- Электрофизиология

Содержание
- 2. Результат возбуждения: Нервная → передача нервного импульса (электрический, химический) Мышечная → сокращение мышцы, совершение работы Секреторная
- 3. ЭЛЕКТРОФИЗИОЛОГИЯ в 1902 г. Ю. Бернштейном была развита мембранно-ионная теория, которая модифицирована и экспериментально обоснована А.
- 4. Транспорт: активный и пассивный Ионные каналы – интегральные белки, которые обеспечивают пассивный транспорт ионов по градиенту
- 5. ЭЛЕКТРОФИЗИОЛОГИЯ Мембранный потенциал Потенциал покоя Разность потенциалов между наружной и внутренней поверхностью мембраны покоящейся клетки. ПП
- 6. ЭЛЕКТРОФИЗИОЛОГИЯ В развитии возбуждения выделяют 4 этапа: 1) предшествующее возбуждению состояние покоя (статическая поляризация); 2) деполяризацию;
- 7. ЭЛЕКТРОФИЗИОЛОГИЯ Статическая поляризация – наличие постоянной разности потенциалов между наружной и внутренней поверхностями клеточной мембраны. Деполяризация
- 8. ЭЛЕКТРОФИЗИОЛОГИЯ Для возникновения возбуждения сила раздражающего тока должна нарастать достаточно круто. При медленном нарастании силы тока
- 9. ЭЛЕКТРОФИЗИОЛОГИЯ Свойства возбудимых тканей: 1. лабильность – способность возбудимой ткани реагировать на раздражение с определенной скоростью.
- 10. ЭЛЕКТРОФИЗИОЛОГИЯ Местный/локальный (генераторный, постсинаптический): нет латентного (скрытого) периода - возникает сразу же при действии раздражителя; нет
- 11. ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ Bся нервная система представляет собой совокупность нейронов, которые контактируют друг с другом при
- 12. ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ Синапсы можно классифицировать: по местоположению и принадлежности соответствующим клеткам — нервно—мышечные, нейро—нейрональные,
- 13. Механизм передачи в химическом синапсе: Поступление нервного импульса в синаптическую бляшку, Деполяризация ПреМ и повышение ее
- 14. Большинство нейромедиаторов — аминокислоты и их производные. Одни нейроны модифицируют аминокислоты с образованием аминов (норадреналин, серотонин,
- 15. Основные типы строения нервной системы А— диффузный (кишечнополостные); Б— цепочечный/узловой (членистоногие); В — трубчатый (цнс позвоночных).
- 16. ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ Глия — тип клеток, отличный от нервных и распространенный по всей нервной системе.
- 17. ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ Функционально в нейроне выделяют: 1) воспринимающую часть (дендриты и мембрану сомы нейрона);
- 18. ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ Функции нейронов: 1) генерализация нервного импульса; 2) получение, хранение и передача информации; 3)
- 19. ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ В нервной клетке 2 процесса: Возбуждение и Торможение Внешнее торможение условного рефлекса наступает
- 20. ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ Суммация (пространственная - возникновение ответной реакции при одновременном действии нескольких подпороговых раздражителей.
