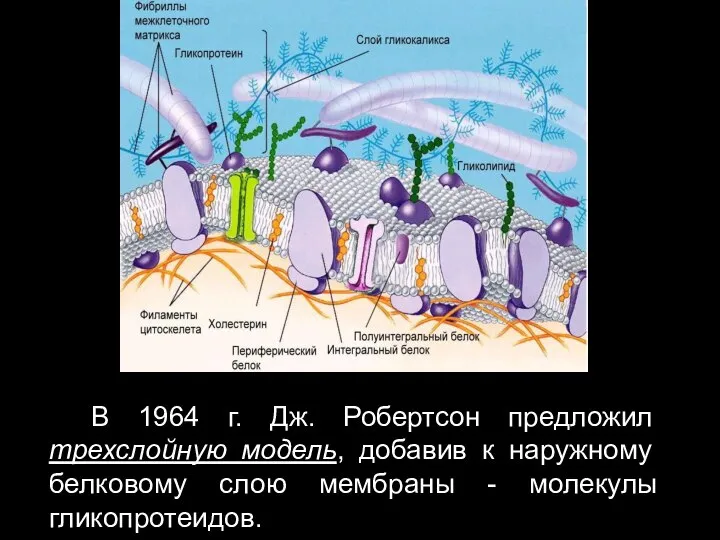
- 21. В 1964 г. Дж. Робертсон предложил трехслойную модель, добавив к наружному белковому слою мембраны - молекулы
- 22. 1966 г. Дж. Ленард и С. Сингер – предложили жидкомозаичную модель. Согласно этой модели белки «плавают»
- 23. Функции биомембран: 1. Барьерная функция - обусловливает создание концентрационных градиентов, являющихся основой механизма электрогенеза (потенциал покоя,
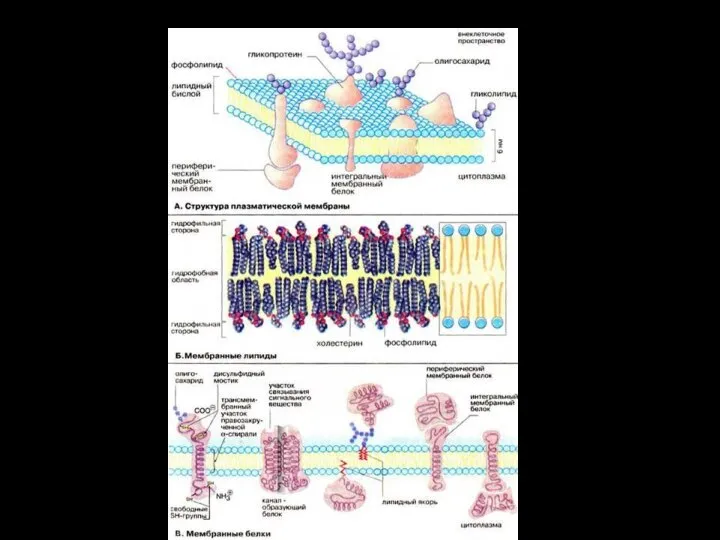
- 24. Структурные элементы мембран: Липиды Белки Углеводы Вода
- 25. Толщина биомембран составляет 4 -10 нм. Соотношение в них между белками и липидами зависит от типа
- 26. Липидный бислой представлен: фосфолипидами (глицерофосфатидами) сфингомиелинами стероидными липидами — холестерином (холестеролом).
- 27. Фосфолипиды. Составляют до 80% от всей массы липидов клеточной мембраны. Молекула фосфолипида состоит из полярной (гидрофильной)
- 28. фосфолипиды Состоят из полярной (гидрофильной) головки, шейки и неполярных (гидрофобных) хвостов. Головка образована остатком фосфорной кислоты.
- 29. Характеристические (полярные) группы фосфолипидов
- 32. Сфинголипиды - содержат аминоспирт сфингозин. Участвуют в образовании: миелиновой оболочки аксонов (сфингомиелины), гликокалекса (ганглиолипиды), отрицательного заряда
- 33. Стеролы (неполярные молекулы). Молекулы стеролов распределяются среди фосфолипидов, плавая подобно айсбергам, они стабилизируют текучесть мембраны, увеличивают
- 34. Фосфолипиды и холестерол
- 35. Мембрана черезвычайно динамичная структура. Характерным свойством мембраны является латеральная и продольная диффузия липидов. 1 – латеральная
- 36. Функциональная классификация мембранных белков: 1. Белки – каналы Обеспечивают транспорт ионов. 2. Белки – транспортёры Участвуют
- 37. Структурная классификация белков мембраны: Белки биологических мембран подразделяют на периферические и интегральные (трансмембранные) . Периферические мембранные
- 38. Интегральные мембранные белки встроены в липидный бислой. Их гидрофильные аминокислоты взаимодействуют с фосфатными группами фосфолипидов, а
- 39. Углеводы Углеводы в составе мембран обнаруживаются лишь в соединении с белками (гликопротеины и протеогликаны) и липидами
- 40. Функции углеводов межклеточное узнавание, межклеточные взаимодействия, поддержание иммунного статуса клетки, обеспечение стабильности белковых молекул в мембране,
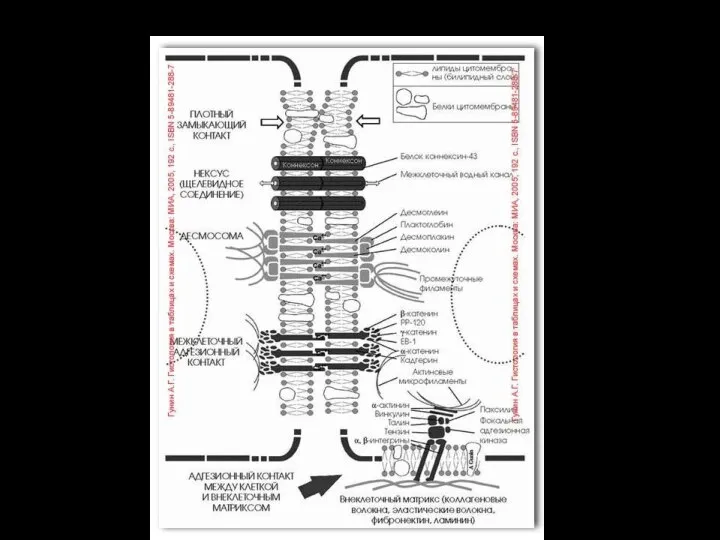
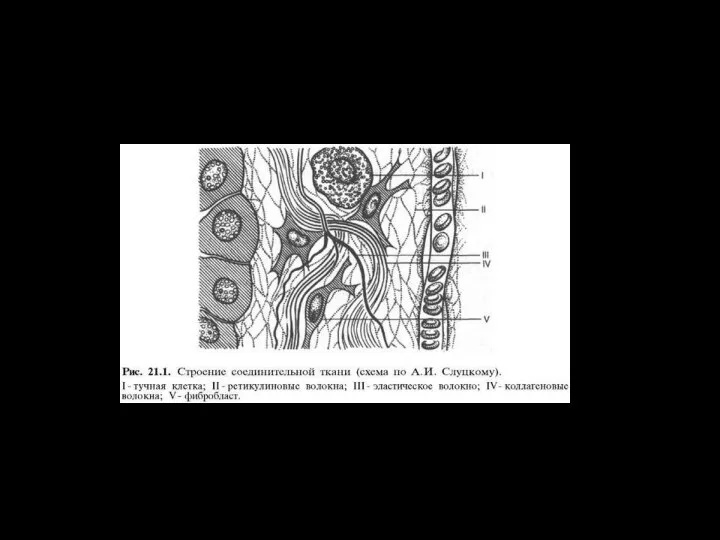
- 41. Взаимодействие цитоскелета с гликокаликсом I – протеогликан, II – коллаген, III – фибронектин (образует плотную сеть),
- 42. Вода Свободная вода омывает мембрану, заполняет каналы, поры и кинки. Вода может находится между липидными слоями
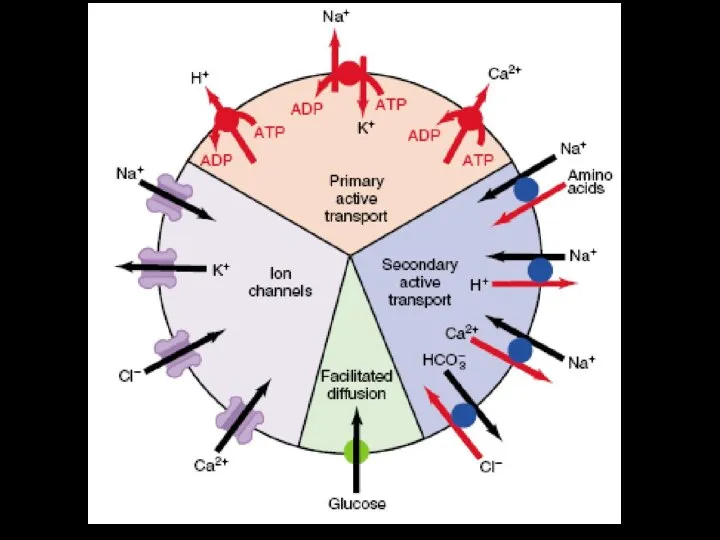
- 43. Транспортные процессы мембраны обеспечивают: 1. Поддержание объема клетки и внутриклеточного ионного состава в определенном интервале, который
- 44. Виды мембранного транспорта: Прямой Опосредованный Прямой (пассивный) транспорт: 1. простая диффузя; 2. фильтрация; 3. осмос; 4.
- 45. Пассивный транспорт Идет без затраты энергии АТФ! Движущие силы: Градиент концентрации вещества (химический градиент) Градиент концентрации
- 46. Диффузия – самопроизвольный процесс проникновения вещества из области большей концентрации в область меньшей его концентрации, в
- 47. 1. Простая диффузия идет непосредственно ЧЕРЕЗ МЕМБРАНУ для незаряженных (жирорастворимых) веществ Таким образом проис-ходит движение небольших
- 48. Посредством простой диффузии, через мембрану, переносятся в основном газы и неполярные вещества, обладающие высокой гидрофобностью. Гидрофильные
- 50. Проницаемость искусственных липидных бислоев для различных веществ
- 51. Простая диффузия может осуществляться ЧЕРЕЗ КИНКИ При повышении температуры тепловая подвижность жирнокислотных цепей приводит к спонтанному
- 53. Простая диффузия идет 3. ЧЕРЕЗ ПОРЫ Канал поры всегда открыт, поэтому химическое вещество проходит через мембрану
- 54. Биомембраны имеют аномально высокую проницаемость для воды. Это объясняется наличием в мембране лабильных и фиксированных водных
- 55. Фильтрация Это движение растворенных веществ под действием гидро-статического давления.
- 56. Осмос — поток воды через полупроницаемую мембрану из компартмента с меньшей концентрацией растворённых в воде осмотически
- 57. Распределение воды между клеточными компартментами, цитозолем и органоидами клетки, между клеткой и интерстициальной жидкостью и её
- 58. Электроосмос В случае электроосмоса движущей силой является электрохимический градиент.
- 59. С участием С участием переносчиков каналов унипорт котранспорт Потен/чувств. Хемо/чувств. Мех/чувств. симпорт Активный транспорт Облегченная диффузия
- 60. ОБЛЕГЧЁННАЯ ДИФФУЗИЯ Для облегчённой диффузии веществ необходимы встроенные в мембрану белковые компоненты (переносчики, каналы). Все эти
- 61. Переносчики Переносчики (транспортёры) осуществляют транспорт через биологические мембраны множества различных ионов (Na+, Cl–, H+, HCO3– и
- 63. Симпорт. Сочетанный транспорт глюкозы и Na+ в тонком кишечнике и канальцах почки обеспечивают мембранные гликопротеины, кодируемые
- 64. Симпорт аминокислот и Na+
