- Биосинтез белка

Содержание
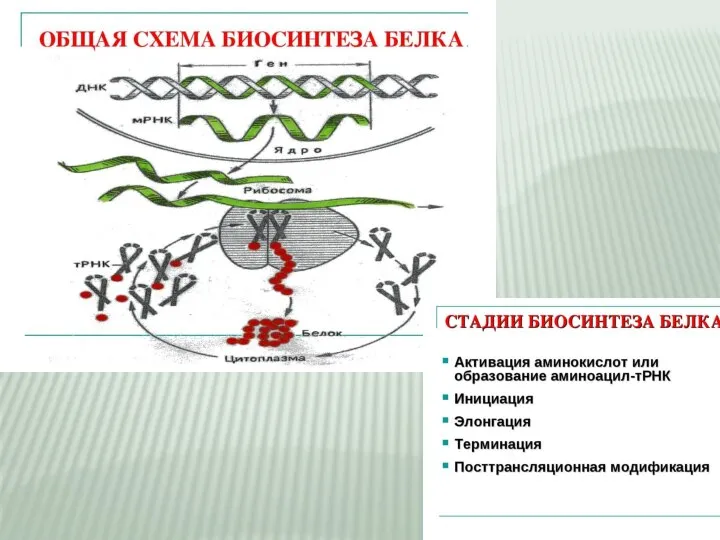
- 2. Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток
- 4. Биосинтез белка ДНК → иРНК → белок
- 6. Биосинтез белка рибосома АУГ – УАЦ – ГЦЦ – АГЦ …. и – РНК т –
- 8. "Трилистник" т-РНК
- 9. Стадии трансляции м – РНК: АУГ ААГ ЦГУ ГГЦ Затем происходит присоединение большой субъединицы рибосомы *Целостная
- 10. Рибосомы представляют собой рибонуклеопротеиновые образования. На рибосомах идёт сборка аминокислот в белки. Белки, входящие в состав
- 11. Функциональный центр рибосомы – ФЦР (два триплета) А аминокислотный центр центр узнавания аминокислоты Р пептидный центр
- 12. Стадии трансляции м – РНК: АУГ – ААГ – ЦГУ – ГГЦ … 2. Элонгация -
- 13. ОбАминокислота взаимодействует с АТФ и активируется, образуя образование аминоацил-тРНК аминоациладенилат, который, не освобождаясь из связи с
- 14. Стадии трансляции 3.Терминация – окончание биосинтеза На стоп-кодонах синтез полипептида прекращается Рибосома вновь разделяется на субъединицы
- 15. В ходе синтеза белка прочтение информации мРНК идёт в направлении от 5'- к З'-концу, обеспечивая синтез
- 17. Стадии трансляции Полисома – молекула и-РНК, на которой находятся несколько рибосом, синтезирующих одинаковые белки
- 18. Регуляция синтеза белка
- 19. АТФ и ГТФ как источники энергии На включение одной аминокислоты в растущую полипептидную цепь клетка затрачивает
- 20. Посттрансляционные изменения - конформационные и структурные изменения полипептидных цепей. Полипептидные цепи могут подвергаться структурным модификациям будучи
- 21. Посттрансляционные изменения - конформационные и структурные изменения полипептидных цепей. Полипептидные цепи могут подвергаться структурным модификациям будучи
- 22. КОВАЛЕНТНЫЕ МОДИФИКАЦИИ Активирование или инактивирование структурных белков и ферментов может происходить в результате присоединения различных химических
- 23. Генетический код и его свойства Генетический, биологический, нуклеотидный, или аминокислотный код - своеобразный "словарь", позволяющий выяснить,
- 24. Свойства генетического кода Триплетность Число кодирующих последовательностей из четырёх нуклеотидов по три равно 43 = 64.