- 21. ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ нервные сети - система нейронных цепочек/комплексов на разных уровнях, предающие торможение/возбуждение Общий
- 22. ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ реципрокное (А)-антагонисты возвратное (Б)-собственный импульс к т.н. паралелльное (В)-в.блокирует само себя, посылая
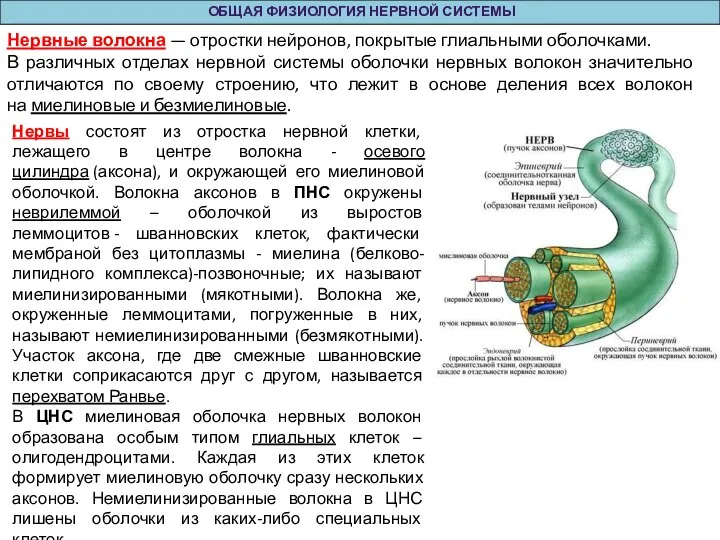
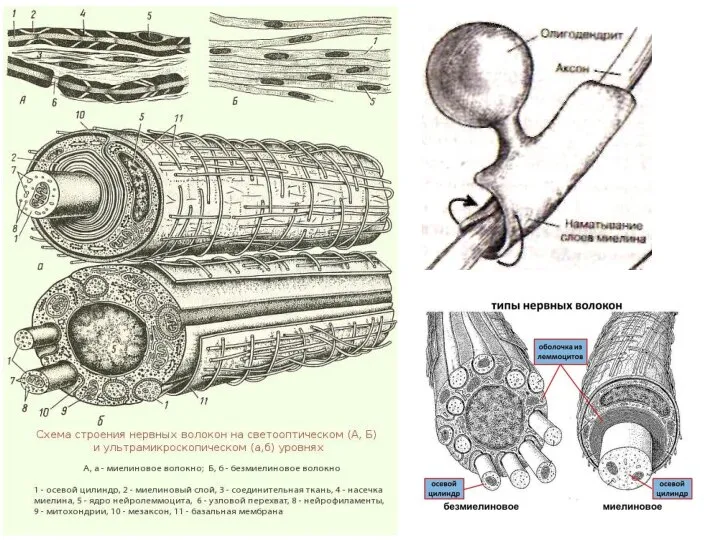
- 24. Нервы состоят из отростка нервной клетки, лежащего в центре волокна - осевого цилиндра (аксона), и окружающей
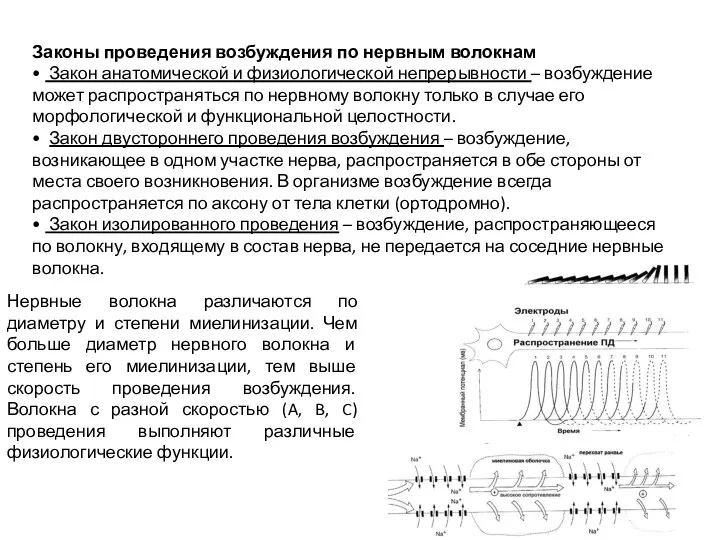
- 27. Законы проведения возбуждения по нервным волокнам • Закон анатомической и физиологической непрерывности – возбуждение может распространяться
- 28. Типы мышечной ткани ФИЗИОЛОГИЯ ОДА. СКЕЛЕТНЫЕ МЫШЦЫ поперечно-полосатая
- 29. Поперечно-полосатая мышечная ткань
- 30. «Клетка» скелетной мышцы - мышечное волокно=симпласт - содержат сократительные нити (миофибриллы), состоящие из двух разных белков
- 32. Мышечные волокна млекопитающих делятся на: экстрафузальные - выполняют всю работу сокращения мышцы интрафузальные - для детектирования
- 34. Строение саркомера
- 35. Гладкая мышечная ткань
- 36. Сердечная мышечная ткань
- 37. ФИЗИОЛОГИЯ ОДА Деполяризация Быстрая реполяризация Плато Медленная реполяризация ПД мышечной ткани
- 38. 1 двигательная концевая пластинка
- 39. ФИЗИОЛОГИЯ ОДА Сопряжение возбуждения с сокращением и расслаблением представляет комплекс процессов, связывающих электрическую деполяризацию на сарколемме
- 40. ФИЗИОЛОГИЯ ОДА Цикл: Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата
- 41. ФИЗИОЛОГИЯ ОДА Мышечное сокращение — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении
- 43. Скачать презентацию
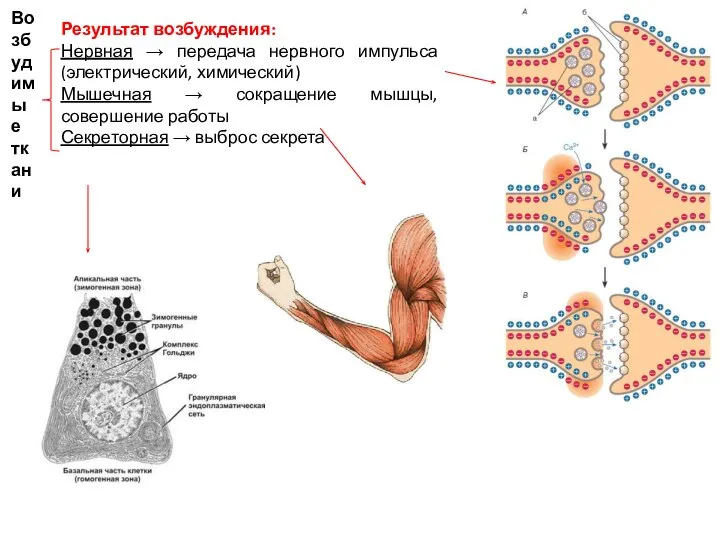
Слайд 2Результат возбуждения:
Нервная → передача нервного импульса (электрический, химический)
Мышечная → сокращение мышцы, совершение
Результат возбуждения:
Нервная → передача нервного импульса (электрический, химический)
Мышечная → сокращение мышцы, совершение
Слайд 3ЭЛЕКТРОФИЗИОЛОГИЯ
в 1902 г. Ю. Бернштейном была развита мембранно-ионная теория, которая модифицирована и
ЭЛЕКТРОФИЗИОЛОГИЯ
в 1902 г. Ю. Бернштейном была развита мембранно-ионная теория, которая модифицирована и

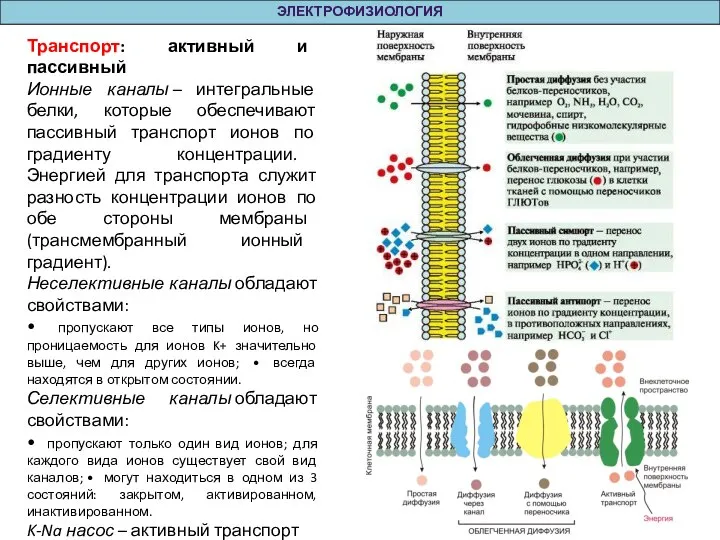
Слайд 4Транспорт: активный и пассивный
Ионные каналы – интегральные белки, которые обеспечивают пассивный транспорт ионов
Транспорт: активный и пассивный
Ионные каналы – интегральные белки, которые обеспечивают пассивный транспорт ионов
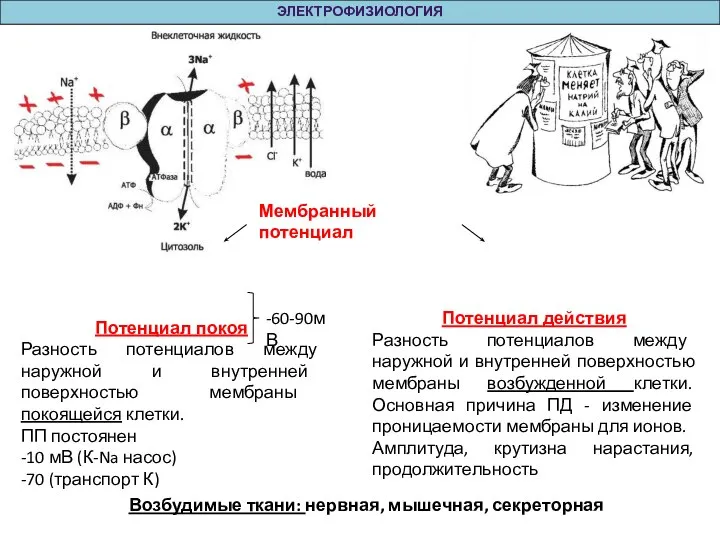
Слайд 5ЭЛЕКТРОФИЗИОЛОГИЯ
Мембранный потенциал
Потенциал покоя
Разность потенциалов между наружной и внутренней поверхностью мембраны покоящейся клетки.
ПП
ЭЛЕКТРОФИЗИОЛОГИЯ
Мембранный потенциал
Потенциал покоя
Разность потенциалов между наружной и внутренней поверхностью мембраны покоящейся клетки.