- 65. Антипорт: Катионные обменники : Nа+/Н+ – обмен, Nа+/Са+–обмен (2Nа/Са) Анионные обменники : СL-/НСО3-
- 67. По механизму действия транспортеры делятся на два типа: 1 тип – совершают челночные движения через мембрану
- 68. Транспорт К+ валиномицином
- 69. Грамицидин, способен встраиваться в мембрану, образуя канал, по которому могут перемещаться протоны и другие одно-валентные катионы
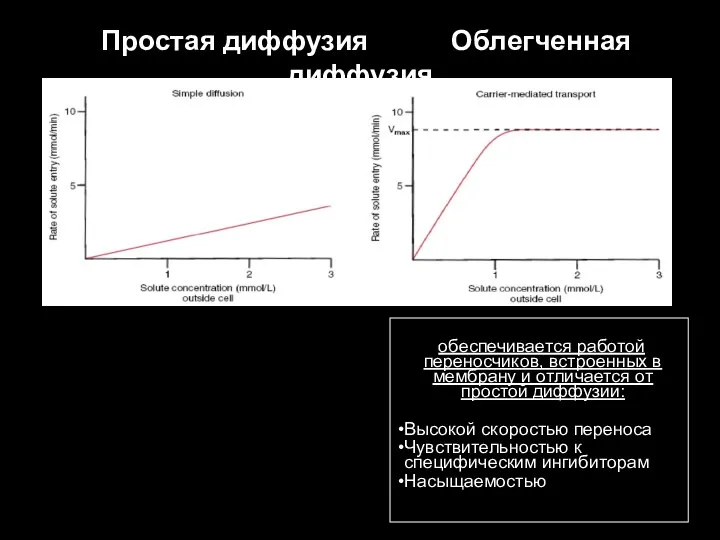
- 70. Простая диффузия Облегченная диффузия обеспечивается работой переносчиков, встроенных в мембрану и отличается от простой диффузии: Высокой
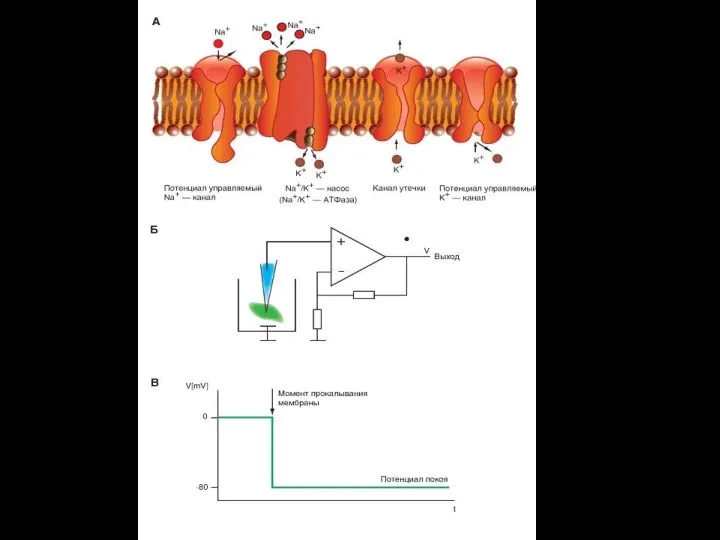
- 71. Ионные каналы – интегральные белки мембраны, выполняющие функции транспорта для соответствующих ионов. Они различаются по избирательной
- 72. Потенциалозависимые ионные каналы управляются мембранным потенциалом (МП). Колебания МП приводят к конформационным изменениям белковой структуры канала,
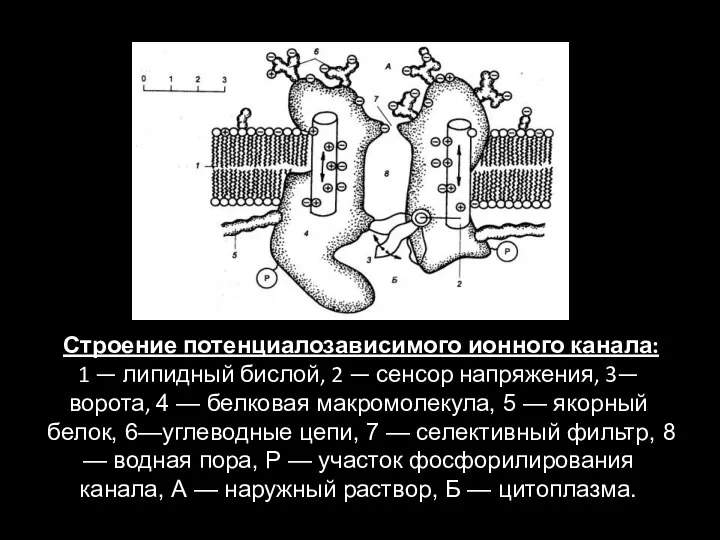
- 73. Строение потенциалозависимого ионного канала: 1 — липидный бислой, 2 — сенсор напряжения, 3— ворота, 4 —
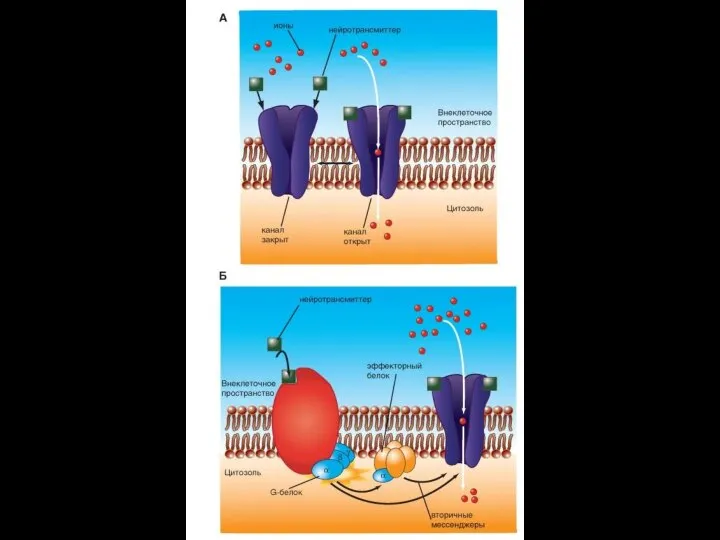
- 74. Xемоуправляемые каналы ( лиганд–зависимые ионные каналы, рецептор–зависимые) Соединение лиганда с рецептором вызывает конформационные изменения в канале,
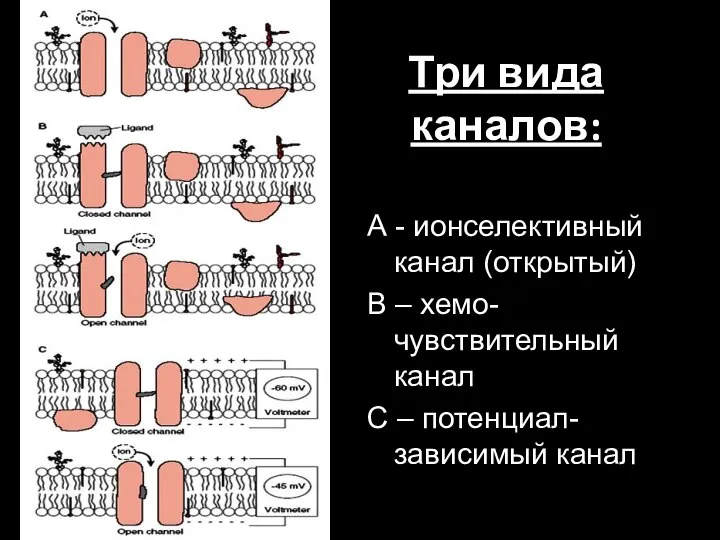
- 76. Три вида каналов: А - ионселективный канал (открытый) В – хемо-чувствительный канал С – потенциал-зависимый канал
- 77. Активный транспорт Осуществляется против электрохимического градиента; Система в высшей степени специфична; Необходимы источники энергии в виде
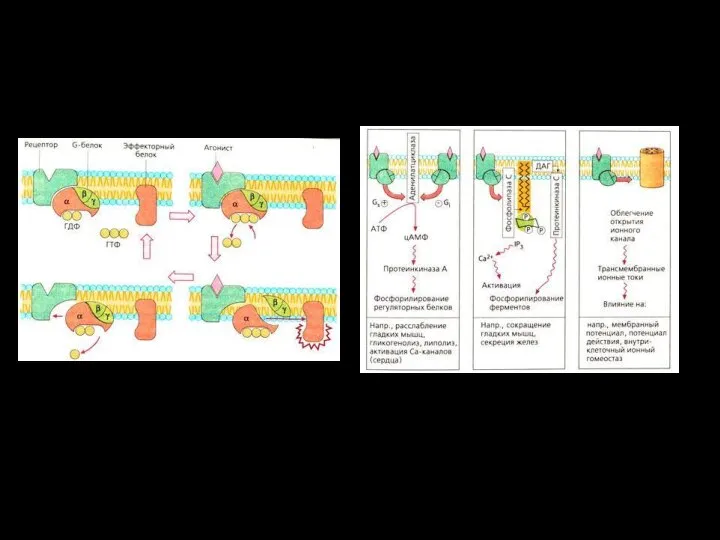
- 78. Первично активный транспорт Движущая сила трансмембранного переноса возникает при ферментативном гидролизе макроэргических связей АТФ. 3Na+/2K+ или
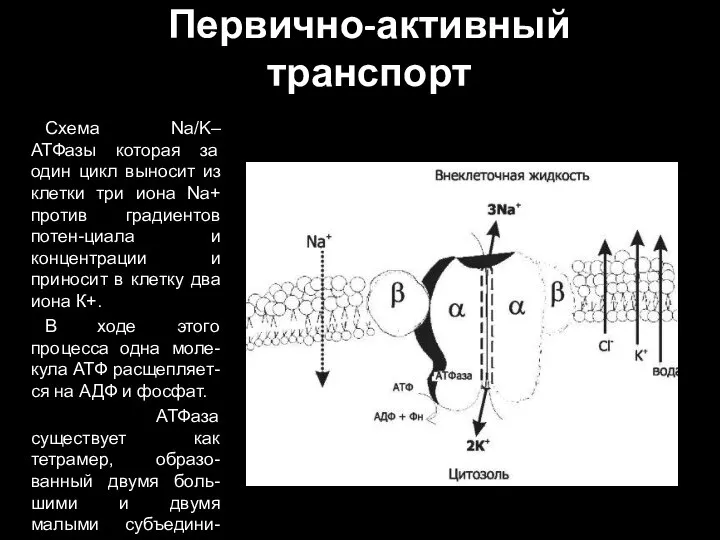
- 79. Первично-активный транспорт Схема Na/K–АТФазы которая за один цикл выносит из клетки три иона Na+ против градиентов
- 80. Работа натрий-калиевого насоса
- 81. Вторично активный транспорт. Движущая сила для трансмембранного переноса одного вещества (или ионов) против электро-химического градиента возникает
- 82. Вторичный активный транспорт В качестве источника энергии использует химичес-кий или электрохимический градиент какого-либо ве-щества .
- 83. Эндоцитоз Образуется окаймленная везикула (3), Везикула сливается с вакуолью (4). Вакуоль и ее содержимое претерпевают превращения
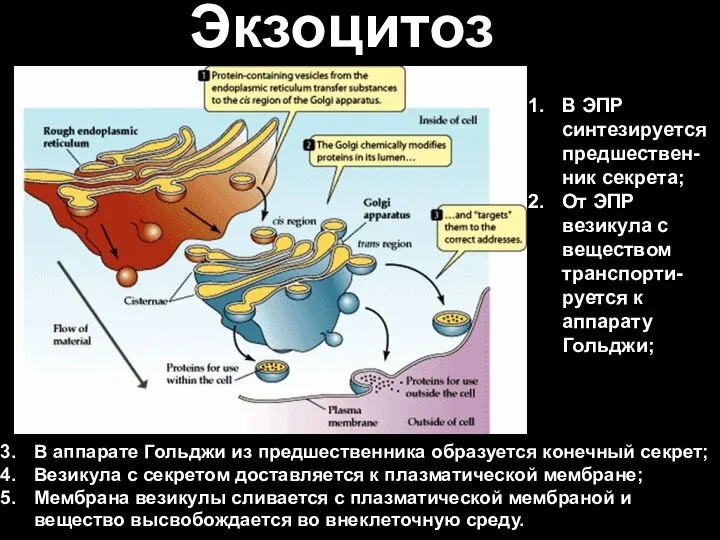
- 84. Экзоцитоз В аппарате Гольджи из предшественника образуется конечный секрет; Везикула с секретом доставляется к плазматической мембране;
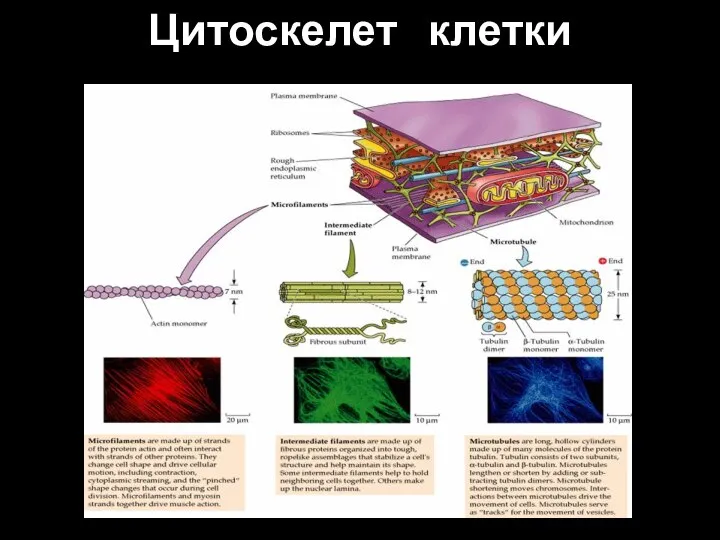
- 85. Цитоскелет клетки
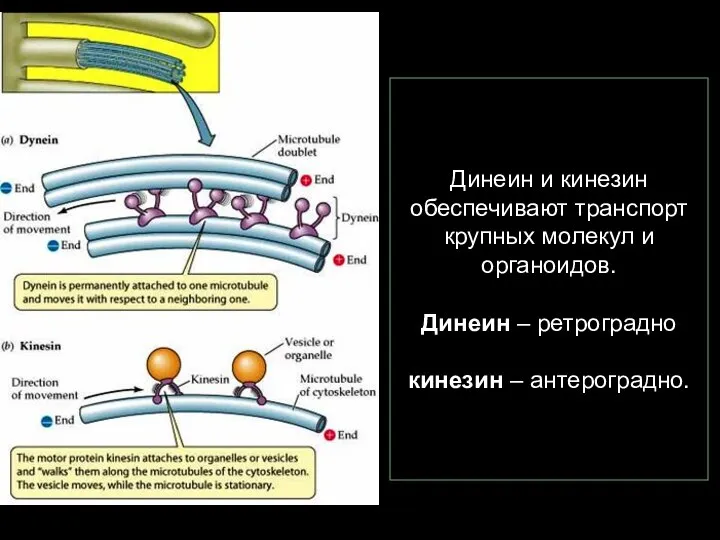
- 87. Динеин и кинезин обеспечивают транспорт крупных молекул и органоидов. Динеин – ретроградно кинезин – антероградно.
- 89. Основные проявления жизнедеятельности Физиологический покой Физиологическая активность Раздражение Возбуждение Торможение
- 90. Общая характеристика возбудимых тканей. Все клетки нашего организма обладают свойством раздражимости. Раздражимость – это способность клеток
- 91. Раздражение и виды раздражителей. Раздражение – это процесс воздействия на живой объект внешних по отношению к
- 92. По природе раздражителя: 1. физические ( температура, звук, свет, электрический ток и т.д.). 2. химические (соли,
- 93. По биологическому значению: 1. адекватные – раздражитель к которому в естественных условиях у биосистем есть специализированные
- 94. По количественному признаку или порогу раздражения: В зависимости от силы действия раздражители делятся на: 1. пороговые
- 95. Все возбудимые ткани обладают общими свойствами: Возбудимостью Проводимостью Лабильностью Аккомодацией
- 96. Возбудимость количественно характеризуется порогом возбуждения, который определяется минимальной силой раздражения, способной вызывать возбуждение. Проводимость – способность
- 97. Законы раздражения: Закон силы Закон времени Закон крутизны нарастания раздражителя
- 98. Закон силы Чем сильней раздражение, тем до определенного предела сильнее ответная реакция биосистемы. Закон постулирован для
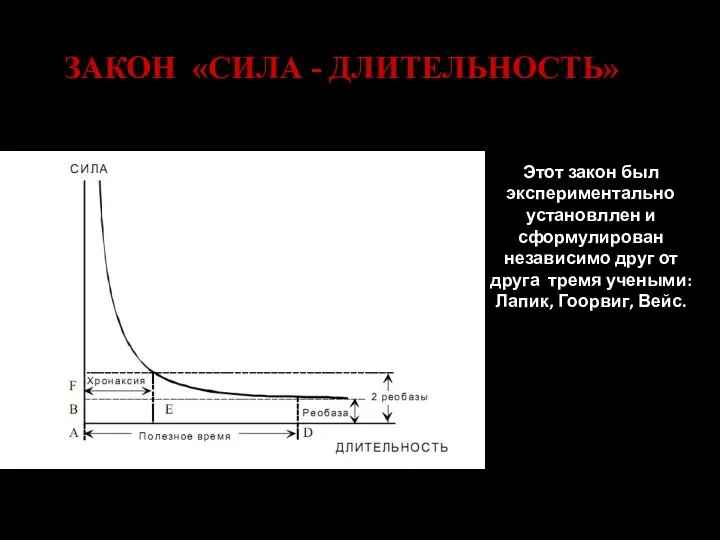
- 99. Закон времени Чем длительнее раздражение, тем сильней до определенных пределов ответная реакция живой ткани. В определенном
- 100. ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ» Этот закон был экспериментально установллен и сформулирован независимо друг от друга тремя
- 102. Закон крутизны нарастания раздражителя или градиента: Чем выше крутизна нарастания раздражителя во времени, тем больше до
- 103. Спасибо за внимание
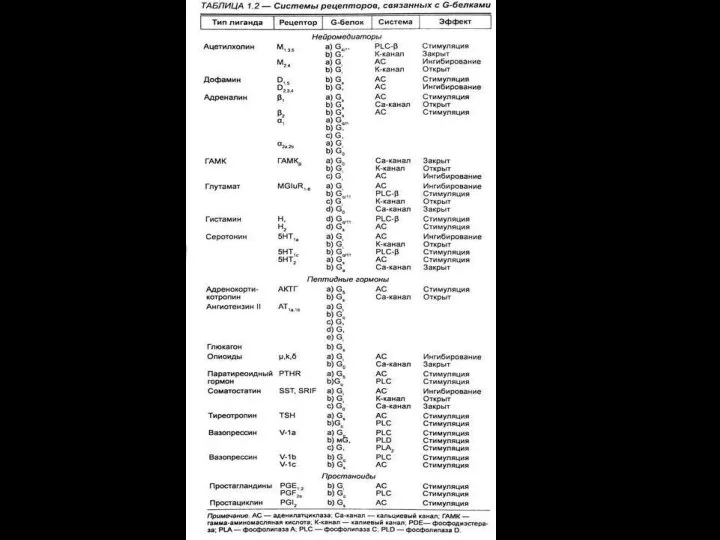
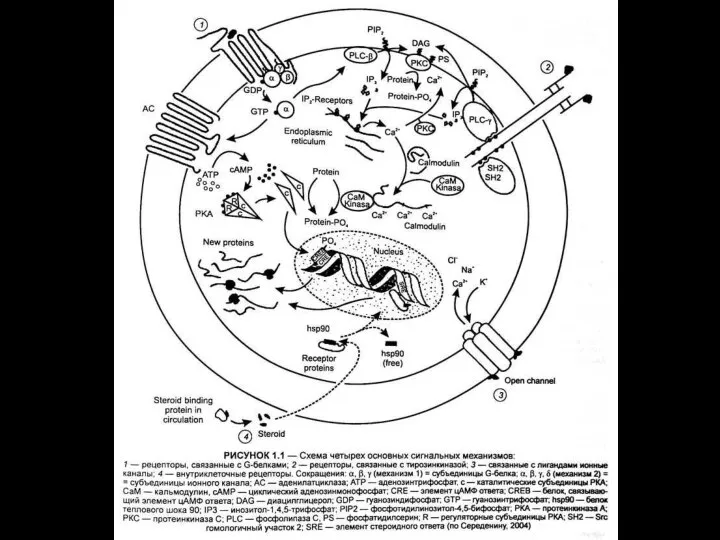
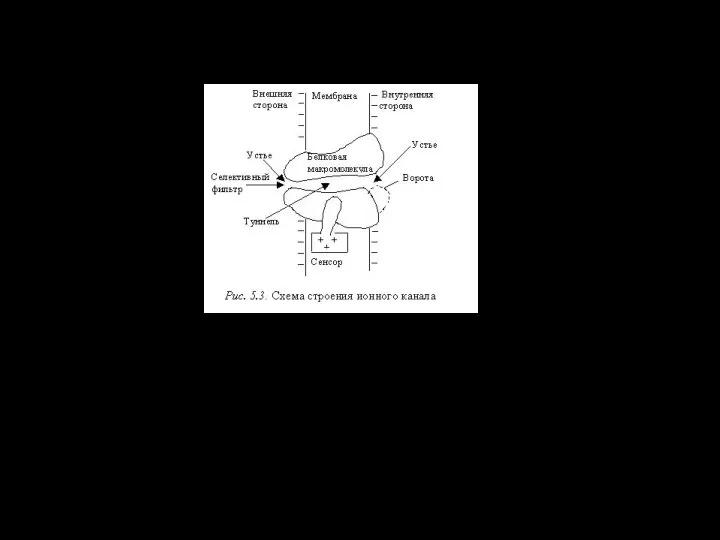
- 116. ионный канал состоит из связанных между собой белковых субъединиц, формирующих в мембране гидрофильную селективную пору
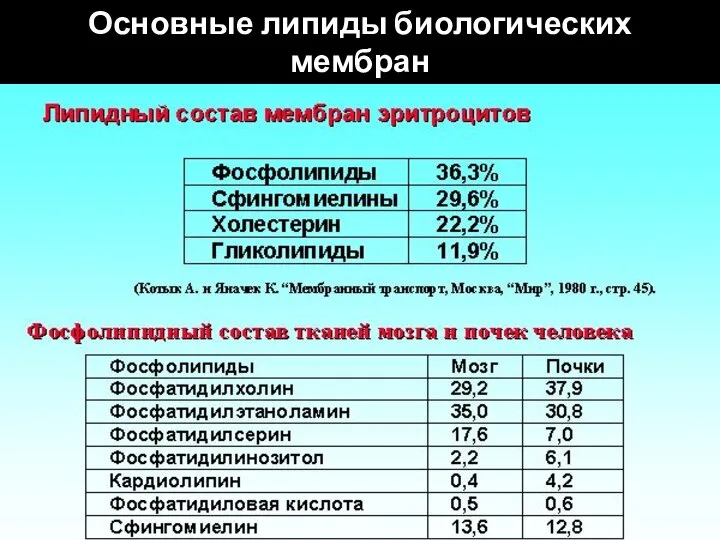
- 117. Основные липиды биологических мембран
- 121. СПОСОБЫ УПАКОВКИ ФОСФОЛИПИДОВ
- 125. Потенциал-управляемый канал
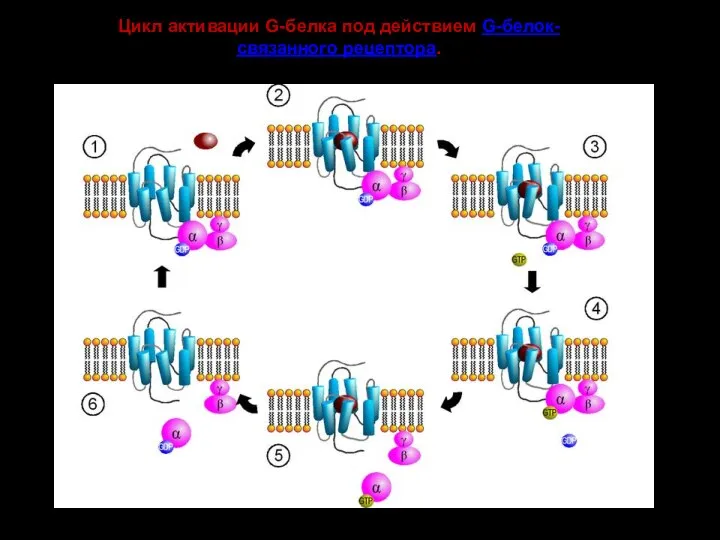
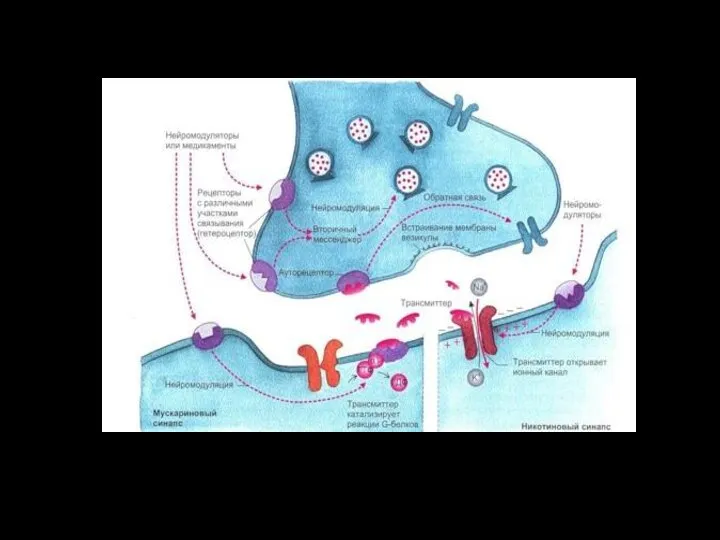
- 129. Цикл активации G-белка под действием G-белок-связанного рецептора.
- 146. Основные липиды биологических мембран
- 148. Скачать презентацию

