- Fin_kletka

Содержание
- 2. КЛЕТОЧНАЯ ТЕОРИЯ Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития
- 3. Прокариоты Сине-зеленые водоросли и бактерии Бактерии: Кольцевидная ДНК Отсутствует ядерная оболочка Отсутствуют аппарат Гольджи и митохондрии
- 7. Эукариотическая клетка
- 9. Плазматическая мембрана Давсон, Даниэлли -1930г. Робертсон – 1935г. Гипотеза о строении элементарной мембраны: 1.Все мембраны имеют
- 10. Жидкостно – мозаическая структура плазматической мембраны Основная гипотеза была сформулирована в 1972 году Николсоном и Сингером
- 11. В основе лежит двойной слой липидов (фосфолипидов), имеющих полярные головки и длинные неполярные хвосты, представленные цепями
- 12. Белки мембран различаются по своему положению в мембране: Они могут глубоко проникать в липидный бислой или
- 13. . Расположение (локализация) белков в мембранах. Трансмембранные белки, например: 1 - гпикофорин А; 2 - рецептор
- 14. Поверхностные белки часто прикрепляются к мембране, взаимодействуя с интегральными Ряд пищеварительных ферментов, участвующих в гидролизе крахмала
- 15. Большая часть погруженных белков -ферменты Интегральные (погруженные ) расположены в определенном порядке, так чтобы продукт реакции
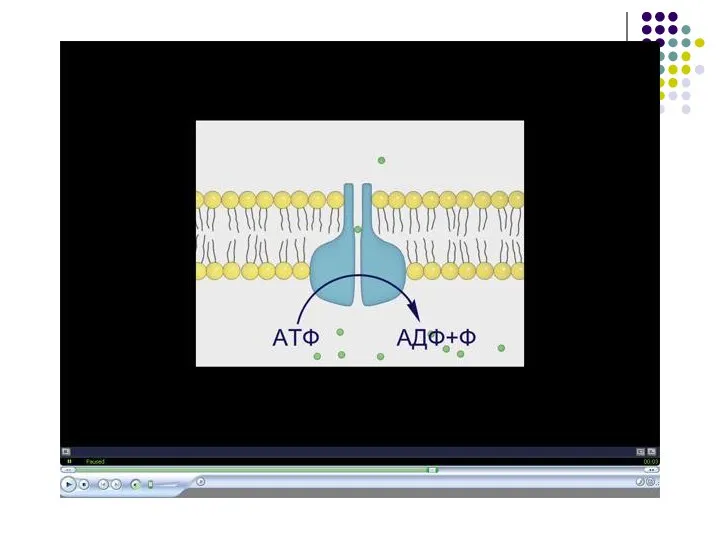
- 17. Функции мембраны обеспечиваются благодаря избирательной проницаемости Транспортная диффузия (от меньшей концентрации к большей) – жирорастворимые соединения,
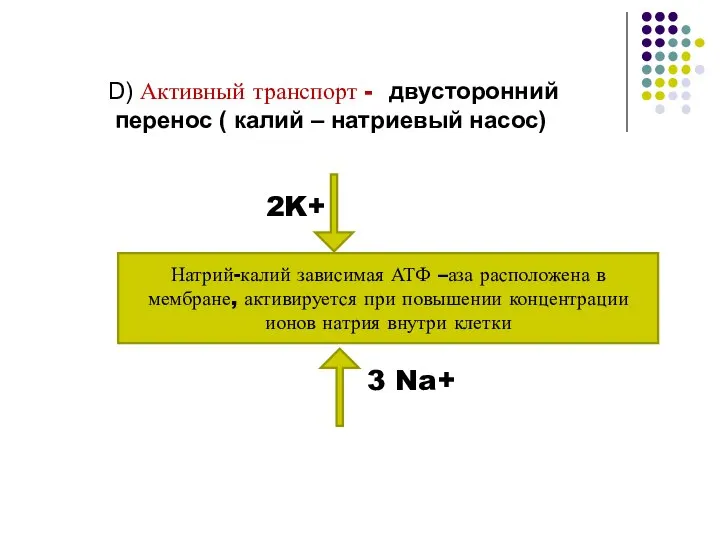
- 18. D) Активный транспорт - двусторонний перенос ( калий – натриевый насос) Натрий-калий зависимая АТФ –аза расположена
- 19. Эндоцитоз (фаго – и пиноцитоз) Экзоцитоз Рецепторная функция мембраны Межклеточные контакты
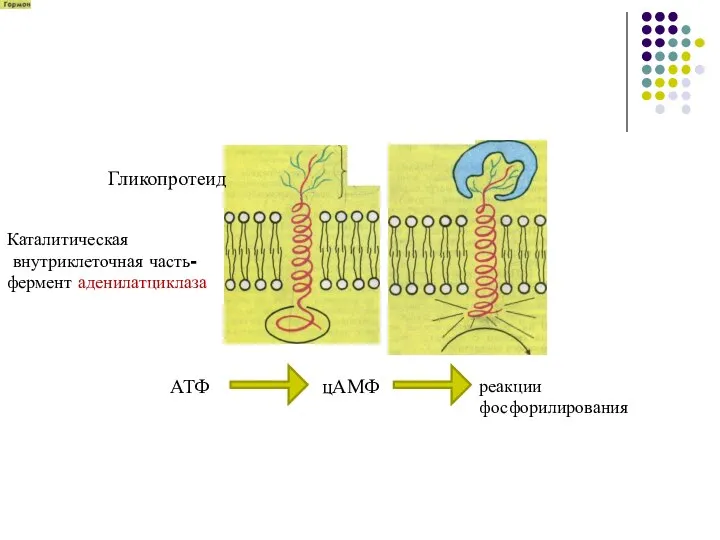
- 20. Гликопротеид Каталитическая внутриклеточная часть- фермент аденилатциклаза АТФ цАМФ реакции фосфорилирования