- 25. Вырожденность В информационных молекулах включение в белок одной и той же аминокислоты определяют несколько кодонов. Линейность
- 26. Основные компоненты белоксинтезирующей системы
- 28. Скачать презентацию
Слайд 2Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков,
Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков,


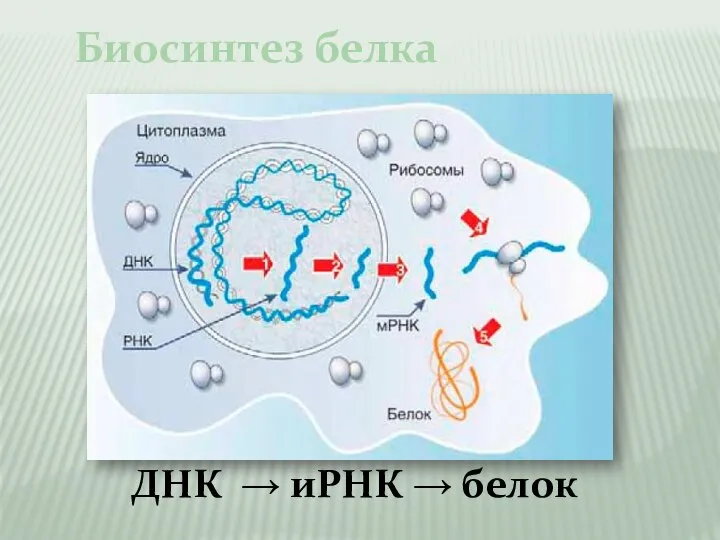
Слайд 4Биосинтез белка
ДНК → иРНК → белок
Биосинтез белка
ДНК → иРНК → белок
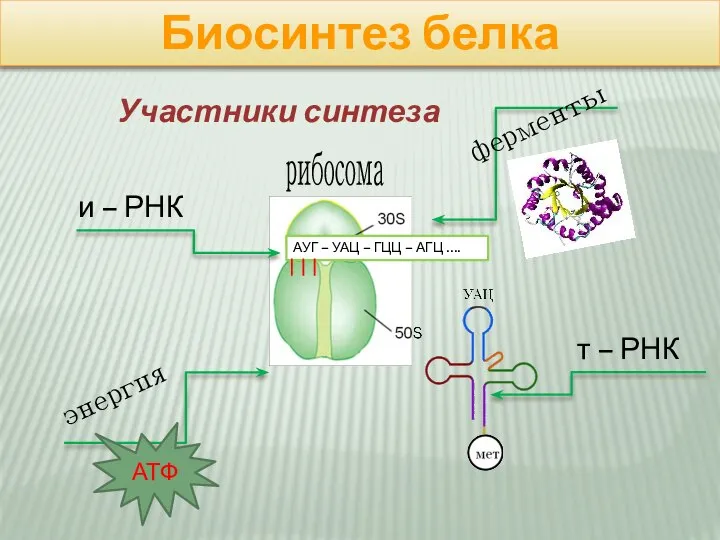
Слайд 6Биосинтез белка
рибосома
АУГ – УАЦ – ГЦЦ – АГЦ ….
и – РНК
Биосинтез белка
рибосома
АУГ – УАЦ – ГЦЦ – АГЦ ….