Слайд 3Владимир Николаевич Великий
Основателем и первым заведующим кафедрой физиологии был профессор
Владимир Николаевич Великий
Основателем и первым заведующим кафедрой физиологии был профессор

Слайд 4Алексей Александрович Кулябко
Профессор Алексей Александрович Кулябко – выдающийся отечественный физиолог,
Алексей Александрович Кулябко
Профессор Алексей Александрович Кулябко – выдающийся отечественный физиолог,

Слайд 5А.А. Кулябко внес значительный вклад в развитие мировой науки. 3 августа 1902
А.А. Кулябко внес значительный вклад в развитие мировой науки. 3 августа 1902

Слайд 6Николай Александрович Попов
Профессор Николай Александрович Попов заведовал кафедрой в 1925-28
Николай Александрович Попов
Профессор Николай Александрович Попов заведовал кафедрой в 1925-28

Слайд 7Борис Иванович Баяндуров
Профессор Борис Иванович Баяндуров – ученик профессора Н.А.
Борис Иванович Баяндуров
Профессор Борис Иванович Баяндуров – ученик профессора Н.А.

Слайд 8В 1946 году за монографию «Трофическая функция головного мозга» он был удостоен
В 1946 году за монографию «Трофическая функция головного мозга» он был удостоен

Слайд 9Евгений Федорович Ларин
Профессор Евгений Федорович Ларин заведовал кафедрой с 1948
Евгений Федорович Ларин
Профессор Евгений Федорович Ларин заведовал кафедрой с 1948

Слайд 10Михаил Андреевич Медведев
С 1975 и по настоящее время кафедру нормальной
Михаил Андреевич Медведев
С 1975 и по настоящее время кафедру нормальной

Слайд 12Основная литература
Нормальная физиология [Текст] : учебник для студентов медицинских вузов по специальностям "Лечебное
Основная литература
Нормальная физиология [Текст] : учебник для студентов медицинских вузов по специальностям "Лечебное
![Основная литература Нормальная физиология [Текст] : учебник для студентов медицинских вузов по](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1003192/slide-11.jpg)
Слайд 13 Дополнительная литература:
1. Практикум по физиологии для студентов врачебных факультетов [Электронный ресурс] :
Дополнительная литература:
1. Практикум по физиологии для студентов врачебных факультетов [Электронный ресурс] :
![Дополнительная литература: 1. Практикум по физиологии для студентов врачебных факультетов [Электронный ресурс]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1003192/slide-12.jpg)
Слайд 14БИОМЕМБРАНЫ
ПЛАН ЛЕКЦИИ:
ИСТОРИЯ ИЗУЧЕНЯ.
ФУНКЦИИ И ОБЩАЯ ХАРАК-ТЕРИСТИКА СТРОЕНИЯ БИОМЕМБРАН.
ВИДЫ МЕМБРАННОГО ТАНСПОРТА.
БИОМЕМБРАНЫ
ПЛАН ЛЕКЦИИ:
ИСТОРИЯ ИЗУЧЕНЯ.
ФУНКЦИИ И ОБЩАЯ ХАРАК-ТЕРИСТИКА СТРОЕНИЯ БИОМЕМБРАН.
ВИДЫ МЕМБРАННОГО ТАНСПОРТА.

Слайд 15Цель лекции:
Дать характеристику строения и функций биомембран и её структурных компонентов. Разобрать
Цель лекции:
Дать характеристику строения и функций биомембран и её структурных компонентов. Разобрать

Слайд 16
Биологическими мембранами
(от лат. membrana – перепонка)
Называют функциональные струк-туры клетки, ограничивающие цито-плазму и
Биологическими мембранами
(от лат. membrana – перепонка)
Называют функциональные струк-туры клетки, ограничивающие цито-плазму и

Слайд 17Половина объема клетки занята органеллами.
Общая площадь поверхности мембран внутриклеточных органелл в
Половина объема клетки занята органеллами.
Общая площадь поверхности мембран внутриклеточных органелл в

Слайд 19Модели мембран:
1. Липидный слой
(Овертон, 1902)
2. Билипидный слой
(Гортер и Грендел, 1925)
3.
Модели мембран:
1. Липидный слой
(Овертон, 1902)
2. Билипидный слой
(Гортер и Грендел, 1925)
3.
Слайд 20В 1925 г. Гортер и Грендель провели ацетоновую экстракцию теней эритроцитов (мембраны
В 1925 г. Гортер и Грендель провели ацетоновую экстракцию теней эритроцитов (мембраны

Слайд 21В 1964 г. Дж. Робертсон предложил трехслойную модель, добавив к наружному белковому
В 1964 г. Дж. Робертсон предложил трехслойную модель, добавив к наружному белковому
Слайд 22 1966 г. Дж. Ленард и С. Сингер – предложили жидкомозаичную модель.
1966 г. Дж. Ленард и С. Сингер – предложили жидкомозаичную модель.