ПП
Слайд 6ЭЛЕКТРОФИЗИОЛОГИЯ
В развитии возбуждения выделяют 4 этапа:
1) предшествующее возбуждению состояние покоя (статическая поляризация);
2)
ЭЛЕКТРОФИЗИОЛОГИЯ
В развитии возбуждения выделяют 4 этапа:
1) предшествующее возбуждению состояние покоя (статическая поляризация);
2)
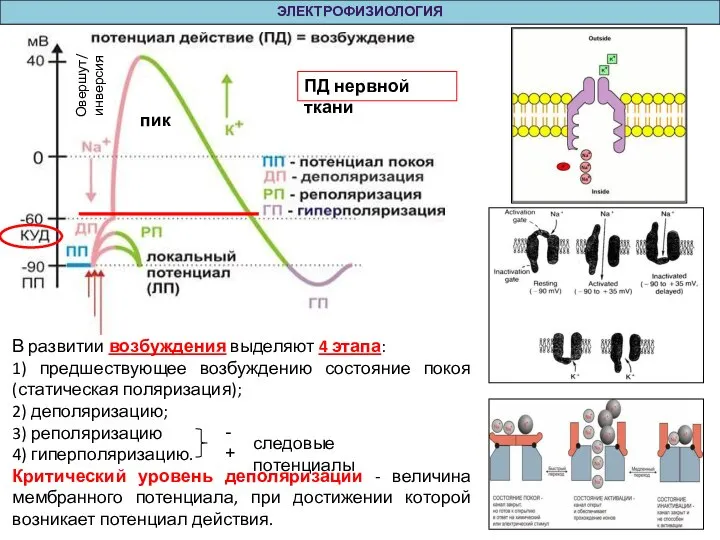
Слайд 7ЭЛЕКТРОФИЗИОЛОГИЯ
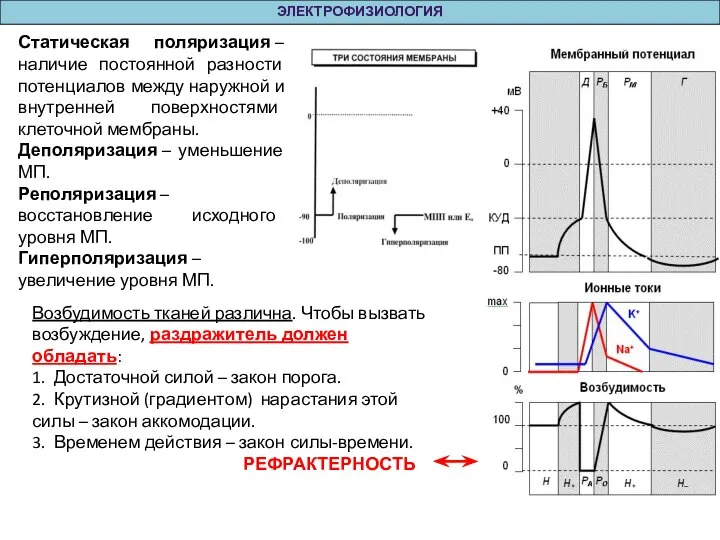
Статическая поляризация – наличие постоянной разности потенциалов между наружной и внутренней поверхностями клеточной
ЭЛЕКТРОФИЗИОЛОГИЯ
Статическая поляризация – наличие постоянной разности потенциалов между наружной и внутренней поверхностями клеточной
Слайд 8ЭЛЕКТРОФИЗИОЛОГИЯ
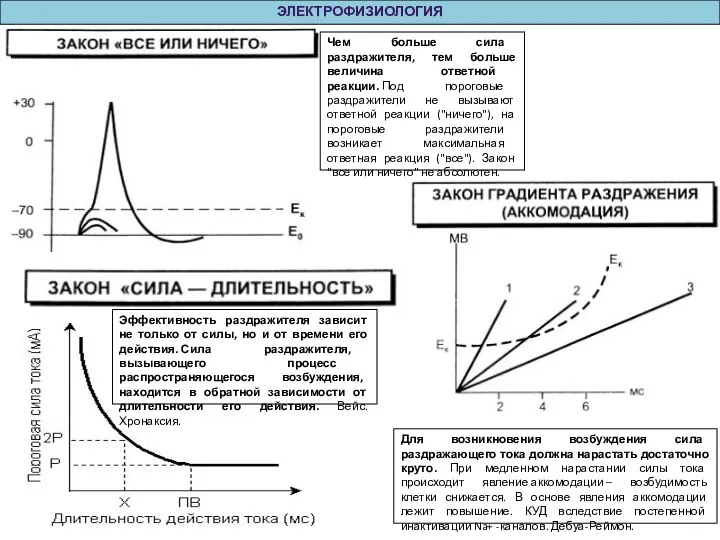
Для возникновения возбуждения сила раздражающего тока должна нарастать достаточно круто. При медленном
ЭЛЕКТРОФИЗИОЛОГИЯ
Для возникновения возбуждения сила раздражающего тока должна нарастать достаточно круто. При медленном
Слайд 9ЭЛЕКТРОФИЗИОЛОГИЯ
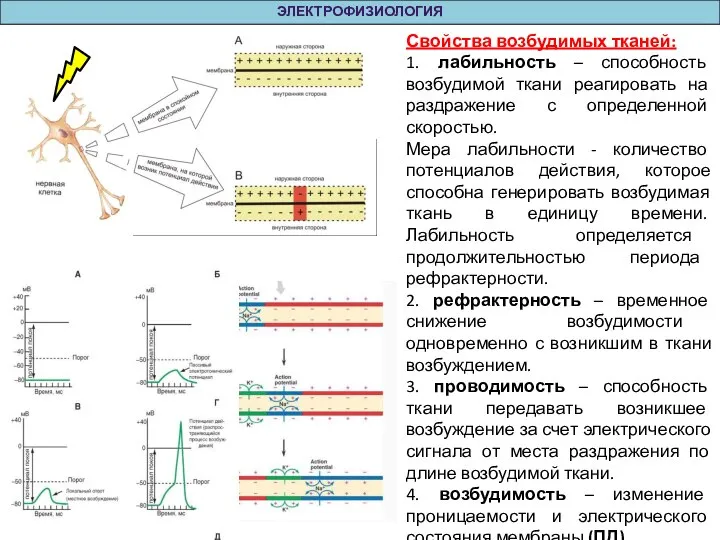
Свойства возбудимых тканей:
1. лабильность – способность возбудимой ткани реагировать на раздражение с
ЭЛЕКТРОФИЗИОЛОГИЯ
Свойства возбудимых тканей:
1. лабильность – способность возбудимой ткани реагировать на раздражение с
Слайд 10ЭЛЕКТРОФИЗИОЛОГИЯ
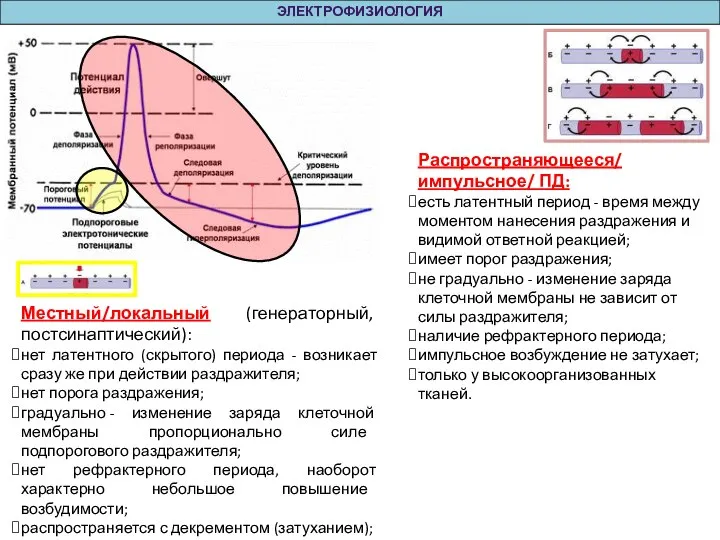
Местный/локальный (генераторный, постсинаптический):
нет латентного (скрытого) периода - возникает сразу же при действии
ЭЛЕКТРОФИЗИОЛОГИЯ
Местный/локальный (генераторный, постсинаптический):
нет латентного (скрытого) периода - возникает сразу же при действии