и – РНК
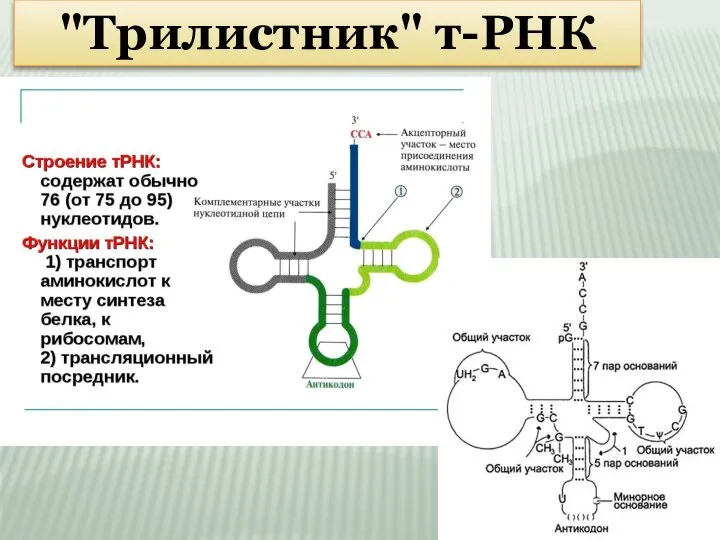
Слайд 8"Трилистник" т-РНК
"Трилистник" т-РНК
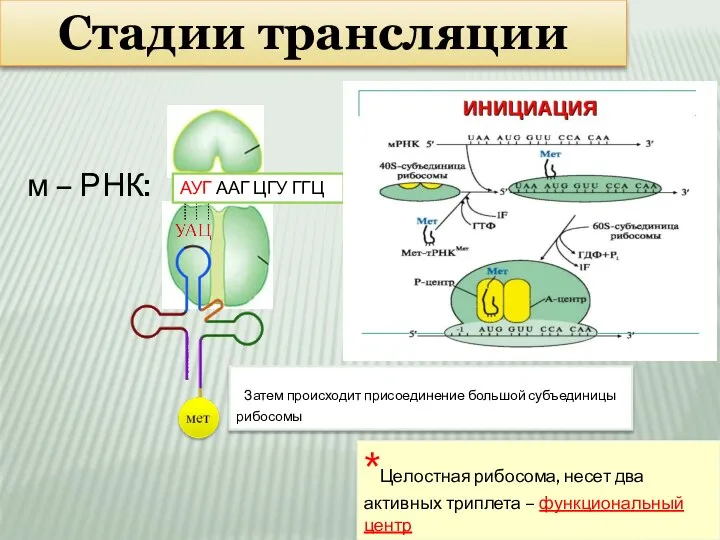
Слайд 9Стадии трансляции
м – РНК:
АУГ ААГ ЦГУ ГГЦ
Затем происходит присоединение большой
Стадии трансляции
м – РНК:
АУГ ААГ ЦГУ ГГЦ
Затем происходит присоединение большой
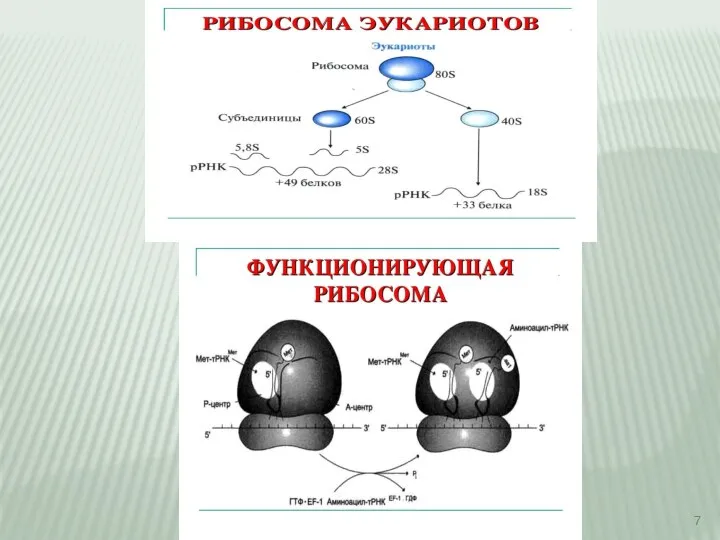
Слайд 10Рибосомы представляют собой рибонуклеопротеиновые образования. На рибосомах идёт сборка аминокислот в белки.
Рибосомы представляют собой рибонуклеопротеиновые образования. На рибосомах идёт сборка аминокислот в белки.

