Содержание
- 2. Онтогенез, его типы и периодизация Онтогенез (от греч. ontos — существо, genesis — развитие) — это
- 3. Тех, кто отдавал предпочтение яйцеклеткам, называли овистами, а тех, кто большое значение придавал мужским половым клеткам,
- 4. В свою очередь, для объяснения «механизма» образования зародыша в половой клетке использовалась теория «вложения», в соответствии
- 5. Ланкестер (L. R. Lankester), Уитмен (Ch. Whitman), Раб ль (K. Rabl) и другие эмбриологи выдвигают ряд
- 6. Эпигенез ЭПИГЕНЕЗ (греческий epi- на, после, вслед за + genesis зарождение, происхождение) — учение о зародышевом
- 7. Попытку разработать механическую теорию эпигенеза предпринял Р. Декарт, полагавший, что движущей силой развития является «теплота», выделяющаяся
- 8. Согласно современным представлениям, основные закономерности развития определяются генетической информацией, содержащейся в ДНК родительских половых клеток. Формирование
- 9. Понятия о росте организмов (размножении клеток), дифферен-цировке клеток и о морфогенезе позволяют сформулировать заключение о развитии
- 10. Периоды Проэмбриональный (от греч. pro — до, embryon — зародыш) период в индивидуальном развитии организмов связан
- 11. В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, гаструлы, образования зародышевых листков, гистогенез и органогенез.
- 12. Стадию морулы проходят все многоклеточные животные, размножающиеся половым путем. В зависимости от видовой принадлежности деление идет
- 13. Гаструляция. Гаструляция (от греч. gastre — полость сосуда) — это следующий за образованием бластулы процесс движения
- 14. Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако для хордовых в период гаструляции характерно
- 16. Скачать презентацию
Слайд 2 Онтогенез, его типы и периодизация
Онтогенез (от греч. ontos — существо, genesis —
Онтогенез, его типы и периодизация Онтогенез (от греч. ontos — существо, genesis —

Слайд 3Тех, кто отдавал предпочтение яйцеклеткам, называли овистами, а тех, кто большое значение
Тех, кто отдавал предпочтение яйцеклеткам, называли овистами, а тех, кто большое значение

Слайд 4 В свою очередь, для объяснения «механизма» образования зародыша в половой клетке
В свою очередь, для объяснения «механизма» образования зародыша в половой клетке

Слайд 5Ланкестер (L. R. Lankester), Уитмен (Ch. Whitman), Раб ль (K. Rabl) и
Ланкестер (L. R. Lankester), Уитмен (Ch. Whitman), Раб ль (K. Rabl) и

Слайд 6 Эпигенез
ЭПИГЕНЕЗ (греческий epi- на, после, вслед за + genesis зарождение, происхождение) —
Эпигенез ЭПИГЕНЕЗ (греческий epi- на, после, вслед за + genesis зарождение, происхождение) —

Слайд 7Попытку разработать механическую теорию эпигенеза предпринял Р. Декарт, полагавший, что движущей силой
Попытку разработать механическую теорию эпигенеза предпринял Р. Декарт, полагавший, что движущей силой

Слайд 8Согласно современным представлениям, основные закономерности развития определяются генетической информацией, содержащейся в ДНК
Согласно современным представлениям, основные закономерности развития определяются генетической информацией, содержащейся в ДНК

Слайд 9Понятия о росте организмов (размножении клеток), дифферен-цировке клеток и о морфогенезе позволяют
Понятия о росте организмов (размножении клеток), дифферен-цировке клеток и о морфогенезе позволяют

Слайд 10Периоды
Проэмбриональный (от греч. pro — до, embryon — зародыш) период в индивидуальном
Периоды Проэмбриональный (от греч. pro — до, embryon — зародыш) период в индивидуальном

Слайд 11В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, гаструлы, образования зародышевых
В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, гаструлы, образования зародышевых

Слайд 12Стадию морулы проходят все многоклеточные животные, размножающиеся половым путем. В зависимости от
Стадию морулы проходят все многоклеточные животные, размножающиеся половым путем. В зависимости от

Слайд 13Гаструляция.
Гаструляция (от греч. gastre — полость сосуда) — это следующий за
Гаструляция. Гаструляция (от греч. gastre — полость сосуда) — это следующий за

Слайд 14Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако для хордовых
Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако для хордовых