Слайд 11ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Bся нервная система представляет собой совокупность нейронов, которые контактируют друг
ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Bся нервная система представляет собой совокупность нейронов, которые контактируют друг
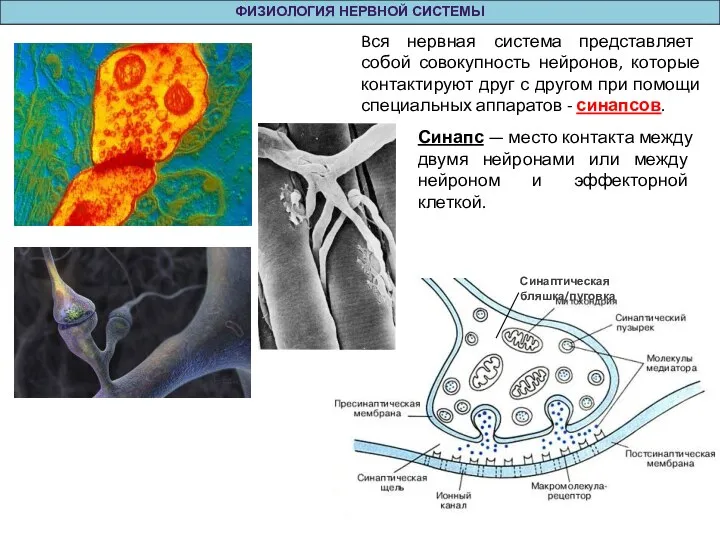
Слайд 12 ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Синапсы можно классифицировать:
по местоположению и принадлежности соответствующим клеткам — нервно—мышечные, нейро—нейрональные,
ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Синапсы можно классифицировать:
по местоположению и принадлежности соответствующим клеткам — нервно—мышечные, нейро—нейрональные,

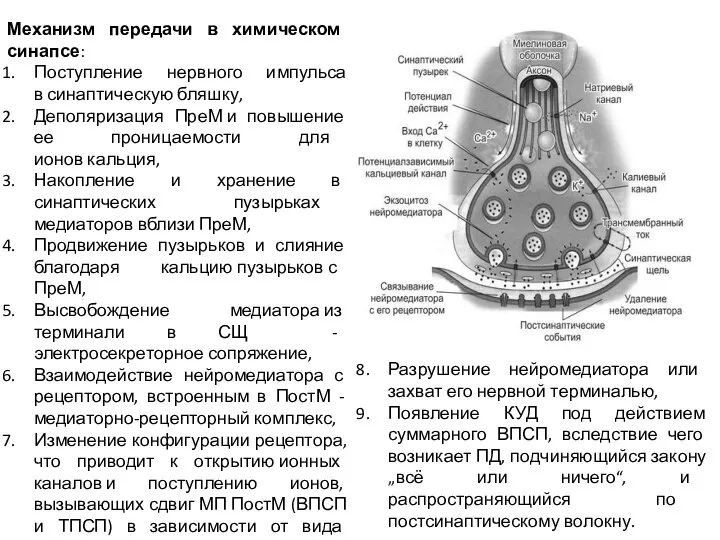
Слайд 13Механизм передачи в химическом синапсе:
Поступление нервного импульса в синаптическую бляшку,
Деполяризация ПреМ и повышение
Механизм передачи в химическом синапсе:
Поступление нервного импульса в синаптическую бляшку,
Деполяризация ПреМ и повышение
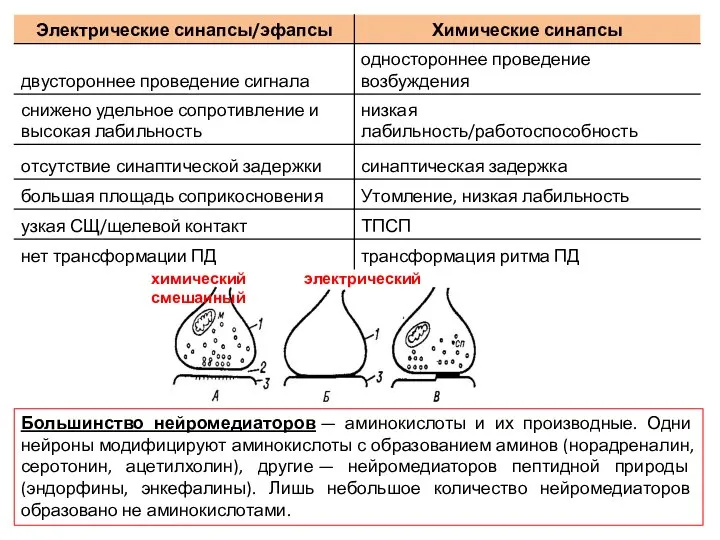
Слайд 14Большинство нейромедиаторов — аминокислоты и их производные. Одни нейроны модифицируют аминокислоты с образованием
Большинство нейромедиаторов — аминокислоты и их производные. Одни нейроны модифицируют аминокислоты с образованием
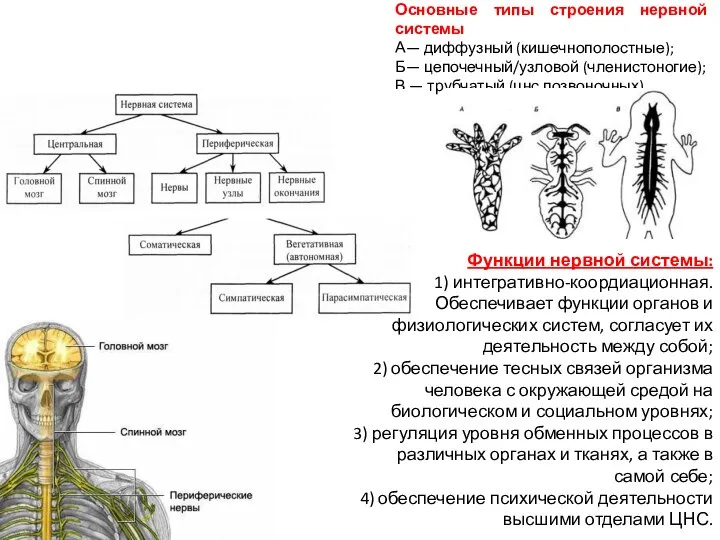
Слайд 15Основные типы строения нервной системы
А— диффузный (кишечнополостные);
Б— цепочечный/узловой (членистоногие);
В — трубчатый (цнс позвоночных).
Функции
Основные типы строения нервной системы
А— диффузный (кишечнополостные);
Б— цепочечный/узловой (членистоногие);
В — трубчатый (цнс позвоночных).