Слайд 23
Функции биомембран:
1. Барьерная функция - обусловливает создание концентрационных градиентов, являющихся основой механизма
Функции биомембран:
1. Барьерная функция - обусловливает создание концентрационных градиентов, являющихся основой механизма

Слайд 24Структурные элементы мембран:
Липиды
Белки
Углеводы
Вода
Структурные элементы мембран:
Липиды
Белки
Углеводы
Вода

Слайд 25Толщина биомембран составляет 4 -10 нм.
Соотношение в них между белками и
Толщина биомембран составляет 4 -10 нм.
Соотношение в них между белками и

Слайд 26Липидный бислой представлен:
фосфолипидами (глицерофосфатидами)
сфингомиелинами
стероидными липидами — холестерином (холестеролом).
Липидный бислой представлен:
фосфолипидами (глицерофосфатидами)
сфингомиелинами
стероидными липидами — холестерином (холестеролом).

Слайд 27Фосфолипиды.
Составляют до 80% от всей массы липидов клеточной мембраны.
Молекула фосфолипида состоит
Фосфолипиды.
Составляют до 80% от всей массы липидов клеточной мембраны.
Молекула фосфолипида состоит

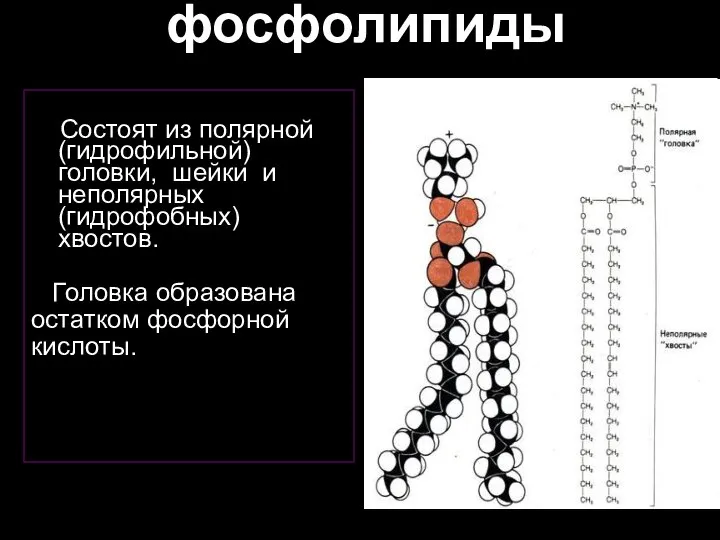
Слайд 28фосфолипиды
Состоят из полярной (гидрофильной) головки, шейки и неполярных (гидрофобных) хвостов.
Головка
фосфолипиды
Состоят из полярной (гидрофильной) головки, шейки и неполярных (гидрофобных) хвостов.
Головка
Слайд 29
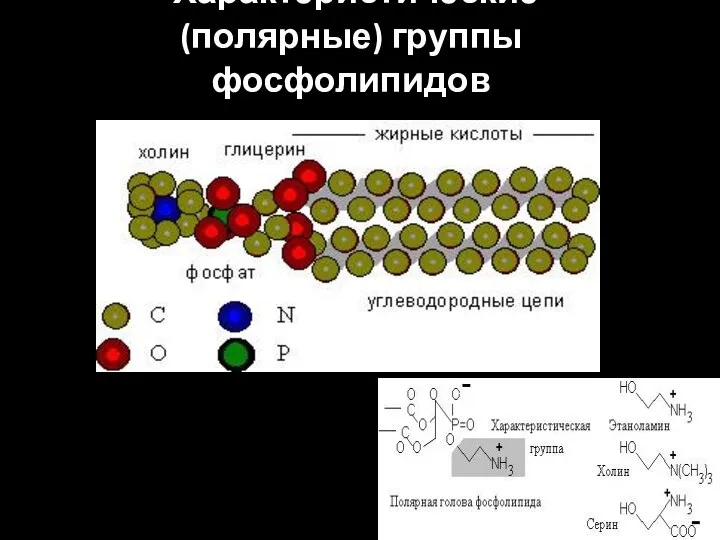
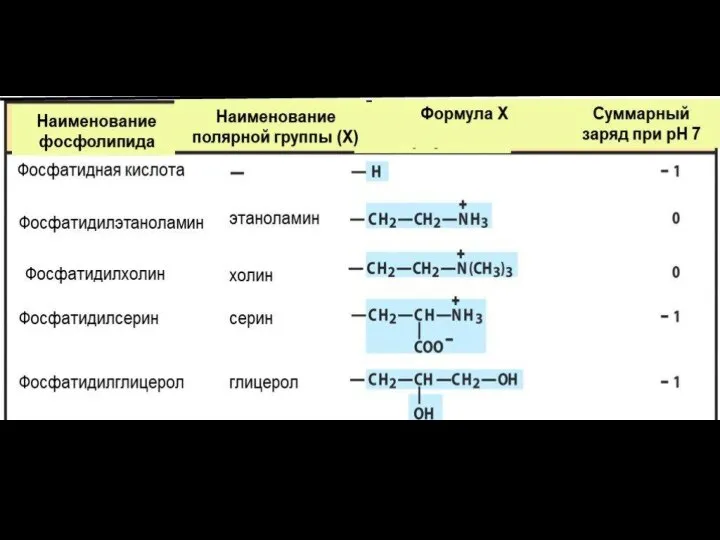
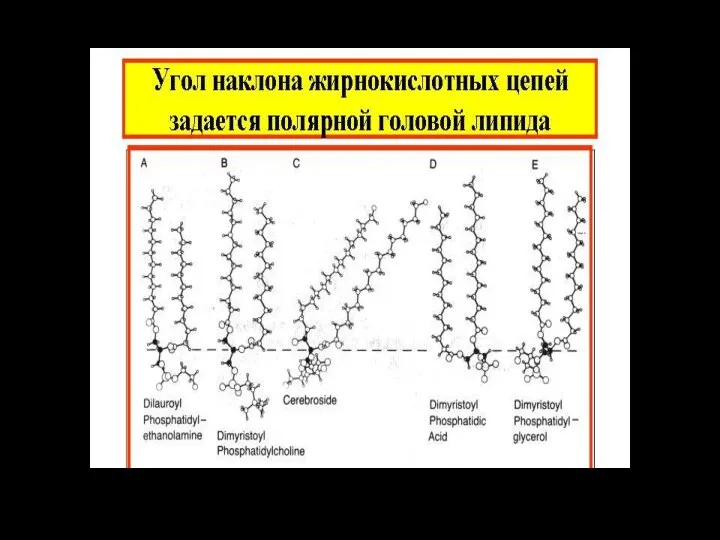
Характеристические (полярные) группы фосфолипидов
Характеристические (полярные) группы фосфолипидов
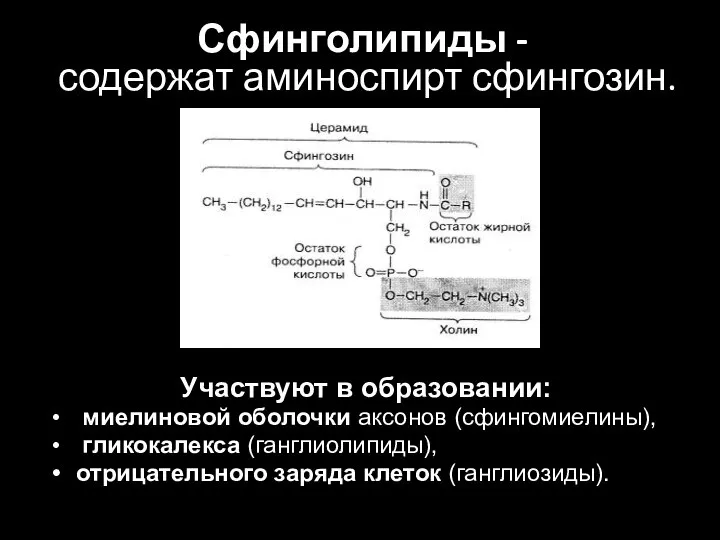
Слайд 32Сфинголипиды -
содержат аминоспирт сфингозин.
Участвуют в образовании:
миелиновой оболочки аксонов (сфингомиелины),
Сфинголипиды -
содержат аминоспирт сфингозин.
Участвуют в образовании:
миелиновой оболочки аксонов (сфингомиелины),
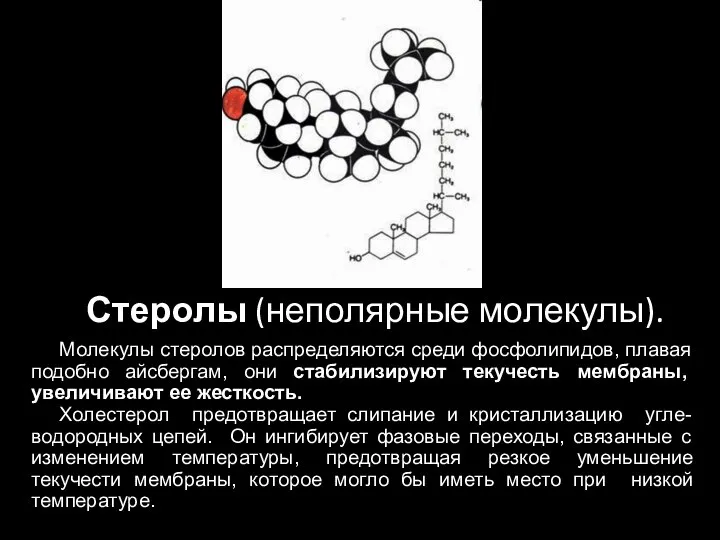
Слайд 33Стеролы (неполярные молекулы).
Молекулы стеролов распределяются среди фосфолипидов, плавая подобно айсбергам, они стабилизируют
Стеролы (неполярные молекулы).
Молекулы стеролов распределяются среди фосфолипидов, плавая подобно айсбергам, они стабилизируют
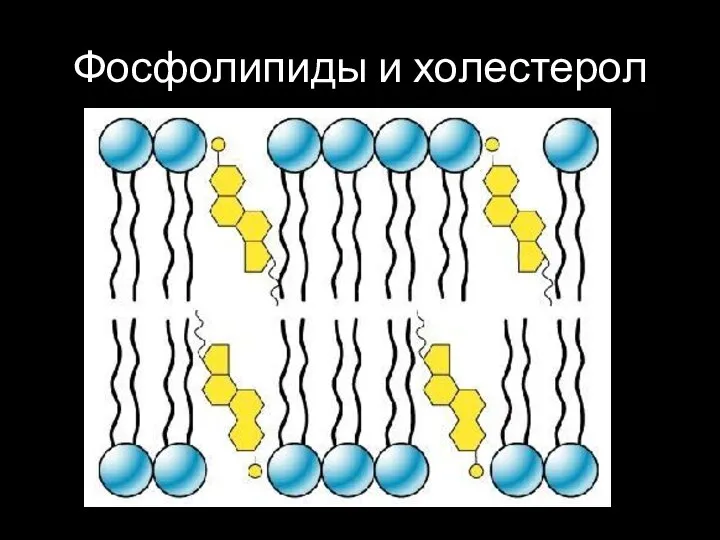
Слайд 34Фосфолипиды и холестерол
Фосфолипиды и холестерол
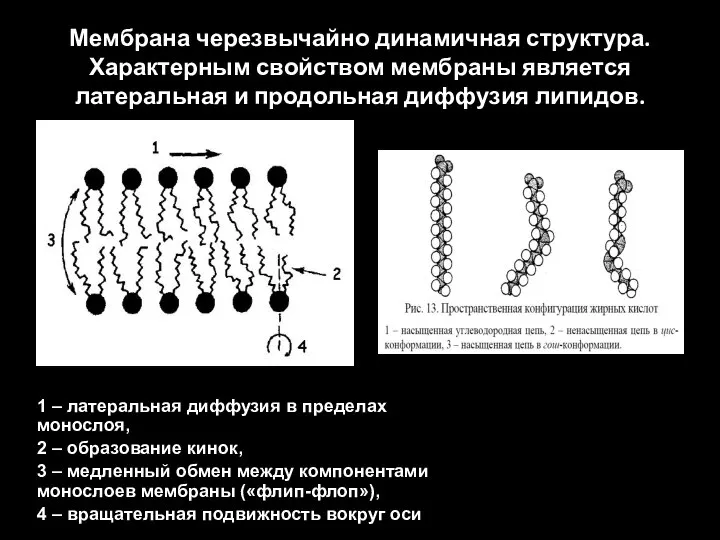
Слайд 35Мембрана черезвычайно динамичная структура. Характерным свойством мембраны является латеральная и продольная диффузия
Мембрана черезвычайно динамичная структура. Характерным свойством мембраны является латеральная и продольная диффузия
Слайд 36
Функциональная классификация мембранных белков:
1. Белки – каналы
Обеспечивают транспорт ионов.
2. Белки – транспортёры
Участвуют
Функциональная классификация мембранных белков:
1. Белки – каналы
Обеспечивают транспорт ионов.
2. Белки – транспортёры
Участвуют

Слайд 37Структурная классификация белков мембраны:
Белки биологических мембран подразделяют на периферические и интегральные
Структурная классификация белков мембраны:
Белки биологических мембран подразделяют на периферические и интегральные

Слайд 38Интегральные мембранные белки встроены в липидный бислой. Их гидрофильные аминокислоты взаимодействуют с
Интегральные мембранные белки встроены в липидный бислой. Их гидрофильные аминокислоты взаимодействуют с