- 22. ЯДРО Самая крупная органелла (3-10мкм) Содержимое ядра отделено от цитоплазмы оболочкой, состоящей из двух(8нм) близкорасположенных мембран(30нм).На
- 23. Ядро В ядре находятся хромосомы – Днк в комп- лексе с гистоновыми и негистоновыми белками. Ядрышко
- 24. Эндоплазматическая сеть -ЭПС Система соединенных между собой канальцев и полостей разной величины. Их стенки представляют собой
- 25. ЭПС Шероховатая-содержит прикрепленные рибосомы, где идет синтез белка Гладкая ЭПС Здесь находятся ферменты синтеза и расщепления
- 26. Аппарат Гольджи Состоит из окруженных мембранами полостей, уложенных в стопку, в которых происходит сортировка и упаковка
- 27. Аппарат Гольджи Представляет собой образованную мембраной систему плоских цистерн, вакуолей и мелких пузырьков. В аппарат Гольджи
- 28. Синтез сложных углеводов, осуществляет их связь с белками, образуя сложные комплексы -гликопротеины Здесь образуется гликопротеин –
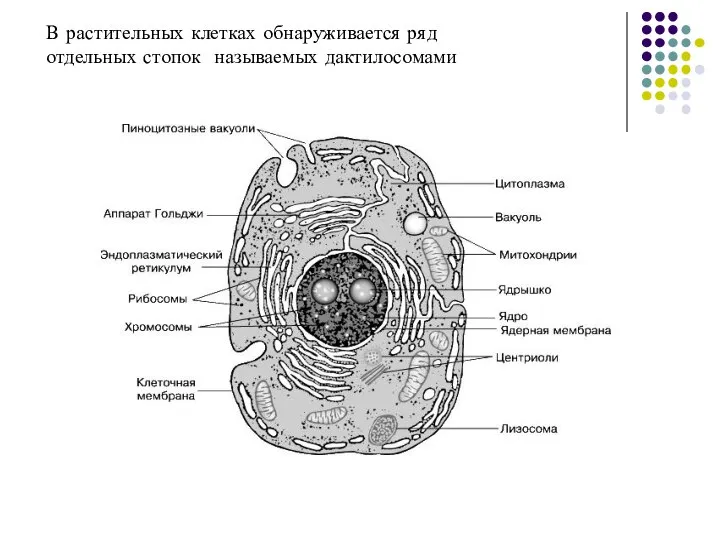
- 29. В растительных клетках обнаруживается ряд отдельных стопок называемых дактилосомами
- 30. Митохондрии Энергетические станции клетки функции которых сводятся к окислению органических соединений и использованию образовавшейся при этом
- 32. В наружной мембране присутствуют большое число каналообразующего белка порина – наружная мембрана становится проницаемой для крупных
- 33. В матриксе находятся ферментные системы окисления глюкозы и жирных кислот, а также ферменты цикла Кребса и
- 34. митохондрии Белок синтезирующий аппарат Мелкие рибосомы (Кs-55S) ДНК – 16500 нуклеотидный последовательностей Есть крупные рибосомы, но
- 35. Процесс энергообразования Превращение образовавшегося в цитозоле пирувата и жирных кислот в ацетил –СО –А. Окисление ацетил