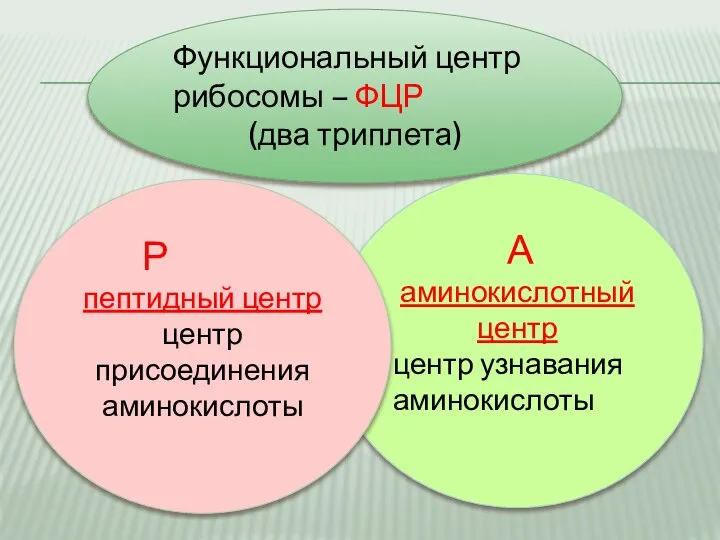
Слайд 11Функциональный центр рибосомы – ФЦР
(два триплета)
А аминокислотный центр
центр узнавания аминокислоты
Функциональный центр рибосомы – ФЦР
(два триплета)
А аминокислотный центр
центр узнавания аминокислоты
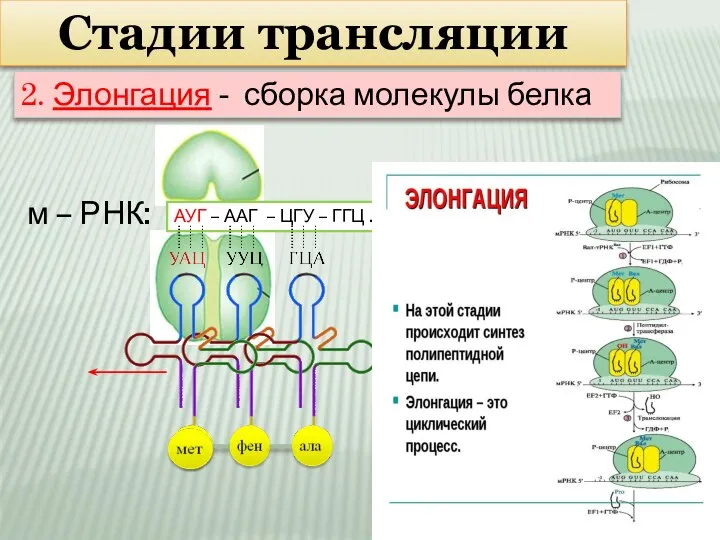
Слайд 12Стадии трансляции
м – РНК:
АУГ – ААГ – ЦГУ – ГГЦ …
Стадии трансляции
м – РНК:
АУГ – ААГ – ЦГУ – ГГЦ …
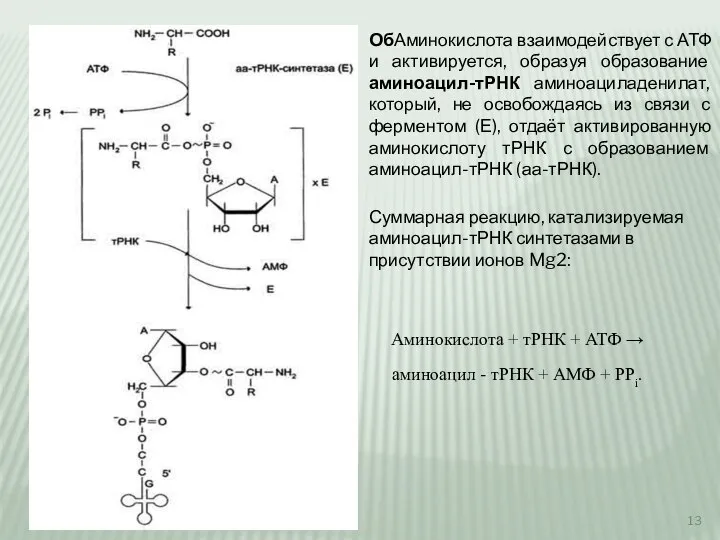
Слайд 13ОбАминокислота взаимодействует с АТФ и активируется, образуя образование аминоацил-тРНК аминоациладенилат, который, не
ОбАминокислота взаимодействует с АТФ и активируется, образуя образование аминоацил-тРНК аминоациладенилат, который, не
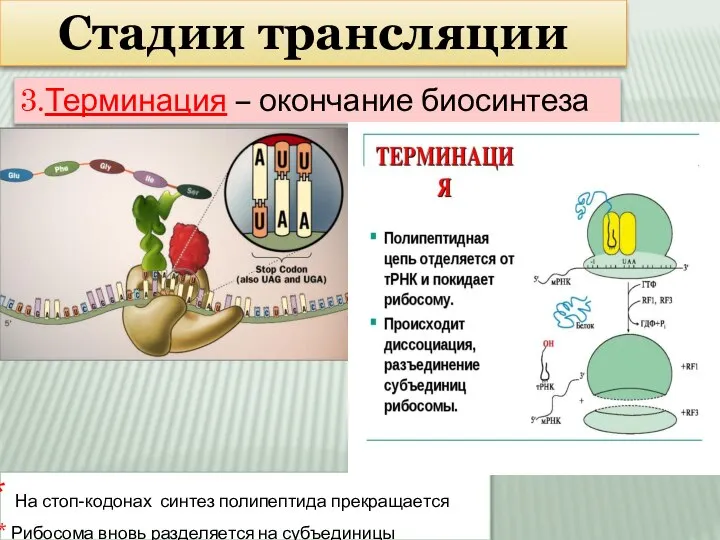
Слайд 14Стадии трансляции
3.Терминация – окончание биосинтеза
На стоп-кодонах синтез полипептида прекращается
Рибосома вновь
Стадии трансляции
3.Терминация – окончание биосинтеза
На стоп-кодонах синтез полипептида прекращается
Рибосома вновь
Слайд 15В ходе синтеза белка прочтение информации мРНК идёт в направлении от 5'-
В ходе синтеза белка прочтение информации мРНК идёт в направлении от 5'-


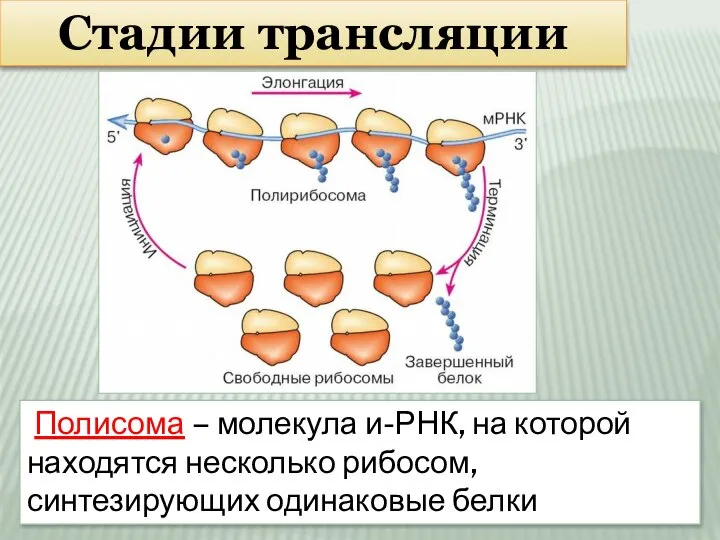
Слайд 17Стадии трансляции
Полисома – молекула и-РНК, на которой находятся несколько рибосом, синтезирующих
Стадии трансляции
Полисома – молекула и-РНК, на которой находятся несколько рибосом, синтезирующих
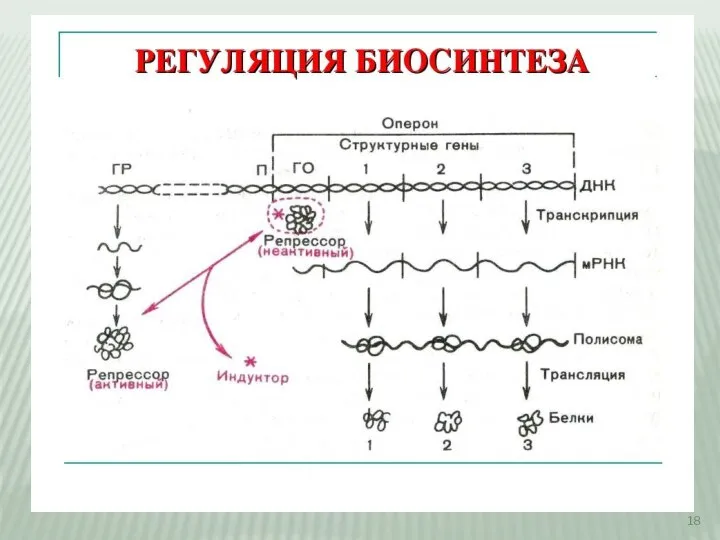
Слайд 18Регуляция синтеза белка