Слайд 39Углеводы
Углеводы в составе мембран обнаруживаются лишь в соединении с белками (гликопротеины и
Углеводы
Углеводы в составе мембран обнаруживаются лишь в соединении с белками (гликопротеины и

Слайд 40Функции углеводов
межклеточное узнавание,
межклеточные взаимодействия,
поддержание иммунного статуса клетки,
обеспечение стабильности белковых молекул в
Функции углеводов
межклеточное узнавание,
межклеточные взаимодействия,
поддержание иммунного статуса клетки,
обеспечение стабильности белковых молекул в

Слайд 41Взаимодействие цитоскелета с гликокаликсом
I – протеогликан,
II – коллаген,
III – фибронектин
Взаимодействие цитоскелета с гликокаликсом
I – протеогликан,
II – коллаген,
III – фибронектин

Слайд 42
Вода
Свободная вода омывает мембрану, заполняет каналы, поры и кинки. Вода может находится
Вода
Свободная вода омывает мембрану, заполняет каналы, поры и кинки. Вода может находится

Слайд 43
Транспортные процессы мембраны обеспечивают:
1. Поддержание объема клетки и внутриклеточного ионного состава в
Транспортные процессы мембраны обеспечивают:
1. Поддержание объема клетки и внутриклеточного ионного состава в

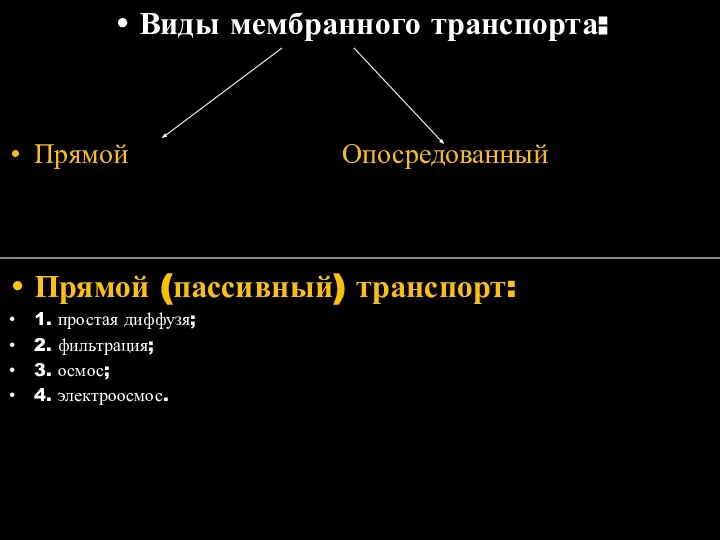
Слайд 44Виды мембранного транспорта:
Прямой Опосредованный
Прямой (пассивный) транспорт:
1. простая диффузя;
2. фильтрация;
3. осмос;
4. электроосмос.
Виды мембранного транспорта:
Прямой Опосредованный
Прямой (пассивный) транспорт:
1. простая диффузя;
2. фильтрация;
3. осмос;
4. электроосмос.
Слайд 45Пассивный транспорт
Идет без затраты энергии АТФ!
Движущие силы:
Градиент концентрации вещества (химический градиент)
Градиент
Пассивный транспорт
Идет без затраты энергии АТФ!
Движущие силы:
Градиент концентрации вещества (химический градиент)
Градиент

Слайд 46Диффузия – самопроизвольный процесс проникновения вещества из области большей концентрации в область

Слайд 47 1. Простая диффузия идет непосредственно
ЧЕРЕЗ МЕМБРАНУ для незаряженных (жирорастворимых) веществ
Таким
1. Простая диффузия идет непосредственно
ЧЕРЕЗ МЕМБРАНУ для незаряженных (жирорастворимых) веществ
Таким

Слайд 48Посредством простой диффузии, через мембрану, переносятся в основном газы и неполярные вещества,
Посредством простой диффузии, через мембрану, переносятся в основном газы и неполярные вещества,

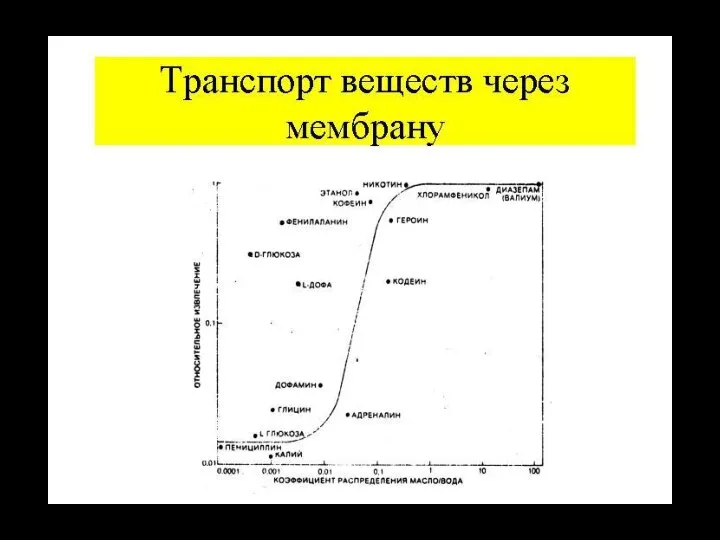
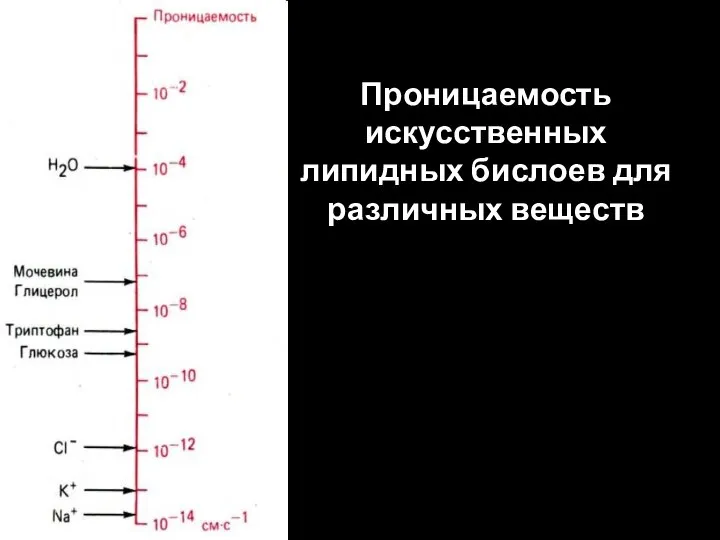
Слайд 50Проницаемость искусственных липидных бислоев для различных веществ
Проницаемость искусственных липидных бислоев для различных веществ
Слайд 51
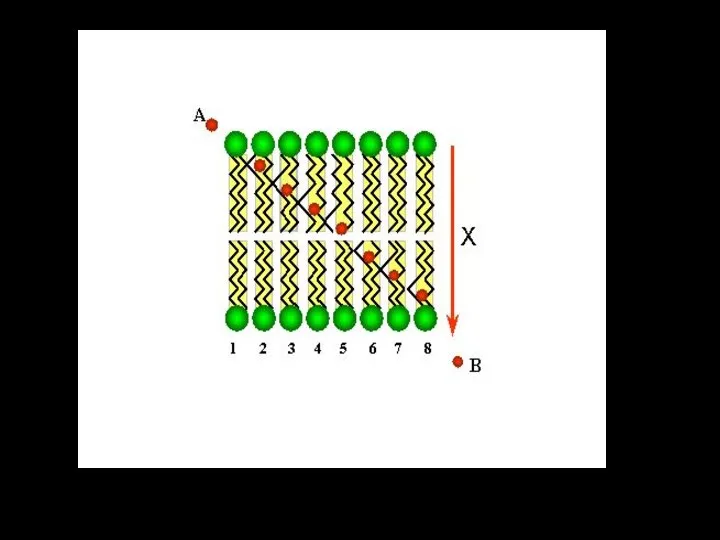
Простая диффузия может осуществляться
ЧЕРЕЗ КИНКИ
При повышении температуры тепловая подвижность жирнокислотных цепей
Простая диффузия может осуществляться
ЧЕРЕЗ КИНКИ
При повышении температуры тепловая подвижность жирнокислотных цепей

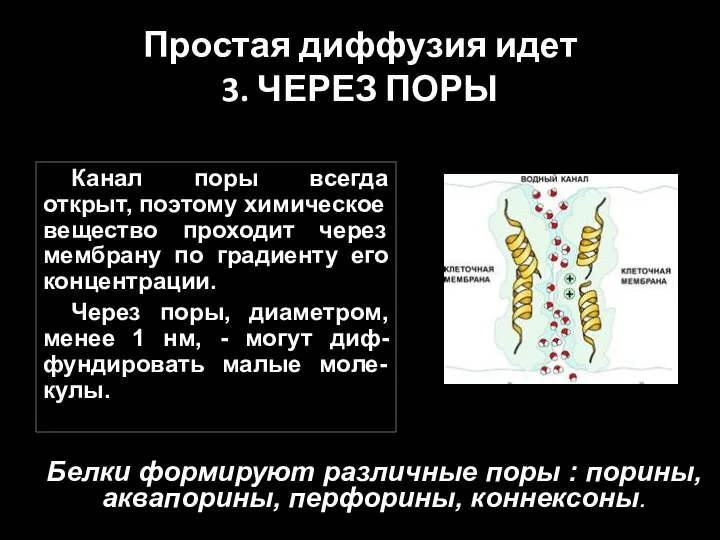
Слайд 53Простая диффузия идет
3. ЧЕРЕЗ ПОРЫ
Канал поры всегда открыт, поэтому химическое вещество
Простая диффузия идет
3. ЧЕРЕЗ ПОРЫ
Канал поры всегда открыт, поэтому химическое вещество
Слайд 54Биомембраны имеют аномально высокую проницаемость для воды.
Это объясняется наличием в мембране
Биомембраны имеют аномально высокую проницаемость для воды.
Это объясняется наличием в мембране

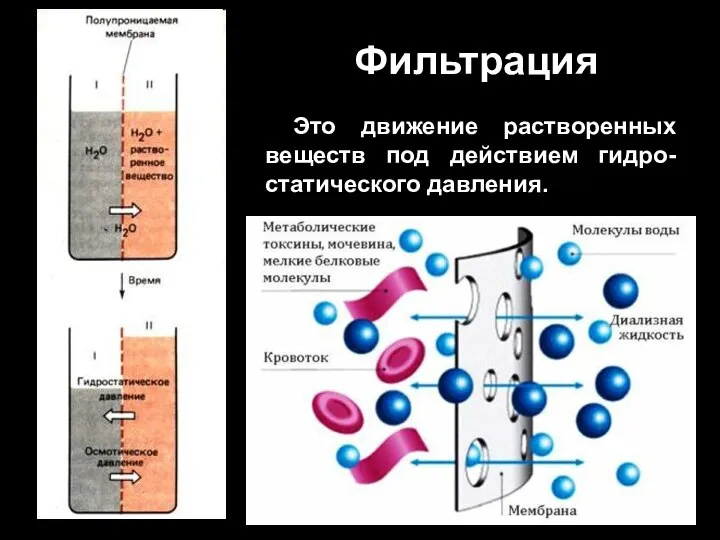
Слайд 55Фильтрация
Это движение растворенных веществ под действием гидро-статического давления.
Фильтрация
Это движение растворенных веществ под действием гидро-статического давления.
Слайд 56Осмос — поток воды через полупроницаемую мембрану из компартмента с меньшей концентрацией растворённых
Осмос — поток воды через полупроницаемую мембрану из компартмента с меньшей концентрацией растворённых

Слайд 57Распределение воды между клеточными компартментами, цитозолем и органоидами клетки, между клеткой и
Распределение воды между клеточными компартментами, цитозолем и органоидами клетки, между клеткой и

Слайд 58Электроосмос
В случае электроосмоса движущей силой является электрохимический градиент.
Электроосмос
В случае электроосмоса движущей силой является электрохимический градиент.

Слайд 59
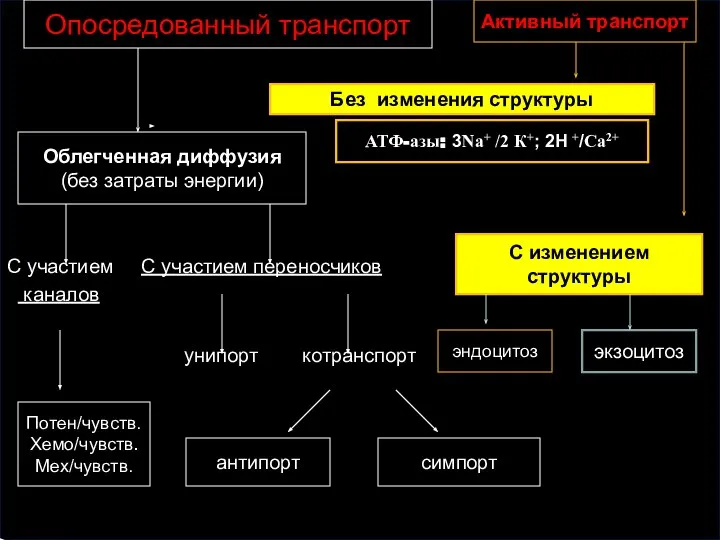
С участием С участием переносчиков
каналов
унипорт котранспорт
Потен/чувств.
Хемо/чувств.
Мех/чувств.
симпорт
Активный транспорт
Облегченная диффузия
(без
С участием С участием переносчиков
каналов
унипорт котранспорт
Потен/чувств.
Хемо/чувств.
Мех/чувств.
симпорт
Активный транспорт
Облегченная диффузия
(без
Слайд 60ОБЛЕГЧЁННАЯ ДИФФУЗИЯ
Для облегчённой диффузии веществ необходимы встроенные в мембрану белковые компоненты (переносчики,
ОБЛЕГЧЁННАЯ ДИФФУЗИЯ
Для облегчённой диффузии веществ необходимы встроенные в мембрану белковые компоненты (переносчики,