- 36. На кристах: окислительное фосфорилирование Служит для превращения энергии запасенной в процессе гликолиза и цикла Кребса в
- 37. Потребление кислорода в качестве окислителя называют «внутриклеточным дыханием» – поэтому электронно – транспортную цепь ферментов, осуществляющих
- 38. Заключительный этап: Синтез АТФ идет в АТФ –сомах с участием АТФ – синтетаз – сложного комплекса
- 39. пластиды Это полуавтономные структуры (могут существовать относительно автономно от ядерной ДНК клетки), которые присутствуют в растительных
- 40. Выделяют три группы пластид: 1) лейкопласты. Имеют округлую форму, не окрашены и содержат питательные вещества (крахмал);
- 43. Хлоропласты представляют собой структуры, ограниченные двумя мембранами — внутренней и внешней. Внешняя мембрана, как и внутренняя,
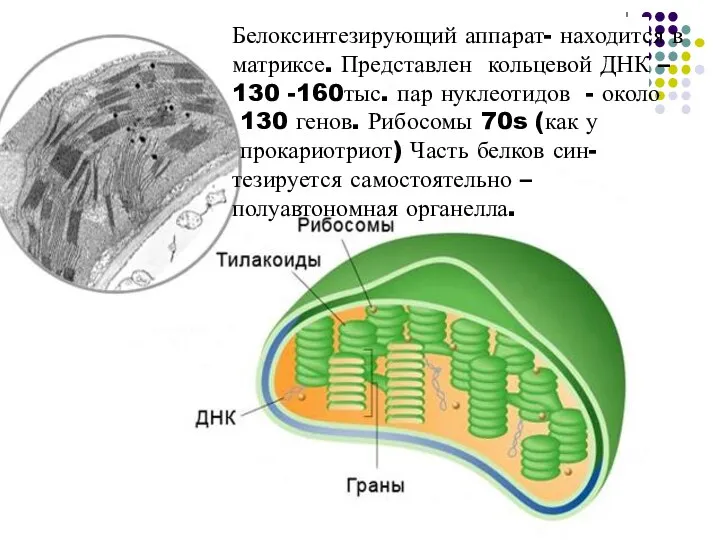
- 44. Белоксинтезирующий аппарат- находится в матриксе. Представлен кольцевой ДНК – 130 -160тыс. пар нуклеотидов - около 130
- 46. Фотосинтез; Световая фаза Энергия света поглощается хлорофиллом, который фиксирован в мембранах тилакоидов и переводит его в
- 47. Освободившаяся энергия служит для фосфорилирования АДФ в АТФ. Часть возбужденных светом электронов используется также для восстановления
- 48. Под действием солнечного света в хлоропластах происходит фотолиз воды при этом возникают электроны, которые замещают потери
- 51. Темновая фаза В составе хлоропласта есть фермент, который катализирует соединение молекулы углекислого газа с производным сахара
- 52. Лизосомы и вакуоли- учить самостоятельно
- 53. Немембранные органеллы клетки
- 54. Опорно – двигательная система клетки Жгутики Микрофиламенты микротрубочки
- 57. МИКРОТРУБОЧКА(microtubula), полая цилиндрич. структура клеток эукариотных организмов. Дл. от 100 нм до 1 млн. нм, диам.