Регуляция синтеза белка
Слайд 19АТФ и ГТФ как источники энергии
На включение одной аминокислоты в растущую полипептидную
АТФ и ГТФ как источники энергии
На включение одной аминокислоты в растущую полипептидную

Слайд 20Посттрансляционные изменения - конформационные и структурные изменения полипептидных цепей.
Полипептидные цепи могут подвергаться
Посттрансляционные изменения - конформационные и структурные изменения полипептидных цепей.
Полипептидные цепи могут подвергаться

Слайд 21Посттрансляционные изменения - конформационные и структурные изменения полипептидных цепей.
Полипептидные цепи могут подвергаться
Посттрансляционные изменения - конформационные и структурные изменения полипептидных цепей.
Полипептидные цепи могут подвергаться

Слайд 22КОВАЛЕНТНЫЕ МОДИФИКАЦИИ
Активирование или инактивирование структурных белков и ферментов может происходить в результате
КОВАЛЕНТНЫЕ МОДИФИКАЦИИ
Активирование или инактивирование структурных белков и ферментов может происходить в результате

Слайд 23 Генетический код и его свойства
Генетический, биологический, нуклеотидный, или аминокислотный код -
Генетический код и его свойства
Генетический, биологический, нуклеотидный, или аминокислотный код -

Слайд 24Свойства генетического кода
Триплетность
Число кодирующих последовательностей из четырёх нуклеотидов по три равно 43 =
Свойства генетического кода
Триплетность
Число кодирующих последовательностей из четырёх нуклеотидов по три равно 43 =