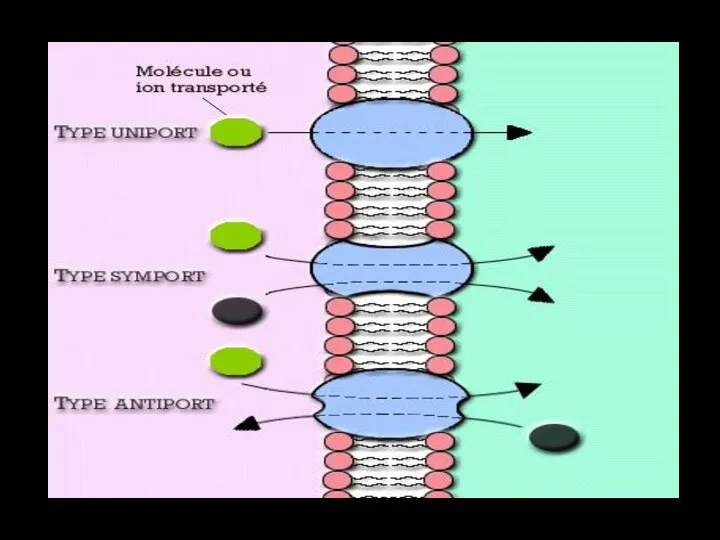
Слайд 61Переносчики
Переносчики (транспортёры) осуществляют транспорт через биологические мембраны множества различных ионов (Na+, Cl–,
Переносчики
Переносчики (транспортёры) осуществляют транспорт через биологические мембраны множества различных ионов (Na+, Cl–,

Слайд 63Симпорт.
Сочетанный транспорт глюкозы и Na+ в тонком кишечнике и канальцах почки
Симпорт.
Сочетанный транспорт глюкозы и Na+ в тонком кишечнике и канальцах почки

Слайд 64Симпорт аминокислот и Na+
Симпорт аминокислот и Na+
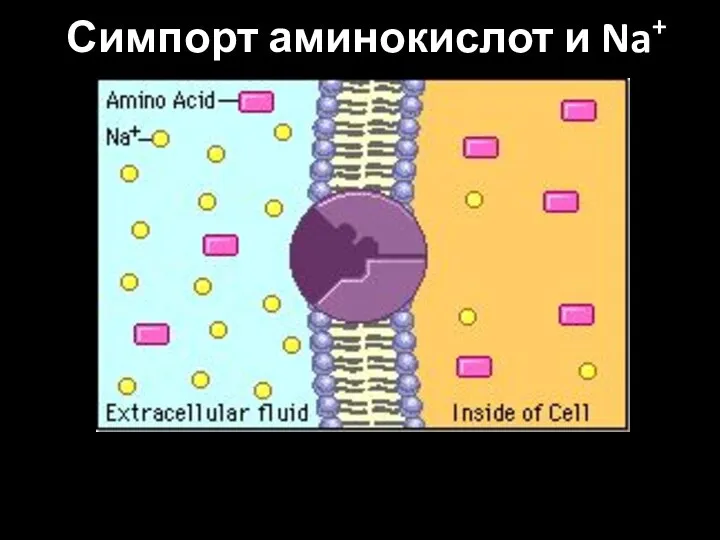
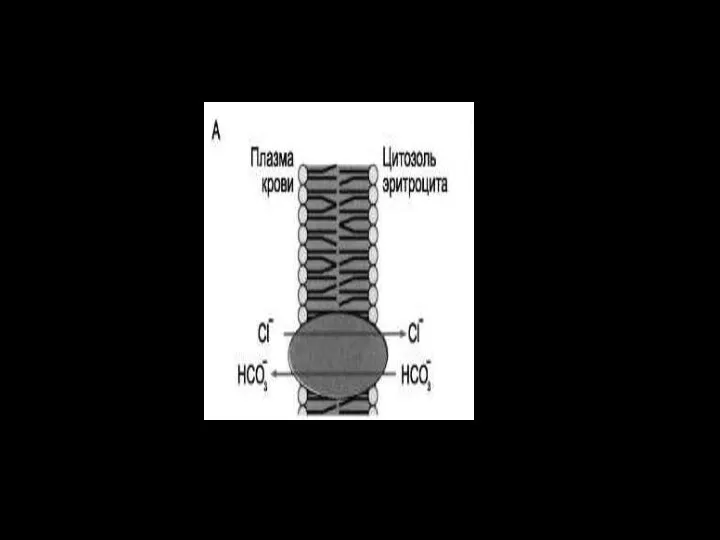
Слайд 65Антипорт:
Катионные обменники :
Nа+/Н+ – обмен, Nа+/Са+–обмен (2Nа/Са)
Анионные обменники :
СL-/НСО3-
Антипорт:
Катионные обменники :
Nа+/Н+ – обмен, Nа+/Са+–обмен (2Nа/Са)
Анионные обменники :
СL-/НСО3-

Слайд 67По механизму действия транспортеры делятся на два типа:
1 тип – совершают челночные
По механизму действия транспортеры делятся на два типа:
1 тип – совершают челночные

Слайд 68Транспорт К+ валиномицином
Транспорт К+ валиномицином
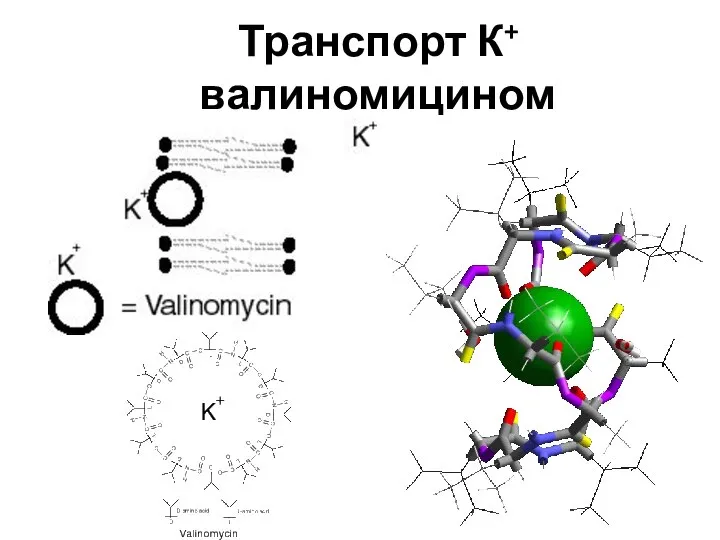
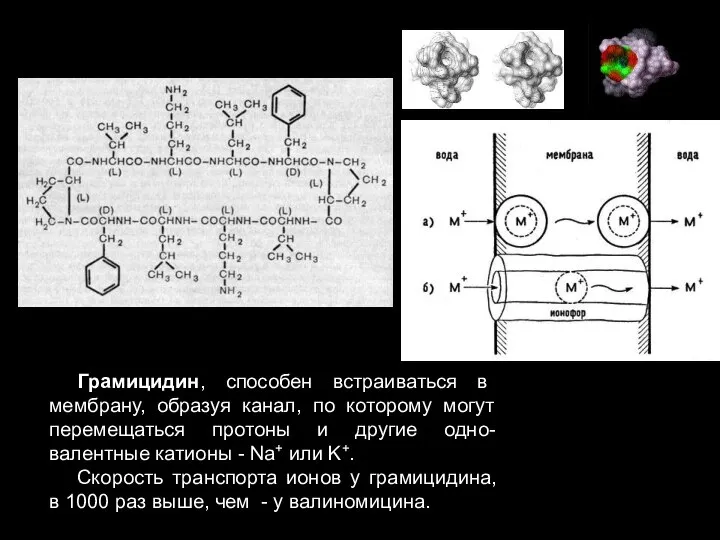
Слайд 69Грамицидин, способен встраиваться в мембрану, образуя канал, по которому могут перемещаться протоны
Грамицидин, способен встраиваться в мембрану, образуя канал, по которому могут перемещаться протоны
Слайд 70 Простая диффузия Облегченная диффузия
обеспечивается работой переносчиков, встроенных в мембрану
Простая диффузия Облегченная диффузия
обеспечивается работой переносчиков, встроенных в мембрану
Слайд 71Ионные каналы – интегральные белки мембраны, выполняющие функции транспорта для соответствующих ионов.
Они
Ионные каналы – интегральные белки мембраны, выполняющие функции транспорта для соответствующих ионов.
Они

Слайд 72Потенциалозависимые ионные каналы управляются мембранным потенциалом (МП).
Колебания МП приводят к конформационным
Потенциалозависимые ионные каналы управляются мембранным потенциалом (МП).
Колебания МП приводят к конформационным

Слайд 73 Строение потенциалозависимого ионного канала:
1 — липидный бислой, 2 — сенсор напряжения,
Строение потенциалозависимого ионного канала: 1 — липидный бислой, 2 — сенсор напряжения,
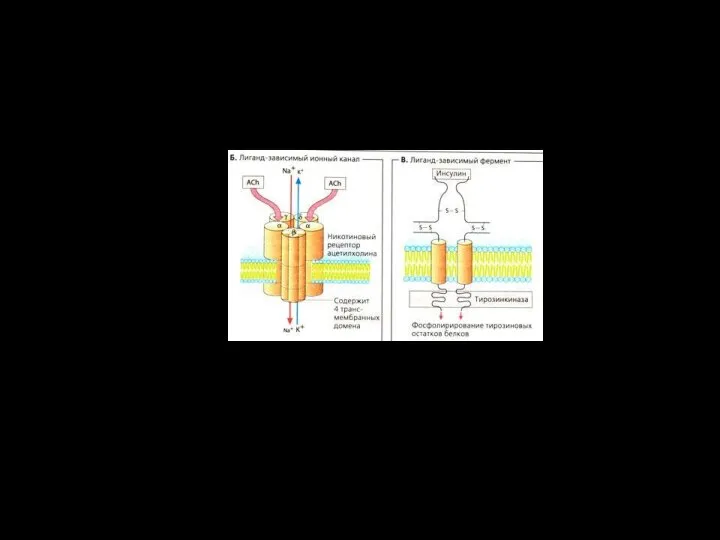
Слайд 74Xемоуправляемые каналы ( лиганд–зависимые ионные каналы, рецептор–зависимые)
Соединение лиганда с рецептором вызывает конформационные
Xемоуправляемые каналы ( лиганд–зависимые ионные каналы, рецептор–зависимые)
Соединение лиганда с рецептором вызывает конформационные

Слайд 76Три вида каналов:
А - ионселективный канал (открытый)
В – хемо-чувствительный канал
С – потенциал-зависимый
Три вида каналов:
А - ионселективный канал (открытый)
В – хемо-чувствительный канал
С – потенциал-зависимый
Слайд 77Активный транспорт
Осуществляется против электрохимического градиента;
Система в высшей степени специфична;
Необходимы источники энергии в
Активный транспорт
Осуществляется против электрохимического градиента;
Система в высшей степени специфична;
Необходимы источники энергии в

Слайд 78Первично активный транспорт
Движущая сила трансмембранного переноса возникает при ферментативном гидролизе макроэргических связей
Первично активный транспорт
Движущая сила трансмембранного переноса возникает при ферментативном гидролизе макроэргических связей

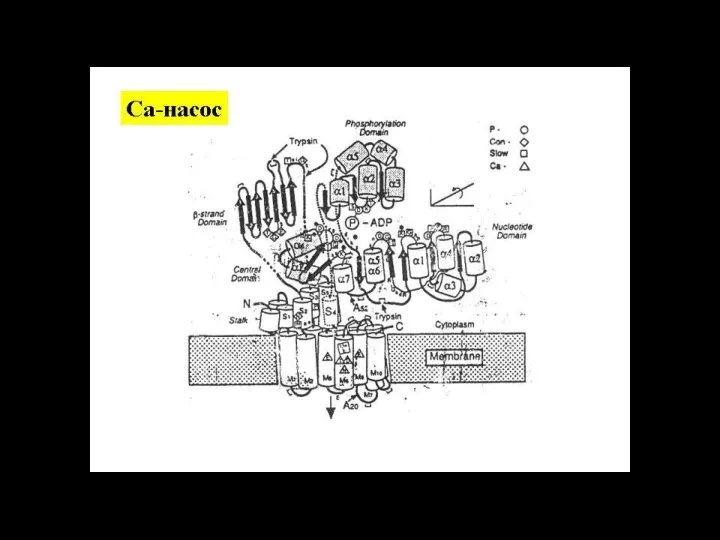
Слайд 79Первично-активный транспорт
Схема Na/K–АТФазы которая за один цикл выносит из клетки три иона
Первично-активный транспорт
Схема Na/K–АТФазы которая за один цикл выносит из клетки три иона
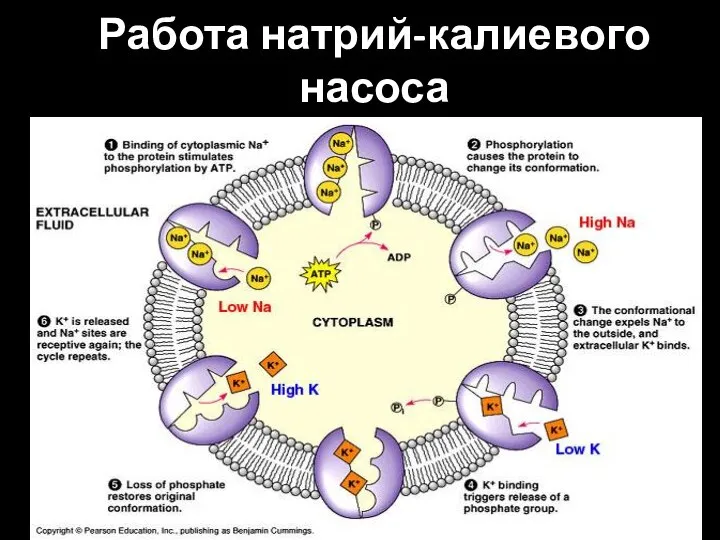
Слайд 80Работа натрий-калиевого насоса
Работа натрий-калиевого насоса
Слайд 81 Вторично активный транспорт.
Движущая сила для трансмембранного переноса одного вещества (или
Вторично активный транспорт.
Движущая сила для трансмембранного переноса одного вещества (или

