- Биомеханика. Термодинамика механохимических процессов

Содержание
- 2. Термодинамика механохимических процессов Живые организмы совершают механическую работу в изотермических (Т-const) и изобарических (P-const) условиях. Следовательно,
- 3. Термодинамика механохимических процессов При постоянной температуре тепловая энергия не может быть использована для совершения работы. Работа
- 4. Термодинамика механохимических процессов Механохимический процесс может выполняться циклически при переходе от одного химического потенциала к другому
- 5. Термодинамика механохимических процессов F l μ1 μ2 n1 n2 Механохимический цикл в координатах F и l
- 6. Структура мышцы и мышечных белков Отдельное мышечное волокно имеет диаметр 20-80 мкм. Оно окружено мембраной, имеющей
- 7. Структура миофибриллы Z-линия саркомер А Н I I На продольном срезе миофибриллы видны т.н. Z-полоски. Пространство
- 8. Структура миофибриллы Z-линия А Н I-полоса Электронно-микроскопические исследования показали, что толстые нити (полоса А) образованы белком
- 9. Структура актина Тропонин Тропонин Тропомиозин G актин Актиновая нить представляет из себя двойную спираль с субъединицами
- 10. Структура миозина ЛММ (150000) ТММ S2 (60000) ТММ S1 (120000) Миозин представляет из себя фибриллярный белок.
- 11. Структура миозина При образовании толстой нити молекулы миозона агрегируют, предположительно, в результате взаимодействий между “хвостами.” Толстые
- 12. Z-линия саркомер А Н I I Согласно данным световой и электронной микроскопии, при сокращении мышцы происходит
- 13. Z-линия А Н I-полоса Основные структурные особенности мышечного сокращения. Согласно данным световой и электронной микроскопии, при
- 14. Биохимия мышечного сокращения. Миозин + АТФ ↔Миозин-АТФ ↔Миозин ↔Миозин +АДФ+Ф АДФ Ф 1.Установлено, что миозин способен
- 15. Тропонин Тропонин Тропомиозин G актин Ca2+ регулирует сократительный процесс, воздействуя на тропонин. При отсутствии кальция тропонин
- 16. Механика мышечного сокращения. Основные закономерности. Различают изометрическое сокращение: при фиксированной длине. l-const и изотоническое: укорочение мышцы
- 17. Изотоническое сокращение. При изотоническом сокращении (с постоянным грузом) после стимуляции напряжение (F/S=E) начинает расти, достигая максимума
- 18. Уравнение Хилла. Экспериментально установленные закономерности Хилл сформулировал в следующем уравнении: (P+a)V=b(P0-P) Где Р – вес груза,
- 19. Уравнение Хилла. Уравнение Хилла справедливо в условиях укорочения, идущего с постоянной скоростью и при длине мышцы
- 20. Работа, производимая мышцей при изотоническом сокращении. А=РVt, где А-работа, Р-вес груза, V-скорость сокращения, t-время. Согласно уравнению
- 21. Z-линия А Н I-полоса Максимальное напряжение, зафиксированное в икроножной мышце лягушки, Е=30н/см2, что соответствует 3• 10-10
- 22. Термодинамика мышечного сокращения. ∆Q= ∆U+A. Согласно первому закону термодинамики, теплота поглощенная (или выделенная) системой (∆Q) равна
- 23. Термодинамика мышечного сокращения. Qx=a Δl t,c Q Qx Δl Δl Δl Изометрическое сокращение t,c Q Изометрическое
- 24. Теория мышечного сокращения. Кинетическая теория Дщеревского, основные положения: Три состояния поперечного мостика: разомкнутое, тянущее замкнутое (сила
- 25. Скорость укорочения мышечного волокна (т.к. укорачиваются к центру) V1=2Nu, где N-число саркомеров. Сила F, генерируемая в
- 27. Скачать презентацию
Слайд 2Термодинамика механохимических процессов
Живые организмы совершают механическую работу в изотермических (Т-const) и изобарических
Термодинамика механохимических процессов
Живые организмы совершают механическую работу в изотермических (Т-const) и изобарических

Слайд 3Термодинамика механохимических процессов
При постоянной температуре тепловая энергия не может быть использована для
Термодинамика механохимических процессов
При постоянной температуре тепловая энергия не может быть использована для

Слайд 4Термодинамика механохимических процессов
Механохимический процесс может выполняться циклически при переходе от одного химического
Термодинамика механохимических процессов
Механохимический процесс может выполняться циклически при переходе от одного химического

Слайд 5Термодинамика механохимических процессов
F
l
μ1
μ2
n1
n2
Механохимический цикл в координатах F и l
Термодинамика механохимических процессов
Термодинамика механохимических процессов
F
l
μ1
μ2
n1
n2
Механохимический цикл в координатах F и l
Термодинамика механохимических процессов