Функции
Слайд 16ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
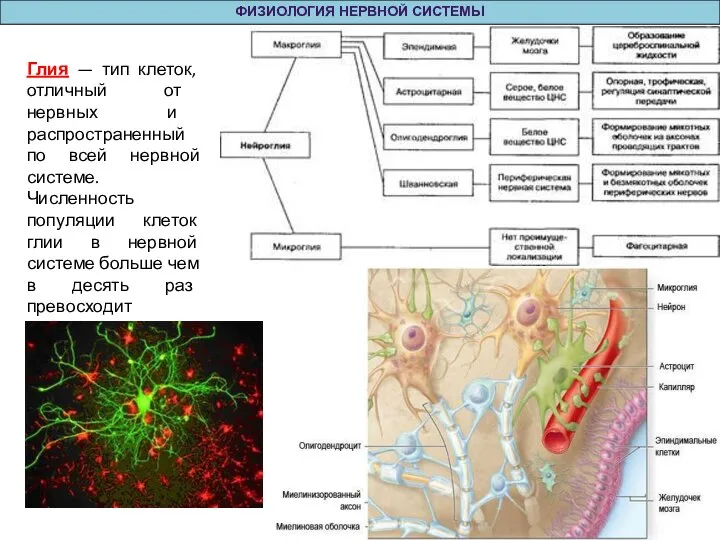
Глия — тип клеток, отличный от нервных и распространенный по
ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Глия — тип клеток, отличный от нервных и распространенный по
Слайд 17 ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
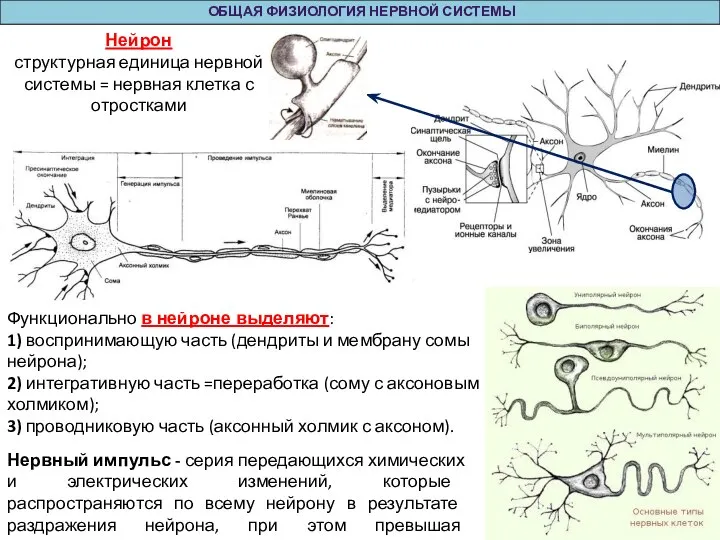
Функционально в нейроне выделяют:
1) воспринимающую часть (дендриты и мембрану
ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Функционально в нейроне выделяют:
1) воспринимающую часть (дендриты и мембрану
Слайд 18ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Функции нейронов:
1) генерализация нервного импульса;
2) получение, хранение и передача информации;
3) способность суммировать возбуждающие
ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Функции нейронов:
1) генерализация нервного импульса;
2) получение, хранение и передача информации;
3) способность суммировать возбуждающие

Слайд 19ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
В нервной клетке 2 процесса: Возбуждение и Торможение
Внешнее торможение условного рефлекса наступает
ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
В нервной клетке 2 процесса: Возбуждение и Торможение
Внешнее торможение условного рефлекса наступает

Слайд 20 ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
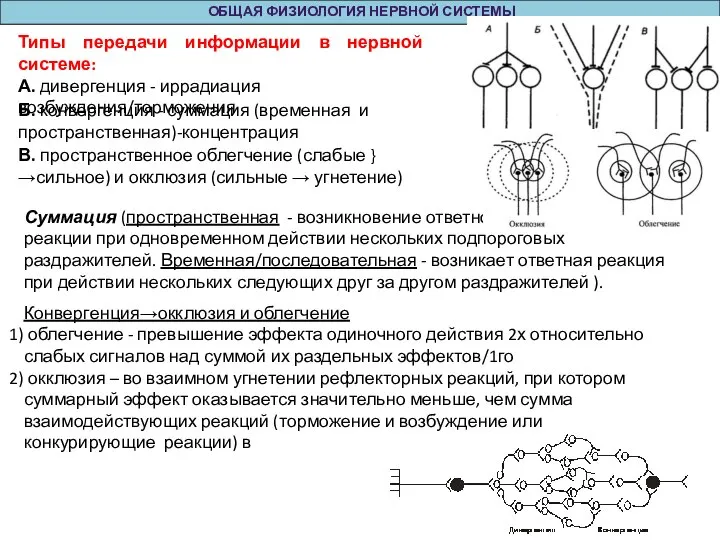
Суммация (пространственная - возникновение ответной
реакции при одновременном
ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
Суммация (пространственная - возникновение ответной
реакции при одновременном
Слайд 21 ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
нервные сети - система нейронных цепочек/комплексов на разных
ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
нервные сети - система нейронных цепочек/комплексов на разных

Слайд 22 ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
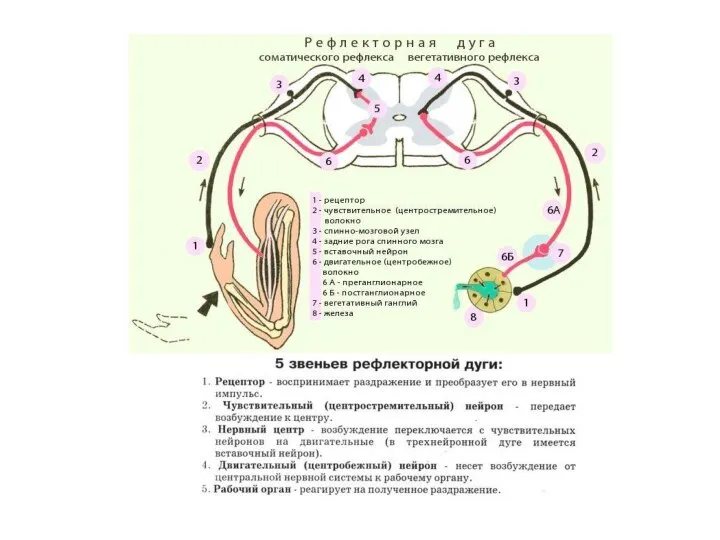
реципрокное (А)-антагонисты
возвратное (Б)-собственный импульс к т.н.
паралелльное (В)-в.блокирует само
ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
реципрокное (А)-антагонисты
возвратное (Б)-собственный импульс к т.н.