 О пользе ночного сна
О пользе ночного сна Крахмал и его свойства
Крахмал и его свойства Изменения, происходящие с витаминами в процессах технологической переработки сырья
Изменения, происходящие с витаминами в процессах технологической переработки сырья Отряд гусеобразные
Отряд гусеобразные Пищеварительная система
Пищеварительная система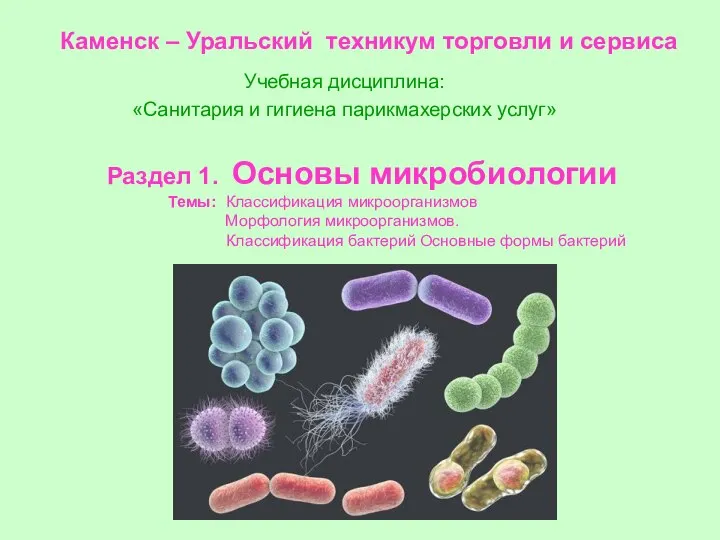 Классификация микроорганизмов. Морфология микроорганизмов. Классификация бактерий
Классификация микроорганизмов. Морфология микроорганизмов. Классификация бактерий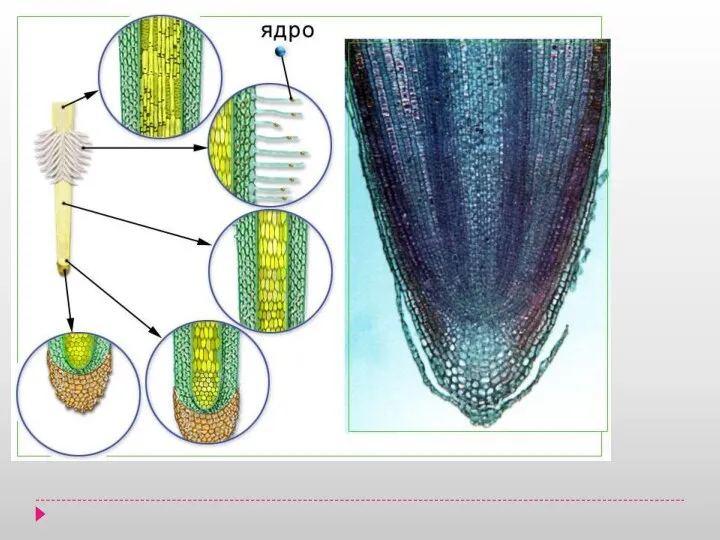 Анатомия корня
Анатомия корня Транскрипція та реплікація
Транскрипція та реплікація Євгеніка: практика, спрямована на створення кращої породи людей
Євгеніка: практика, спрямована на створення кращої породи людей Растения - псаммофиты. Экологические условия произрастания растений на песках
Растения - псаммофиты. Экологические условия произрастания растений на песках Презентация на тему Общая характеристика класса Птиц
Презентация на тему Общая характеристика класса Птиц  Биологический алфавит
Биологический алфавит Семейство кошачьих
Семейство кошачьих Методы исследования в биологии. Урок № 2, 5 класс
Методы исследования в биологии. Урок № 2, 5 класс Пищеварение в ротовой полости
Пищеварение в ротовой полости Гибридологический метод Г. Менделя
Гибридологический метод Г. Менделя Полевые цветы
Полевые цветы Яблоко
Яблоко Геномные и хромосомные мутации в организме человека
Геномные и хромосомные мутации в организме человека История развития эволюционных идей
История развития эволюционных идей Анатомия органа зрения
Анатомия органа зрения Внешнее строение рыб
Внешнее строение рыб Подземные побеги
Подземные побеги Грегор Иоганн Мендель (1822 - 1884). Доминантные и рецессивные признаки
Грегор Иоганн Мендель (1822 - 1884). Доминантные и рецессивные признаки Тип Членистоногие, класс Ракообразные
Тип Членистоногие, класс Ракообразные Гомологичные органы
Гомологичные органы Приспособления животных
Приспособления животных Пищеварение в желудке. Пищеварительные железы – печень, поджелудочная железа (Урок 36)
Пищеварение в желудке. Пищеварительные железы – печень, поджелудочная железа (Урок 36)