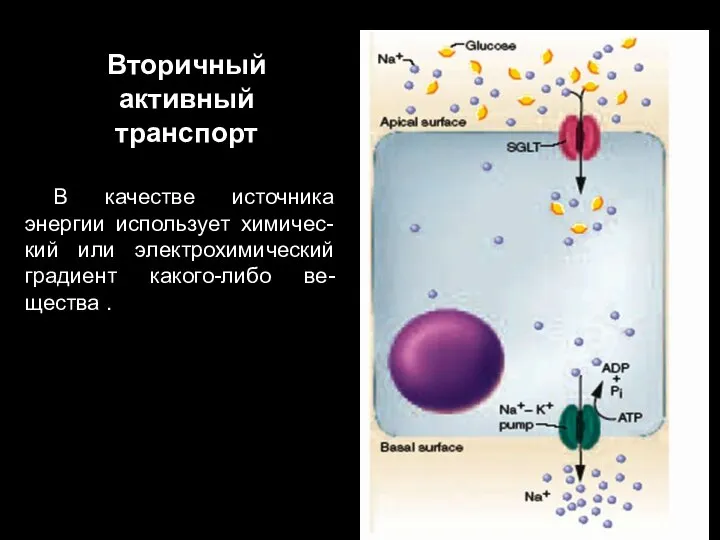
Слайд 82Вторичный активный транспорт
В качестве источника энергии использует химичес-кий или электрохимический градиент какого-либо
Вторичный активный транспорт
В качестве источника энергии использует химичес-кий или электрохимический градиент какого-либо
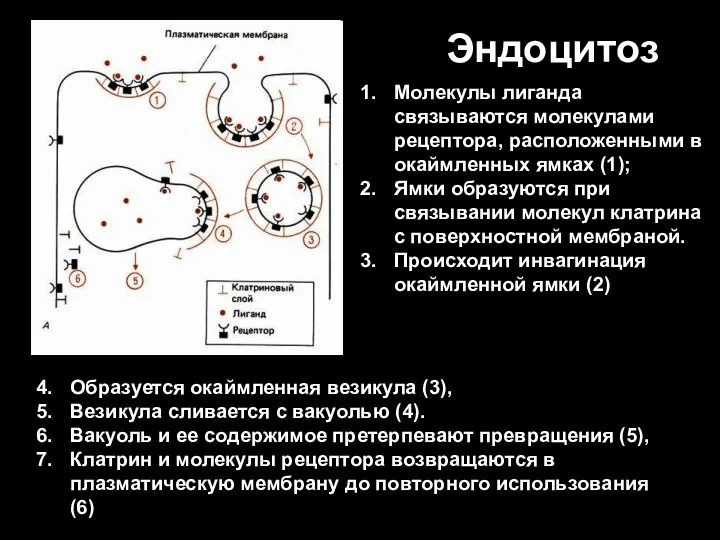
Слайд 83Эндоцитоз
Образуется окаймленная везикула (3),
Везикула сливается с вакуолью (4).
Вакуоль и ее содержимое
Эндоцитоз
Образуется окаймленная везикула (3),
Везикула сливается с вакуолью (4).
Вакуоль и ее содержимое
Слайд 84Экзоцитоз
В аппарате Гольджи из предшественника образуется конечный секрет;
Везикула с секретом доставляется к
Экзоцитоз
В аппарате Гольджи из предшественника образуется конечный секрет;
Везикула с секретом доставляется к
Слайд 85Цитоскелет клетки
Цитоскелет клетки
Слайд 87Динеин и кинезин обеспечивают транспорт крупных молекул и органоидов.
Динеин – ретроградно
Динеин и кинезин обеспечивают транспорт крупных молекул и органоидов. Динеин – ретроградно
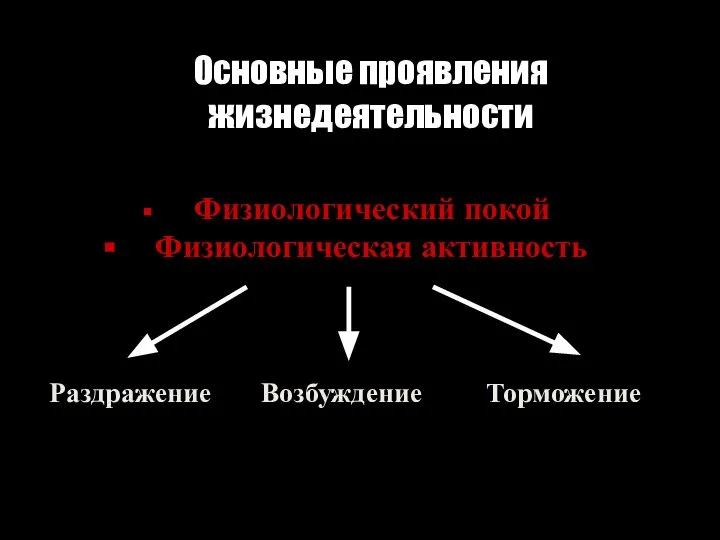
Слайд 89Основные проявления
жизнедеятельности
Физиологический покой
Физиологическая активность
Раздражение Возбуждение Торможение
Основные проявления
жизнедеятельности
Физиологический покой
Физиологическая активность
Раздражение Возбуждение Торможение
Слайд 90Общая характеристика возбудимых тканей.
Все клетки нашего организма обладают свойством раздражимости.
Раздражимость –
Общая характеристика возбудимых тканей.
Все клетки нашего организма обладают свойством раздражимости.
Раздражимость –

Слайд 91Раздражение и виды раздражителей.
Раздражение – это процесс воздействия на живой объект внешних
Раздражение и виды раздражителей.
Раздражение – это процесс воздействия на живой объект внешних

Слайд 92По природе раздражителя:
1. физические ( температура, звук, свет, электрический ток и т.д.).
2.
По природе раздражителя:
1. физические ( температура, звук, свет, электрический ток и т.д.).
2.

Слайд 93По биологическому значению:
1. адекватные – раздражитель к которому в естественных условиях у
1. адекватные – раздражитель к которому в естественных условиях у

Слайд 94По количественному признаку или порогу раздражения:
В зависимости от силы действия раздражители делятся
В зависимости от силы действия раздражители делятся

Слайд 95Все возбудимые ткани обладают
общими свойствами:
Возбудимостью
Проводимостью
Лабильностью
Аккомодацией
Все возбудимые ткани обладают
общими свойствами:
Возбудимостью
Проводимостью
Лабильностью
Аккомодацией

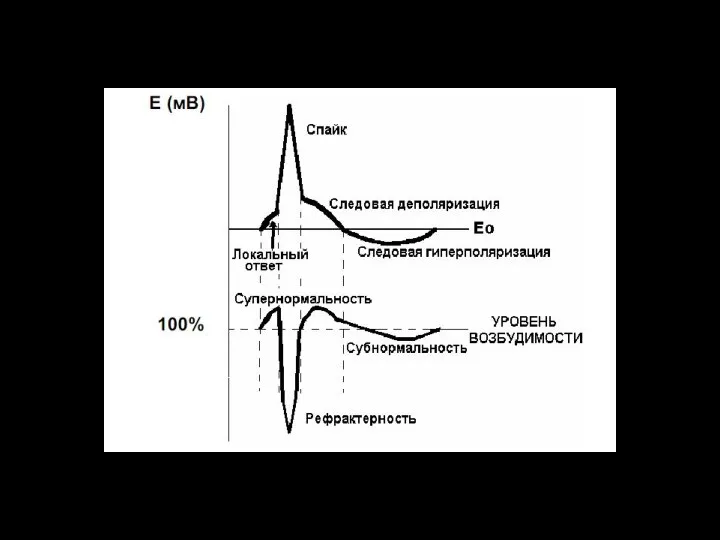
Слайд 96Возбудимость количественно характеризуется порогом возбуждения, который определяется минимальной силой раздражения, способной вызывать

Слайд 97Законы раздражения:
Закон силы
Закон времени
Закон крутизны нарастания раздражителя
Законы раздражения:
Закон силы
Закон времени
Закон крутизны нарастания раздражителя

Слайд 98Закон силы
Чем сильней раздражение, тем до определенного предела сильнее ответная реакция биосистемы.
Закон
Закон силы
Чем сильней раздражение, тем до определенного предела сильнее ответная реакция биосистемы.
Закон

Слайд 99Закон времени
Чем длительнее раздражение, тем сильней до определенных пределов ответная реакция
Закон времени
Чем длительнее раздражение, тем сильней до определенных пределов ответная реакция

Слайд 100ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ»
Этот закон был экспериментально установллен и сформулирован независимо друг
ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ»
Этот закон был экспериментально установллен и сформулирован независимо друг

Слайд 102Закон крутизны нарастания раздражителя или градиента:
Чем выше крутизна нарастания раздражителя во времени,
Закон крутизны нарастания раздражителя или градиента:
Чем выше крутизна нарастания раздражителя во времени,

Слайд 103Спасибо за внимание
Спасибо за внимание

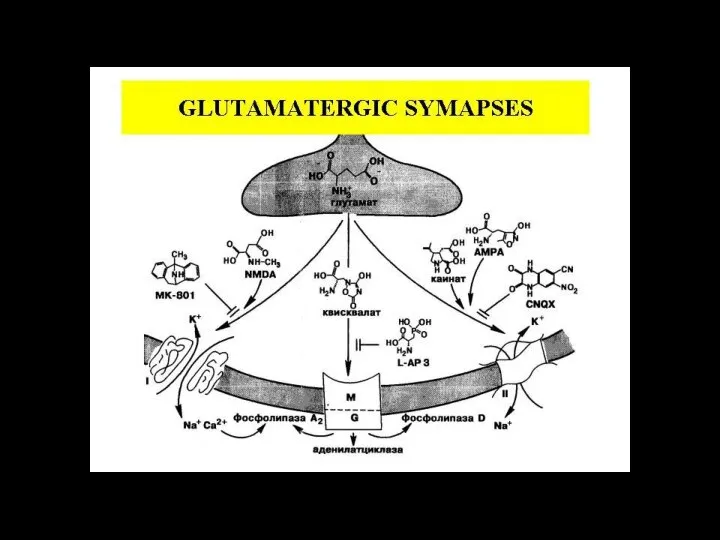
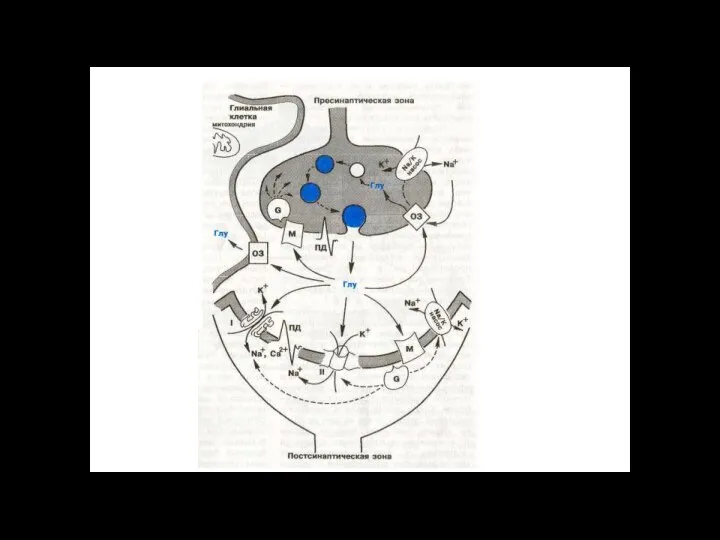
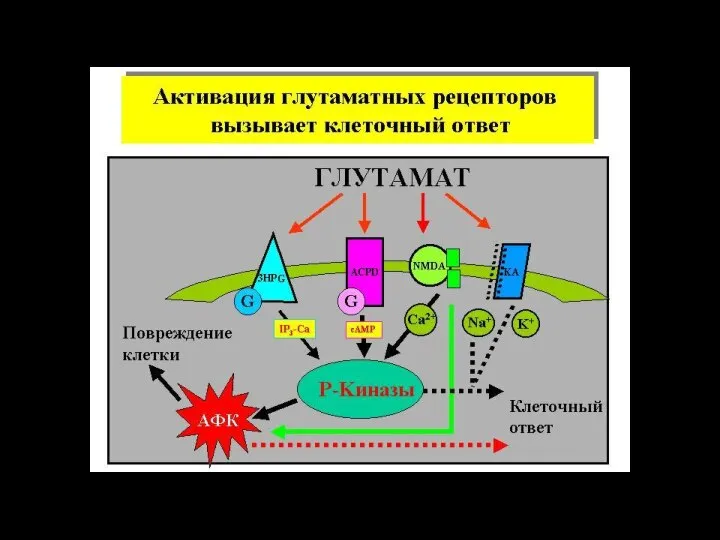
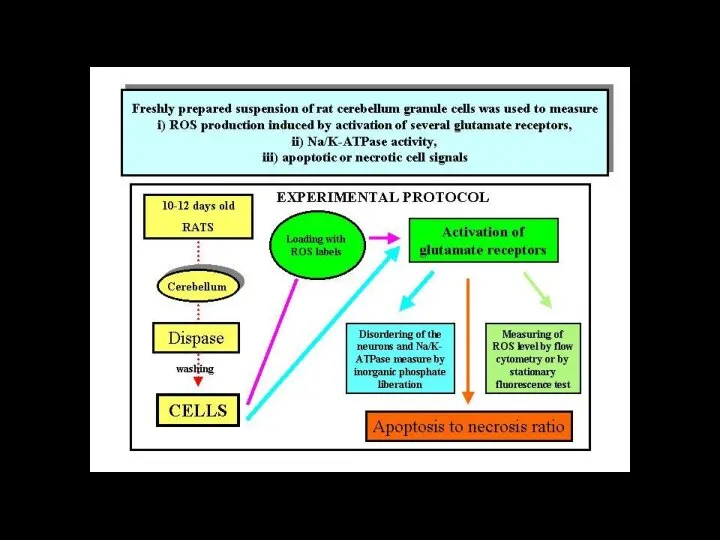