- 59. Микрофиламенты – тонкие белковые нити диаметром 5-8 нм, встречающиеся практически во всех типах клеток. Они могут
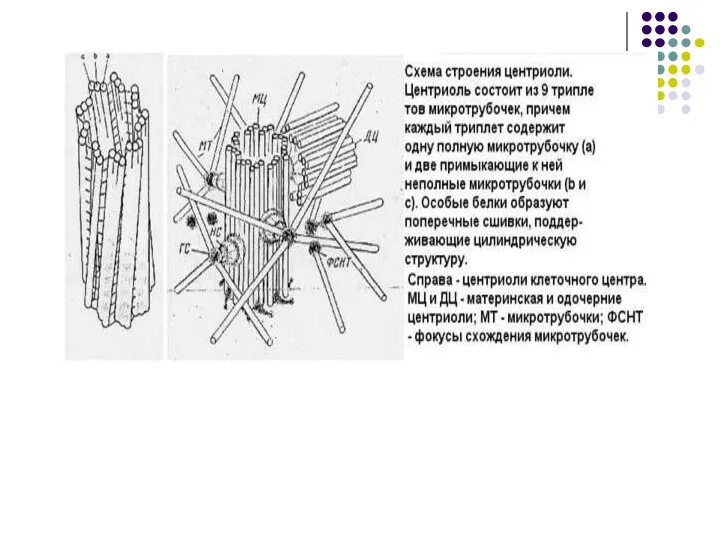
- 60. Клеточный центр Центриоль –цилиндр по окружности которого располагаются 9 триплетов микротрубочек
- 63. Скачать презентацию
Слайд 2КЛЕТОЧНАЯ ТЕОРИЯ
Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа
КЛЕТОЧНАЯ ТЕОРИЯ
Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа

Слайд 3Прокариоты
Сине-зеленые водоросли и бактерии
Бактерии:
Кольцевидная ДНК
Отсутствует ядерная оболочка
Отсутствуют аппарат Гольджи и митохондрии
Жгутики простые
Прокариоты