Слайд 6Структура мышцы и мышечных белков
Отдельное мышечное волокно имеет диаметр 20-80 мкм. Оно
Структура мышцы и мышечных белков
Отдельное мышечное волокно имеет диаметр 20-80 мкм. Оно

Слайд 7Структура миофибриллы
Z-линия
саркомер
А
Н
I
I
На продольном срезе миофибриллы видны т.н. Z-полоски. Пространство
между двумя соседними
Структура миофибриллы
Z-линия
саркомер
А
Н
I
I
На продольном срезе миофибриллы видны т.н. Z-полоски. Пространство
между двумя соседними

Слайд 8Структура миофибриллы
Z-линия
А
Н
I-полоса
Электронно-микроскопические исследования показали, что толстые нити (полоса А) образованы белком миозином,
Структура миофибриллы
Z-линия
А
Н
I-полоса
Электронно-микроскопические исследования показали, что толстые нити (полоса А) образованы белком миозином,

Слайд 9Структура актина
Тропонин
Тропонин
Тропомиозин
G актин
Актиновая нить представляет из себя двойную спираль
с субъединицами G-актина, повторяющимися
Структура актина
Тропонин
Тропонин
Тропомиозин
G актин
Актиновая нить представляет из себя двойную спираль
с субъединицами G-актина, повторяющимися

Слайд 10Структура миозина
ЛММ (150000)
ТММ
S2
(60000)
ТММ
S1
(120000)
Миозин представляет из себя фибриллярный белок. Он состоит
из молекул длиной
Структура миозина
ЛММ (150000)
ТММ
S2
(60000)
ТММ
S1
(120000)
Миозин представляет из себя фибриллярный белок. Он состоит
из молекул длиной

Слайд 11Структура миозина
При образовании толстой нити молекулы миозона агрегируют,
предположительно, в результате взаимодействий между
Структура миозина
При образовании толстой нити молекулы миозона агрегируют,
предположительно, в результате взаимодействий между

Слайд 12Z-линия
саркомер
А
Н
I
I
Согласно данным световой и электронной микроскопии, при сокращении
мышцы происходит сужение I-полос без
Z-линия
саркомер
А
Н
I
I
Согласно данным световой и электронной микроскопии, при сокращении
мышцы происходит сужение I-полос без

Слайд 13Z-линия
А
Н
I-полоса
Основные структурные особенности мышечного сокращения.
Согласно данным световой и электронной микроскопии, при сокращении
мышцы
Z-линия
А
Н
I-полоса
Основные структурные особенности мышечного сокращения.
Согласно данным световой и электронной микроскопии, при сокращении
мышцы

Слайд 14Биохимия мышечного сокращения.
Миозин + АТФ ↔Миозин-АТФ ↔Миозин ↔Миозин +АДФ+Ф
АДФ
Ф
1.Установлено, что миозин
Биохимия мышечного сокращения.
Миозин + АТФ ↔Миозин-АТФ ↔Миозин ↔Миозин +АДФ+Ф
АДФ
Ф
1.Установлено, что миозин

Слайд 15Тропонин
Тропонин
Тропомиозин
G актин
Ca2+ регулирует сократительный процесс, воздействуя на
тропонин. При отсутствии кальция тропонин
Тропонин
Тропонин
Тропомиозин
G актин
Ca2+ регулирует сократительный процесс, воздействуя на
тропонин. При отсутствии кальция тропонин

Слайд 16Механика мышечного сокращения. Основные закономерности.
Различают изометрическое сокращение: при фиксированной длине. l-const
и изотоническое:
Механика мышечного сокращения. Основные закономерности.
Различают изометрическое сокращение: при фиксированной длине. l-const
и изотоническое:

Слайд 17Изотоническое сокращение.
При изотоническом сокращении (с постоянным грузом) после стимуляции напряжение (F/S=E) начинает
Изотоническое сокращение.
При изотоническом сокращении (с постоянным грузом) после стимуляции напряжение (F/S=E) начинает

Слайд 18Уравнение Хилла.
Экспериментально установленные закономерности Хилл сформулировал в следующем
уравнении:
(P+a)V=b(P0-P)
Где Р – вес груза,
Уравнение Хилла.
Экспериментально установленные закономерности Хилл сформулировал в следующем
уравнении:
(P+a)V=b(P0-P)
Где Р – вес груза,

Слайд 19Уравнение Хилла.
Уравнение Хилла справедливо в условиях укорочения, идущего с постоянной скоростью и
Уравнение Хилла.
Уравнение Хилла справедливо в условиях укорочения, идущего с постоянной скоростью и

Слайд 20Работа, производимая мышцей при изотоническом сокращении.
А=РVt, где А-работа, Р-вес груза, V-скорость сокращения,
Работа, производимая мышцей при изотоническом сокращении.
А=РVt, где А-работа, Р-вес груза, V-скорость сокращения,