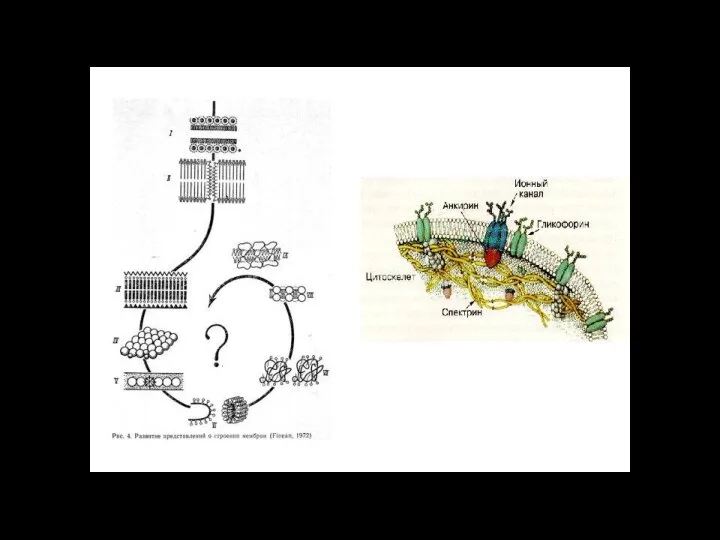
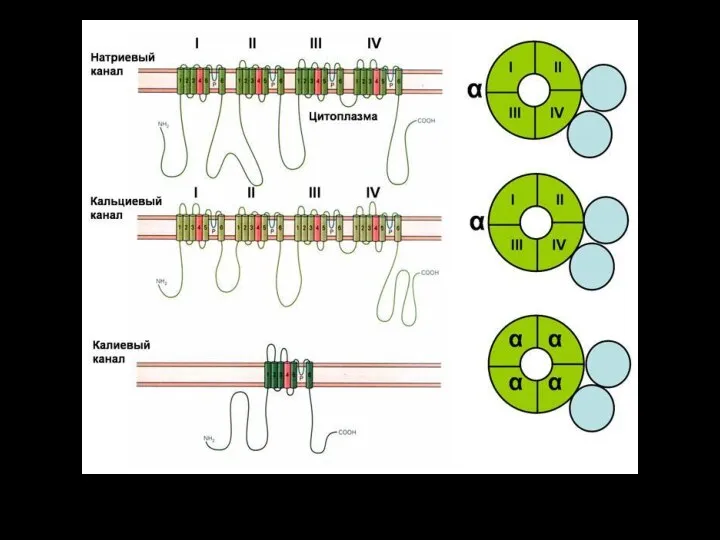
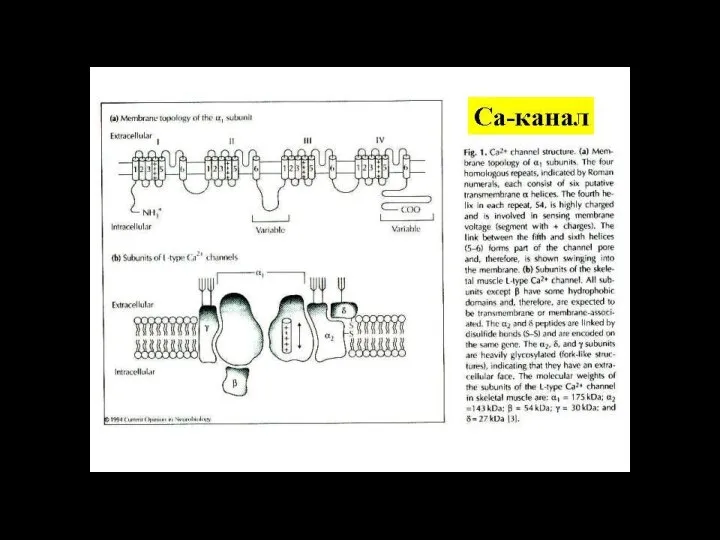
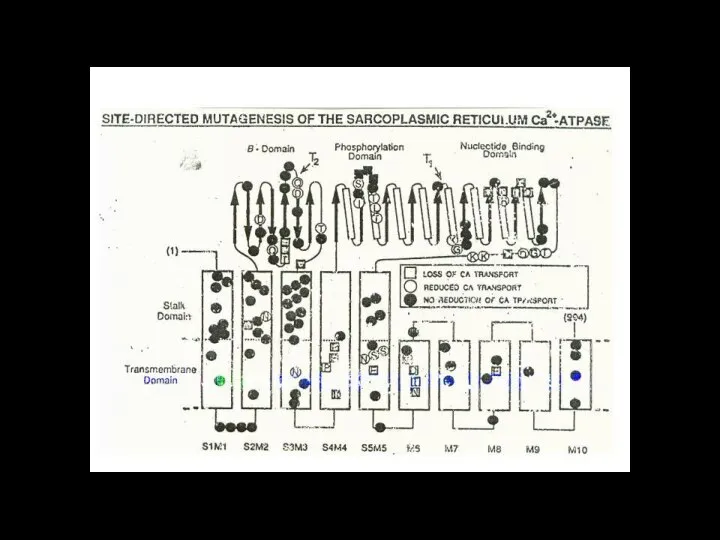
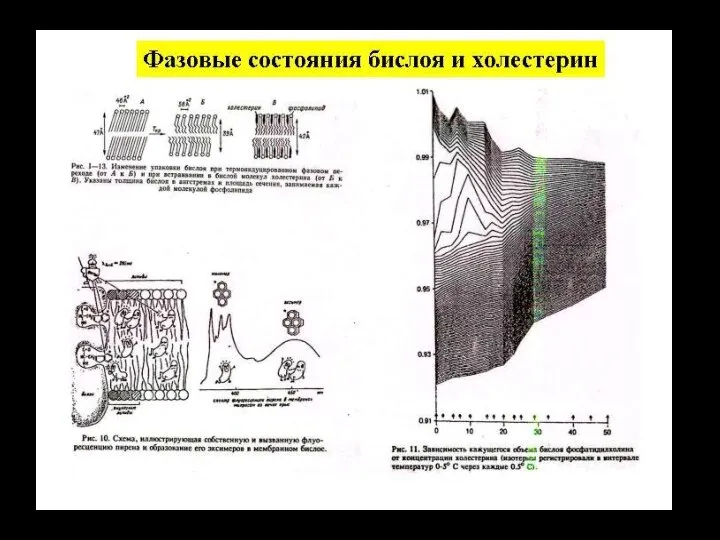

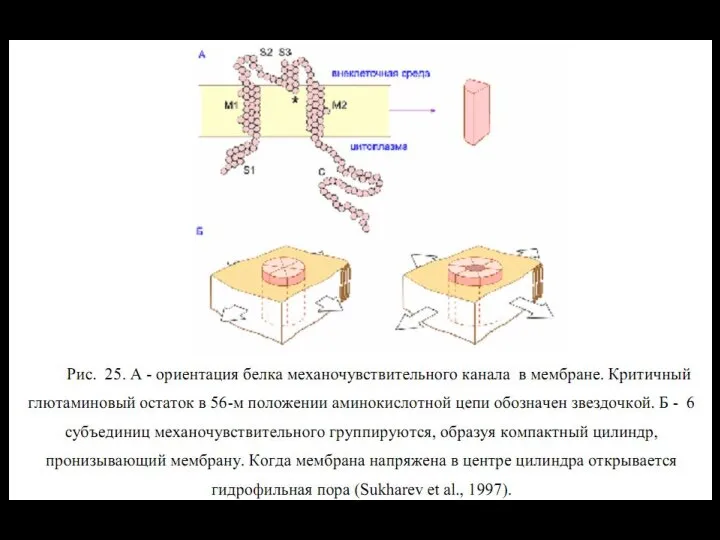
Слайд 116ионный канал
состоит из связанных между собой белковых субъединиц, формирующих в мембране
ионный канал
состоит из связанных между собой белковых субъединиц, формирующих в мембране
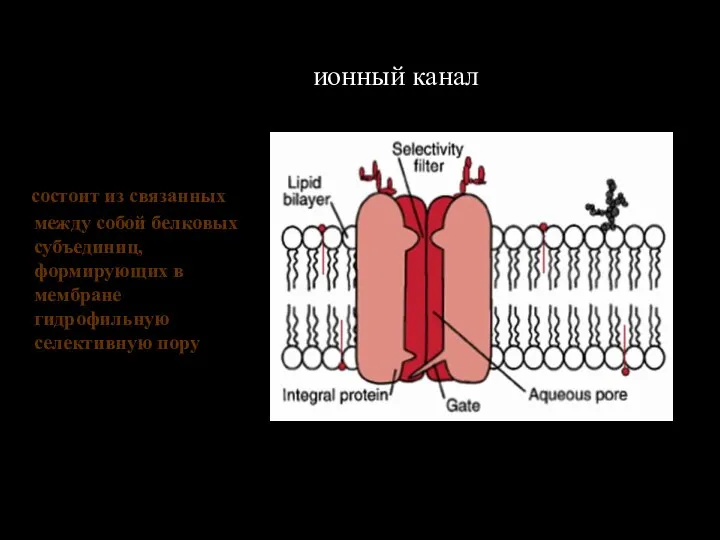
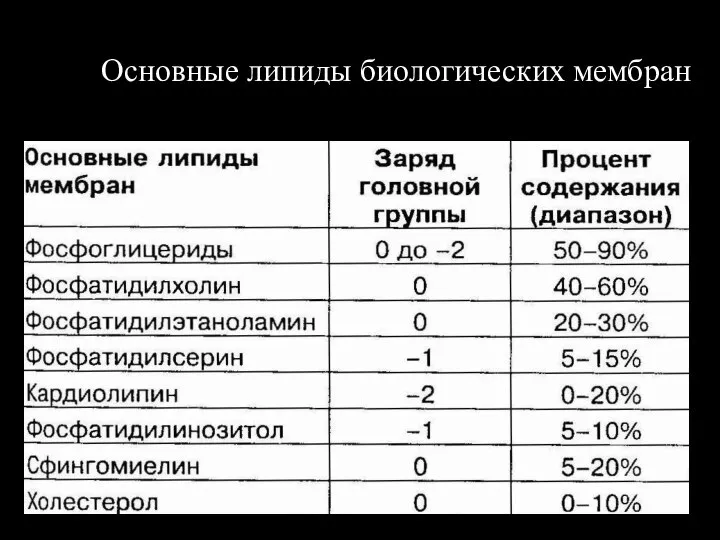
Слайд 117Основные липиды биологических мембран
Основные липиды биологических мембран
Слайд 121СПОСОБЫ УПАКОВКИ ФОСФОЛИПИДОВ
СПОСОБЫ УПАКОВКИ ФОСФОЛИПИДОВ

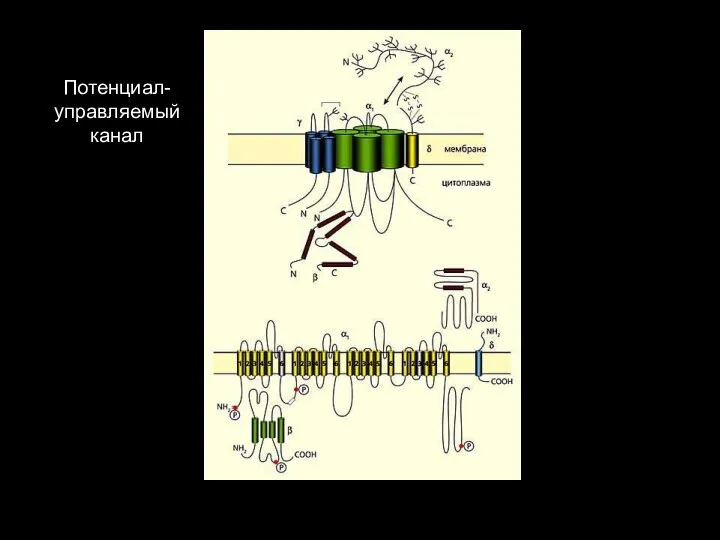
Слайд 125Потенциал-управляемый канал
Потенциал-управляемый канал

Слайд 129Цикл активации G-белка под действием G-белок-связанного рецептора.
Цикл активации G-белка под действием G-белок-связанного рецептора.


Слайд 146Основные липиды биологических мембран
Основные липиды биологических мембран
 Зоофиты (задача 7)
Зоофиты (задача 7) Бактерии, грибы, лишайники
Бактерии, грибы, лишайники Турако Левингстона — птица средних размеров
Турако Левингстона — птица средних размеров Молекулярный уровень. Химическая организация клетки
Молекулярный уровень. Химическая организация клетки Кровообращение
Кровообращение Виды декоративных мышей
Виды декоративных мышей Координация и регуляция
Координация и регуляция Наследственность и изменчивость организмов
Наследственность и изменчивость организмов Белки. Функции белков
Белки. Функции белков Морфологический критерий вида. Сходство внешнего и внутреннего строения особей
Морфологический критерий вида. Сходство внешнего и внутреннего строения особей Класс насекомые
Класс насекомые Работа с графической информацией на ВПР по биологии. Задание 6, 5 класс
Работа с графической информацией на ВПР по биологии. Задание 6, 5 класс Семейства цветковых растений
Семейства цветковых растений Генриетта Яковлевна Городисская
Генриетта Яковлевна Городисская Структуры головного мозга
Структуры головного мозга Заповідники. Ботанічні сади
Заповідники. Ботанічні сади Презентация на тему КАК КЛАССИФИЦИРУЮТ ЖИВЫЕ ОРГАНИЗМЫ
Презентация на тему КАК КЛАССИФИЦИРУЮТ ЖИВЫЕ ОРГАНИЗМЫ  Unusual animals in the Mariana Trench
Unusual animals in the Mariana Trench Выбор щенка
Выбор щенка Акантоцефалезы (Тест)
Акантоцефалезы (Тест) Животный мир водоёма
Животный мир водоёма Муравьиная жизнь
Муравьиная жизнь Основная структурно-функциональная единица всех живых организмов
Основная структурно-функциональная единица всех живых организмов Биосинтез белка
Биосинтез белка Хомяки в домашних условиях
Хомяки в домашних условиях Исчезающие рептилии
Исчезающие рептилии Ethos & logos
Ethos & logos Садовые цветы
Садовые цветы