Сине-зеленые водоросли и бактерии
Бактерии:
Кольцевидная ДНК
Отсутствует ядерная оболочка
Отсутствуют аппарат Гольджи и митохондрии
Жгутики простые

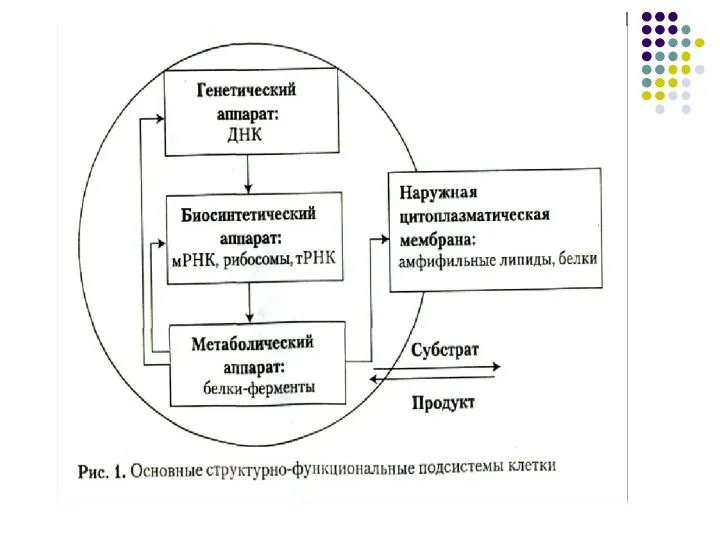
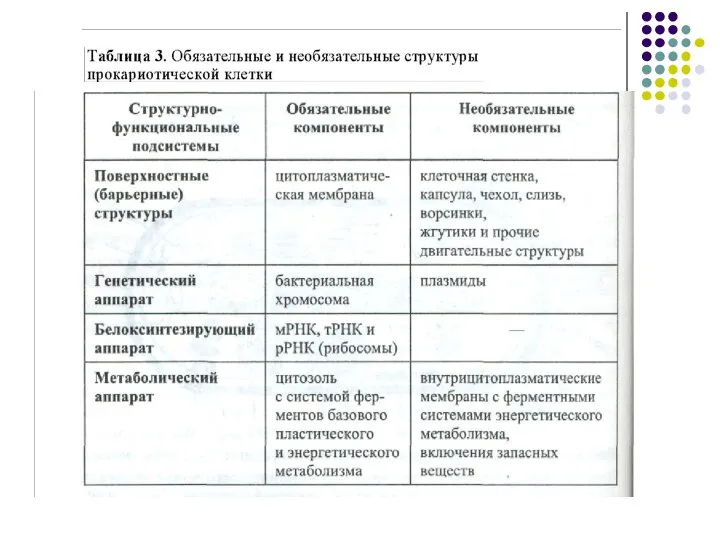
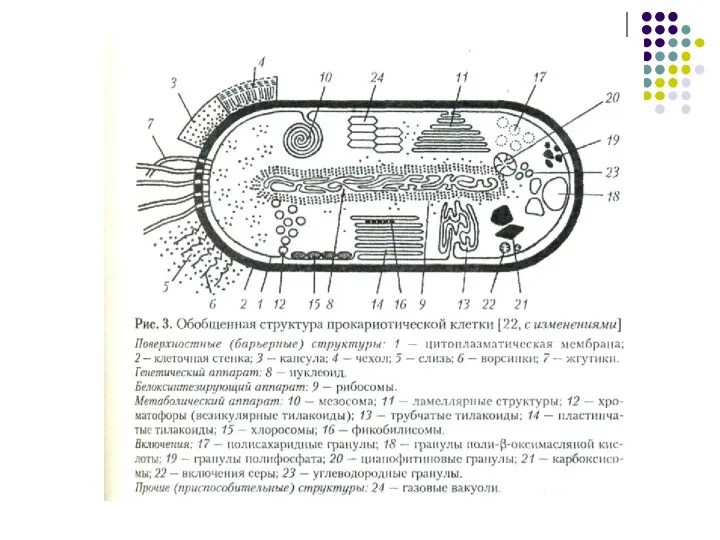
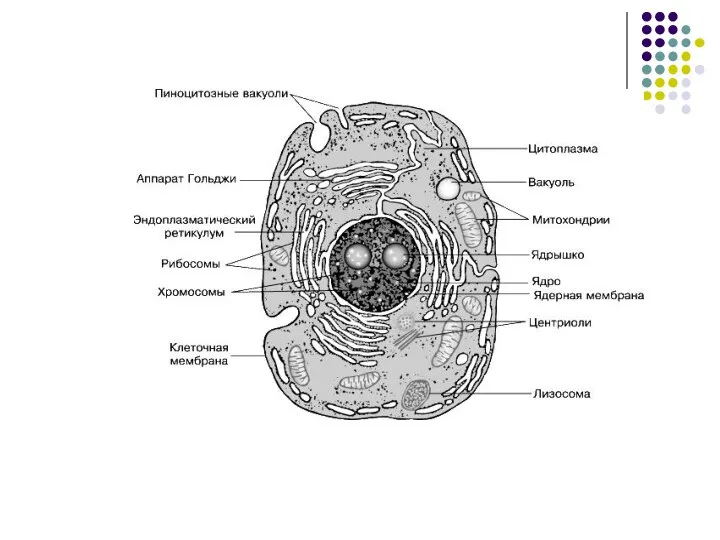
Слайд 7Эукариотическая клетка
Эукариотическая клетка
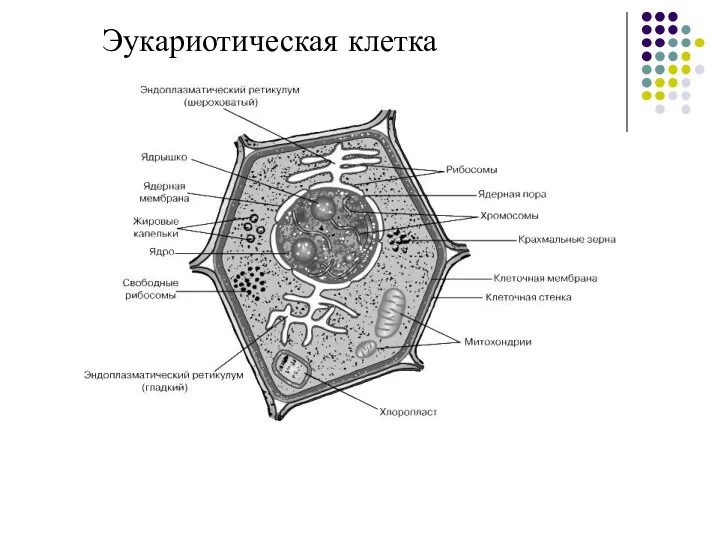
Слайд 9Плазматическая мембрана
Давсон, Даниэлли -1930г.
Робертсон – 1935г.
Гипотеза о строении элементарной мембраны:
1.Все мембраны имеют
Плазматическая мембрана
Давсон, Даниэлли -1930г.
Робертсон – 1935г.