паралелльное (В)-в.блокирует само

Слайд 24Нервы состоят из отростка нервной клетки, лежащего в центре волокна - осевого
Нервы состоят из отростка нервной клетки, лежащего в центре волокна - осевого
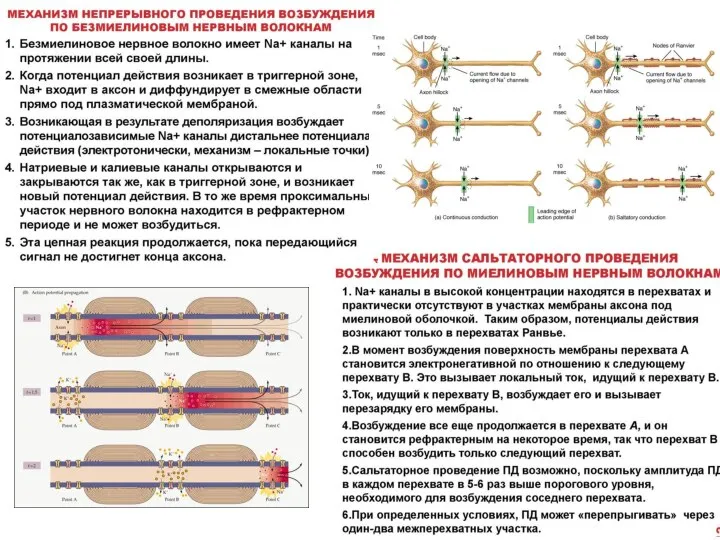
Слайд 27Законы проведения возбуждения по нервным волокнам
• Закон анатомической и физиологической непрерывности – возбуждение может распространяться
Законы проведения возбуждения по нервным волокнам
• Закон анатомической и физиологической непрерывности – возбуждение может распространяться
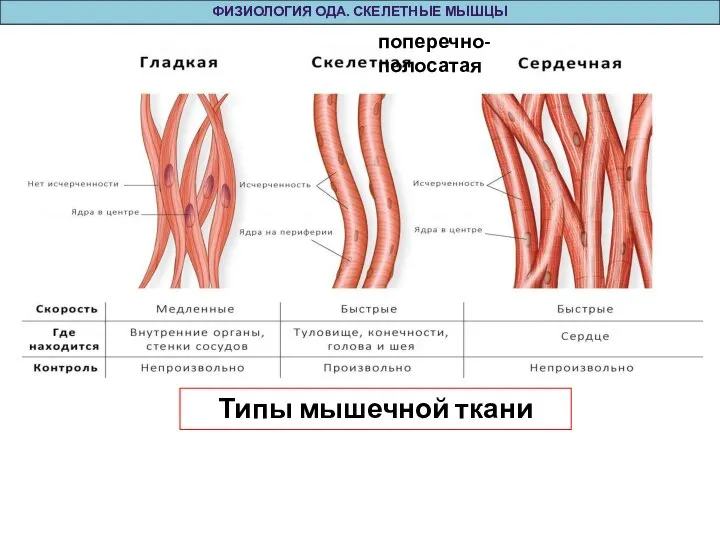
Слайд 28Типы мышечной ткани
ФИЗИОЛОГИЯ ОДА. СКЕЛЕТНЫЕ МЫШЦЫ
поперечно-полосатая
Типы мышечной ткани
ФИЗИОЛОГИЯ ОДА. СКЕЛЕТНЫЕ МЫШЦЫ
поперечно-полосатая
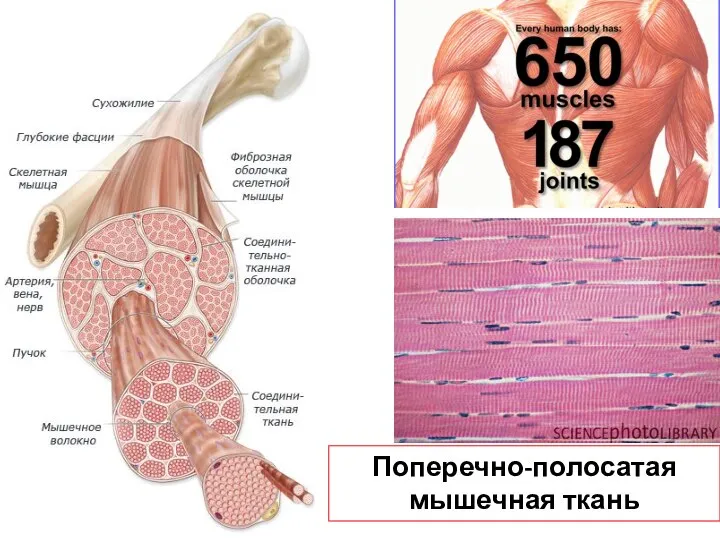
Слайд 29Поперечно-полосатая мышечная ткань
Поперечно-полосатая мышечная ткань
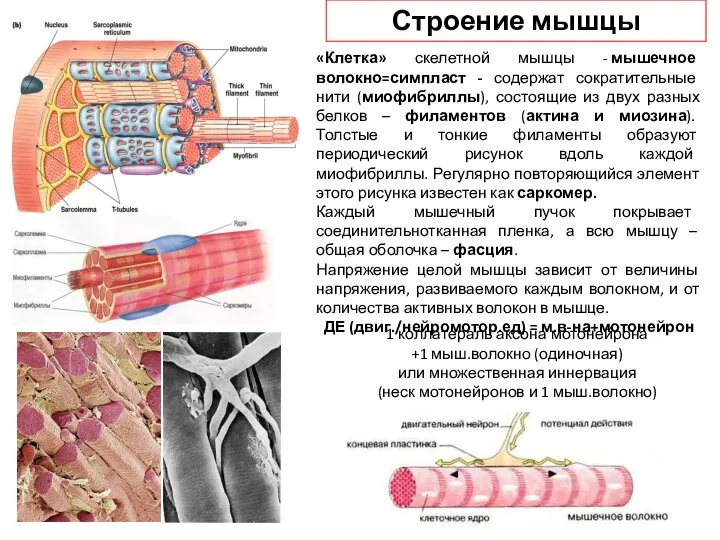
Слайд 30«Клетка» скелетной мышцы - мышечное волокно=симпласт - содержат сократительные нити (миофибриллы), состоящие из
«Клетка» скелетной мышцы - мышечное волокно=симпласт - содержат сократительные нити (миофибриллы), состоящие из
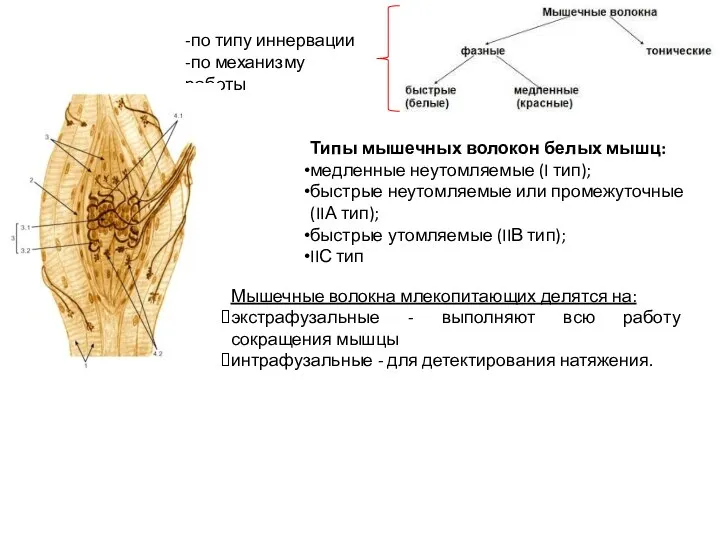
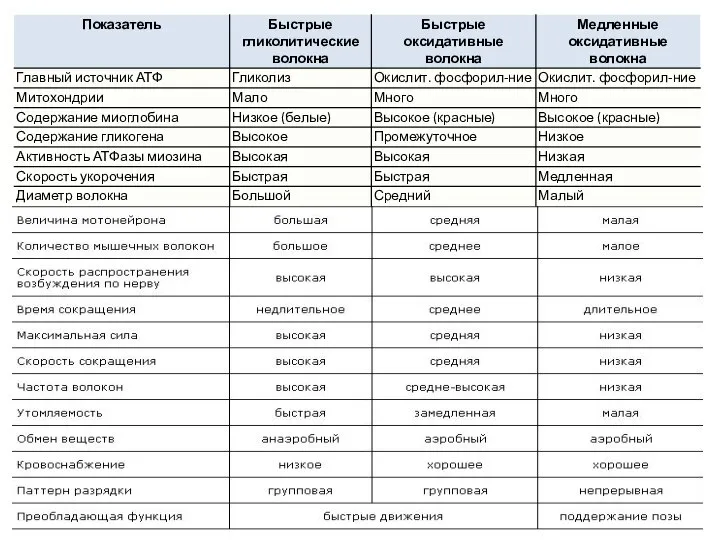
Слайд 32Мышечные волокна млекопитающих делятся на:
экстрафузальные - выполняют всю работу сокращения мышцы
интрафузальные -
Мышечные волокна млекопитающих делятся на:
экстрафузальные - выполняют всю работу сокращения мышцы
интрафузальные -
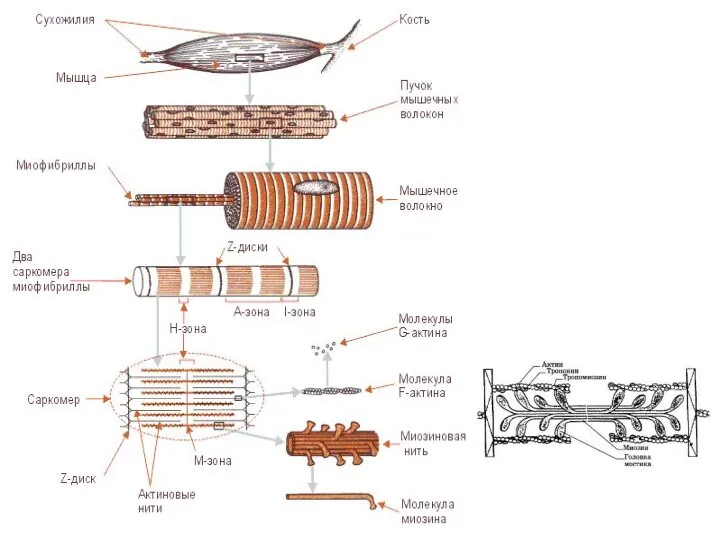
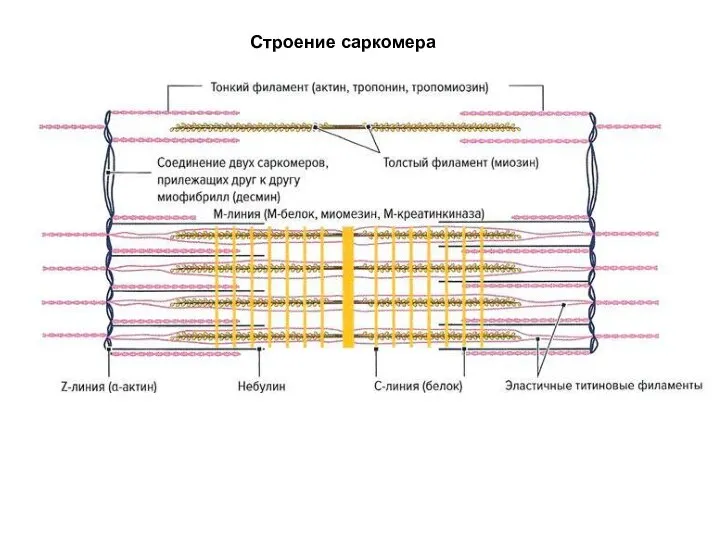
Слайд 34Строение саркомера
Строение саркомера