Слайд 21Z-линия
А
Н
I-полоса
Максимальное напряжение, зафиксированное в икроножной мышце лягушки, Е=30н/см2, что соответствует 3• 10-10
Z-линия
А
Н
I-полоса
Максимальное напряжение, зафиксированное в икроножной мышце лягушки, Е=30н/см2, что соответствует 3• 10-10

Слайд 22Термодинамика мышечного сокращения.
∆Q= ∆U+A. Согласно первому закону термодинамики, теплота поглощенная (или выделенная)
Термодинамика мышечного сокращения.
∆Q= ∆U+A. Согласно первому закону термодинамики, теплота поглощенная (или выделенная)

Слайд 23Термодинамика мышечного сокращения.
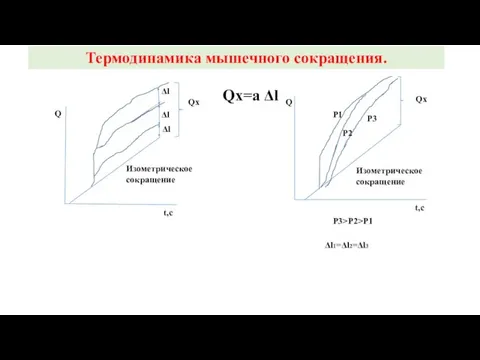
Qx=a Δl
t,c
Q
Qx
Δl
Δl
Δl
Изометрическое
сокращение
t,c
Q
Изометрическое
сокращение
Qx
Р1
Р2
Р3
Р3>P2>P1
Δl1=Δl2=Δl3
Термодинамика мышечного сокращения.
Qx=a Δl
t,c
Q
Qx
Δl
Δl
Δl
Изометрическое
сокращение
t,c
Q
Изометрическое
сокращение
Qx
Р1
Р2
Р3
Р3>P2>P1
Δl1=Δl2=Δl3
Слайд 24Теория мышечного сокращения.
Кинетическая теория Дщеревского, основные положения:
Три состояния поперечного мостика:
разомкнутое,
тянущее
Теория мышечного сокращения.
Кинетическая теория Дщеревского, основные положения:
Три состояния поперечного мостика:
разомкнутое,
тянущее

Слайд 25Скорость укорочения мышечного волокна (т.к. укорачиваются к центру)
V1=2Nu, где N-число саркомеров.
Сила F,
Скорость укорочения мышечного волокна (т.к. укорачиваются к центру)
V1=2Nu, где N-число саркомеров.
Сила F,

 Амфибии (или земноводные)
Амфибии (или земноводные) Зачем подкармливать птиц зимой?
Зачем подкармливать птиц зимой? Задача Ферменты
Задача Ферменты Пищеварение
Пищеварение Наследование количественных признаков.Тест
Наследование количественных признаков.Тест Coherently aligned nanoparticles within a biogenic single crystal: A biological prestressing strategy
Coherently aligned nanoparticles within a biogenic single crystal: A biological prestressing strategy Растительный и животный мир Крыма. Часть4
Растительный и животный мир Крыма. Часть4 Мхи
Мхи Антропогенез
Антропогенез Органы, осуществляющие выделение
Органы, осуществляющие выделение Развитие биологии в XIX в. западной Европе
Развитие биологии в XIX в. западной Европе Размножение живых организмов
Размножение живых организмов Эволюция в вопросах и ответах
Эволюция в вопросах и ответах Значение выделения для живых организмов. Продукты выделения у животных
Значение выделения для живых организмов. Продукты выделения у животных Пути и направления эволюции. 11 класс. 2
Пути и направления эволюции. 11 класс. 2 Буынаяқтылар. Ауыл кенесі, Алма кенесі
Буынаяқтылар. Ауыл кенесі, Алма кенесі Эволюция животного мира . Филогенез
Эволюция животного мира . Филогенез Учение о биосфере (2)
Учение о биосфере (2) Царство бактерий
Царство бактерий Маточное хозяйство
Маточное хозяйство Пути утилизации кислорода в организме
Пути утилизации кислорода в организме Анатомия слухового анализатора
Анатомия слухового анализатора Луговое разнотравье. Экологическая викторина
Луговое разнотравье. Экологическая викторина Животные живого уголка. Аквариум и его обитатели
Животные живого уголка. Аквариум и его обитатели Нервная система
Нервная система Презентация на тему ЖИЗНЕННЫЕ ФОРМЫ РАСТЕНИЙ
Презентация на тему ЖИЗНЕННЫЕ ФОРМЫ РАСТЕНИЙ  Экологические ниши бактерий
Экологические ниши бактерий Para qué son Estes partes de cuerpo humano
Para qué son Estes partes de cuerpo humano