Слайд 25Вырожденность
В информационных молекулах включение в белок одной и той же аминокислоты определяют
Вырожденность
В информационных молекулах включение в белок одной и той же аминокислоты определяют

Слайд 26Основные компоненты белоксинтезирующей системы
Основные компоненты белоксинтезирующей системы

 Презентация на тему Критерии вида
Презентация на тему Критерии вида  Влияние различных условий (температура, pH) на структуру белков. Лабораторная работа 2
Влияние различных условий (температура, pH) на структуру белков. Лабораторная работа 2 Семейство розоцветные. Класс Двудольные
Семейство розоцветные. Класс Двудольные Prezentatsia_Organicheskie_veschestva_Uglevody_Belki
Prezentatsia_Organicheskie_veschestva_Uglevody_Belki Первичная структура белка. Зависимость биологических свойств белков от первичной структуры. Вторичная структура белка
Первичная структура белка. Зависимость биологических свойств белков от первичной структуры. Вторичная структура белка Красная книга. Dactylorhiza traunsteineri (пальцекорник траунштейнера)
Красная книга. Dactylorhiza traunsteineri (пальцекорник траунштейнера) Эти удивительные собаки…
Эти удивительные собаки… Формы размножения организмов и их цитологические основы
Формы размножения организмов и их цитологические основы Презентация _Строение гена прокариот и эукариот _
Презентация _Строение гена прокариот и эукариот _ Простейшие