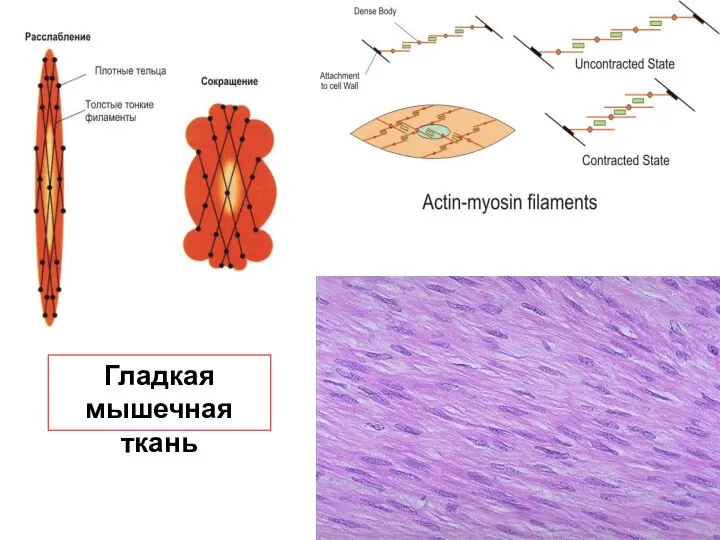
Слайд 35Гладкая
мышечная ткань
Гладкая
мышечная ткань
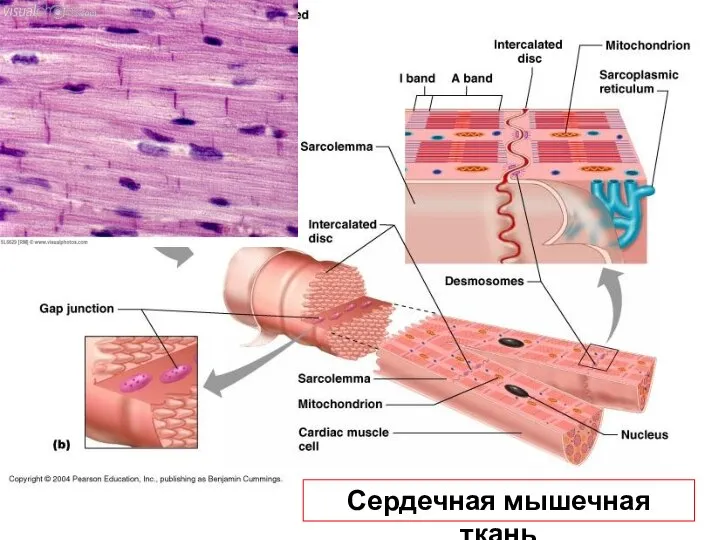
Слайд 36Сердечная мышечная ткань
Сердечная мышечная ткань
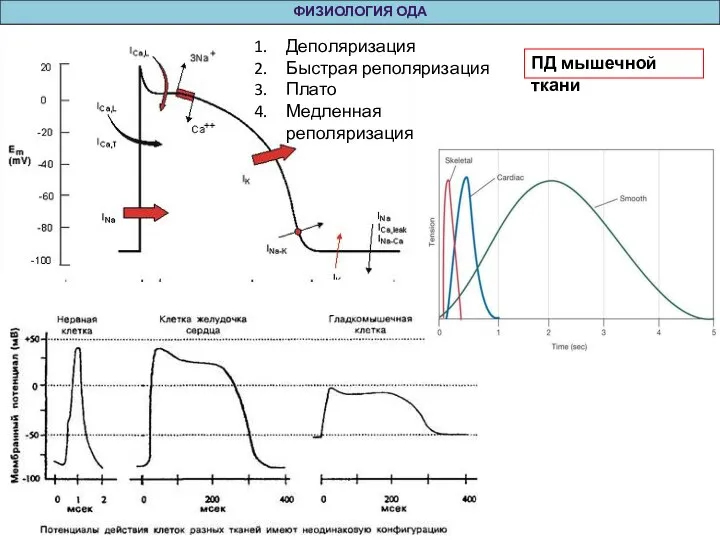
Слайд 37ФИЗИОЛОГИЯ ОДА
Деполяризация
Быстрая реполяризация
Плато
Медленная реполяризация
ПД мышечной ткани
ФИЗИОЛОГИЯ ОДА
Деполяризация
Быстрая реполяризация
Плато
Медленная реполяризация
ПД мышечной ткани
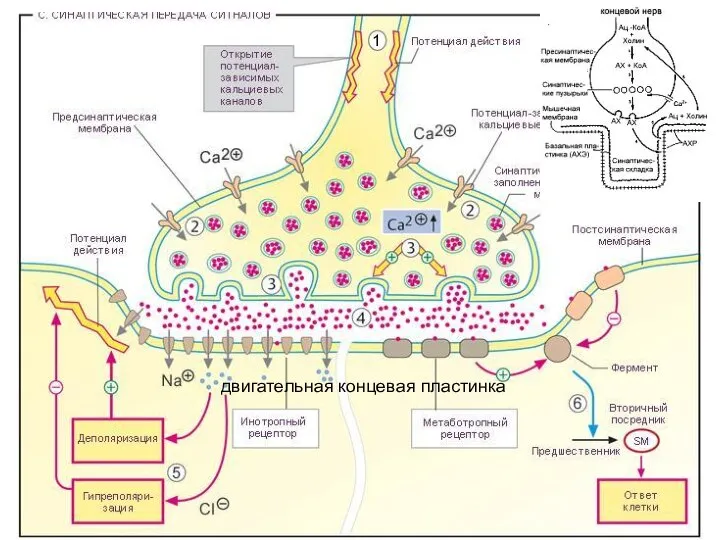
Слайд 381
двигательная концевая пластинка
1
двигательная концевая пластинка
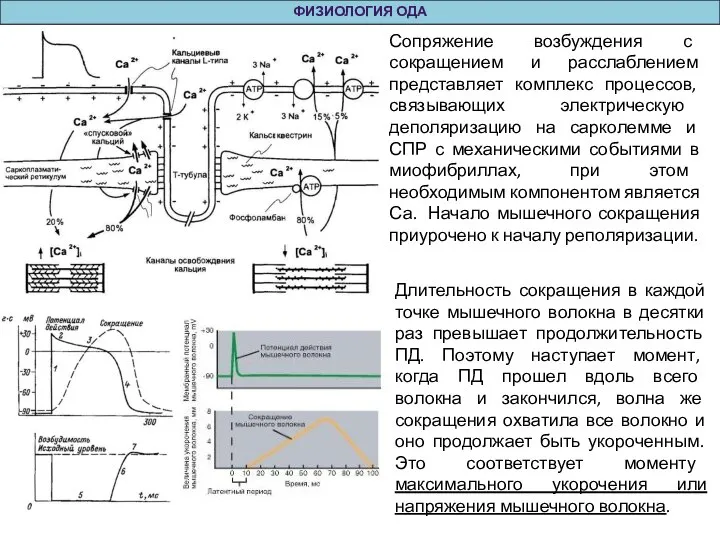
Слайд 39ФИЗИОЛОГИЯ ОДА
Сопряжение возбуждения с сокращением и расслаблением представляет комплекс процессов, связывающих электрическую
ФИЗИОЛОГИЯ ОДА
Сопряжение возбуждения с сокращением и расслаблением представляет комплекс процессов, связывающих электрическую
Слайд 40ФИЗИОЛОГИЯ ОДА
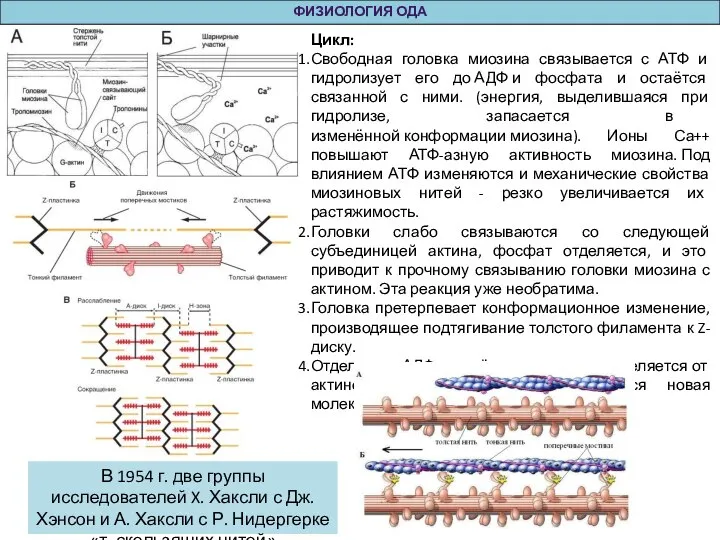
Цикл:
Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата
ФИЗИОЛОГИЯ ОДА
Цикл:
Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата
Слайд 41ФИЗИОЛОГИЯ ОДА
Мышечное сокращение — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки.
ФИЗИОЛОГИЯ ОДА
Мышечное сокращение — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки.

 Снег и лед в жизни растений, животных, людей
Снег и лед в жизни растений, животных, людей Мақалдар сәйкестігі. Табиғаты
Мақалдар сәйкестігі. Табиғаты Почемучка
Почемучка Питание и пищеварение
Питание и пищеварение Цепи питания. Поток энергии
Цепи питания. Поток энергии Хромосомная теория наследственности