Простейшие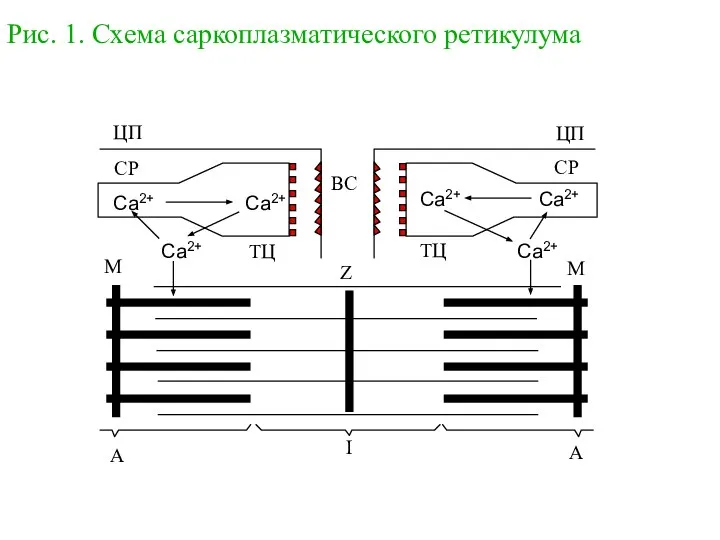 Саркоплазматический ретикулум. Работа Са-АТФазы
Саркоплазматический ретикулум. Работа Са-АТФазы Первоцветы. Кроссворд
Первоцветы. Кроссворд Жизнь растений пресного водоема
Жизнь растений пресного водоема Анализаторы (органы чувств), их строения и функции, зрительный анализатор
Анализаторы (органы чувств), их строения и функции, зрительный анализатор Jean Batiste Lamarck
Jean Batiste Lamarck Экокормушка МБОУ СОШ№25
Экокормушка МБОУ СОШ№25 Игра – зачет по теме : Тип Хордовые. 7 класс
Игра – зачет по теме : Тип Хордовые. 7 класс portfoli
portfoli Шаблон. Творческая работа по биологии
Шаблон. Творческая работа по биологии Дикие животные
Дикие животные Генетический код. Первичное действие гена
Генетический код. Первичное действие гена Строение и функции пищеварительной системы
Строение и функции пищеварительной системы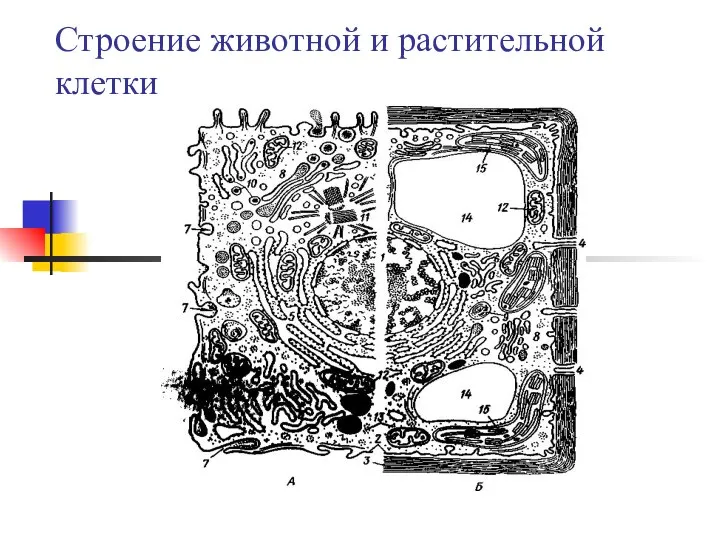 Строение животной и растительной клетки
Строение животной и растительной клетки Организация мониторинговых исследований после пожара на участке Буртинская степь заповедника Оренбургский
Организация мониторинговых исследований после пожара на участке Буртинская степь заповедника Оренбургский Глаз как оптическая система
Глаз как оптическая система Ракообразные. Раки и Крабы
Ракообразные. Раки и Крабы Производство инсулина биотехнологическим способом
Производство инсулина биотехнологическим способом Нут – бараний или турецкий горох
Нут – бараний или турецкий горох