Хромосомная теория наследственности Приспособляемость живых организмов к суровым условиям Антарктиды
Приспособляемость живых организмов к суровым условиям Антарктиды Нервная система человека
Нервная система человека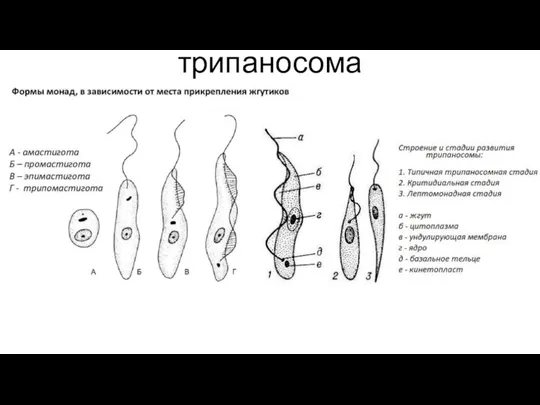 Трипаносома. Лямблия
Трипаносома. Лямблия Колорад қоңызы
Колорад қоңызы Понятие инноваций и их роль в развитии предприятия
Понятие инноваций и их роль в развитии предприятия Фотосинтез
Фотосинтез Успехи селекции микроорганизмов. Биотехнология и генная инженерия
Успехи селекции микроорганизмов. Биотехнология и генная инженерия Цветок. Строение и значение цветка. 6 класс. Часть 2
Цветок. Строение и значение цветка. 6 класс. Часть 2 Пищеварительная система
Пищеварительная система Растения. Распространение растений
Растения. Распространение растений Почему эволюция - это факт
Почему эволюция - это факт Применения информационных технологий в изучении влияния фитогормонов на рост орхидей
Применения информационных технологий в изучении влияния фитогормонов на рост орхидей Такие большие, маленькие насекомые
Такие большие, маленькие насекомые Генотип и фенотип
Генотип и фенотип Класс Млекопитающие. Отряд Приматы
Класс Млекопитающие. Отряд Приматы Осенние цветы
Осенние цветы Египетская гладкошерстная кошка
Египетская гладкошерстная кошка Презентация на тему ОРГАНЫ И СИСТЕМЫ ОРГАНОВ
Презентация на тему ОРГАНЫ И СИСТЕМЫ ОРГАНОВ  Обмен веществ
Обмен веществ Комнатные растения (часть 1)
Комнатные растения (часть 1) Тонкий кишечник
Тонкий кишечник City farming - the technology of the 3rd millenium
City farming - the technology of the 3rd millenium