Гипотеза о строении элементарной мембраны:
1.Все мембраны имеют

Слайд 10Жидкостно – мозаическая структура плазматической мембраны
Основная гипотеза была сформулирована в 1972 году
Жидкостно – мозаическая структура плазматической мембраны
Основная гипотеза была сформулирована в 1972 году

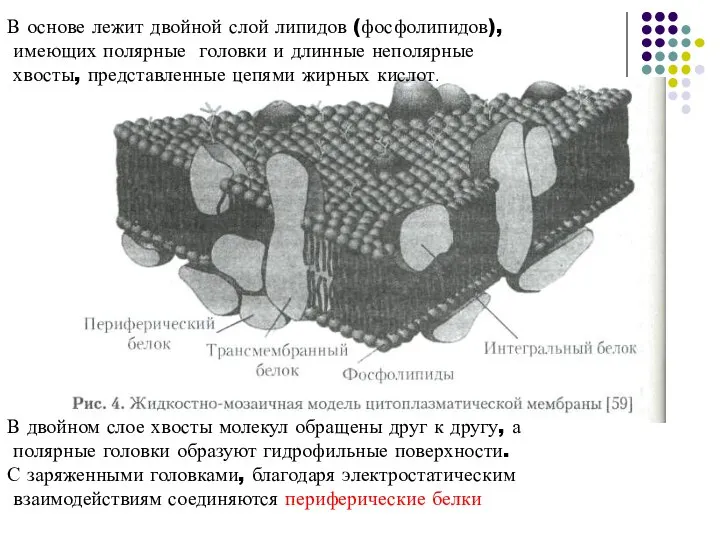
Слайд 11В основе лежит двойной слой липидов (фосфолипидов),
имеющих полярные головки и длинные
В основе лежит двойной слой липидов (фосфолипидов),
имеющих полярные головки и длинные
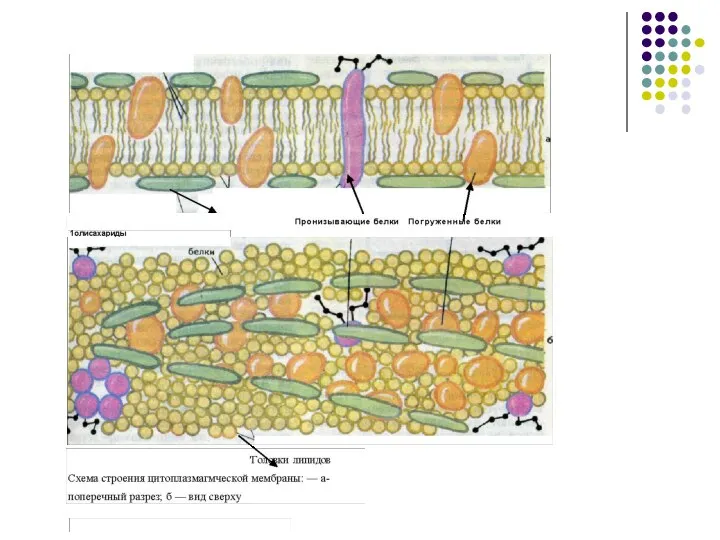
Слайд 12Белки мембран различаются по своему положению в мембране:
Они могут глубоко проникать
Белки мембран различаются по своему положению в мембране:
Они могут глубоко проникать

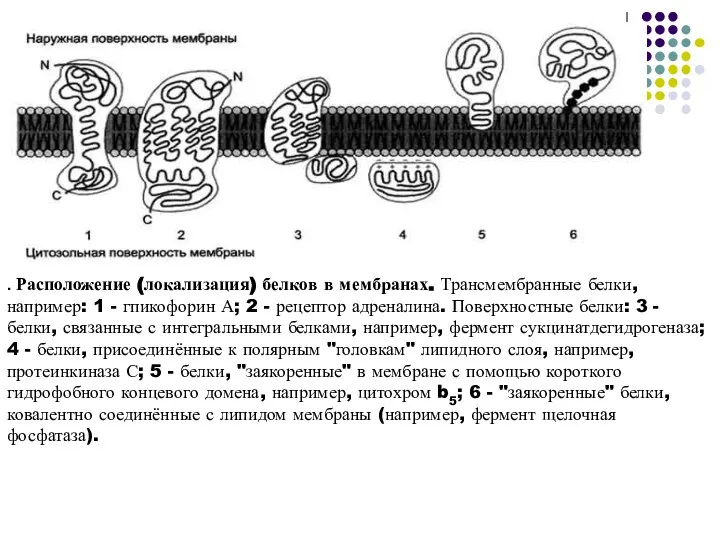
Слайд 13. Расположение (локализация) белков в мембранах. Трансмембранные белки, например: 1 - гпикофорин А;
. Расположение (локализация) белков в мембранах. Трансмембранные белки, например: 1 - гпикофорин А;
Слайд 14Поверхностные белки часто прикрепляются к мембране, взаимодействуя с интегральными
Ряд пищеварительных ферментов, участвующих
Поверхностные белки часто прикрепляются к мембране, взаимодействуя с интегральными
Ряд пищеварительных ферментов, участвующих

Слайд 15Большая часть погруженных белков -ферменты
Интегральные (погруженные ) расположены в определенном порядке, так
Большая часть погруженных белков -ферменты
Интегральные (погруженные ) расположены в определенном порядке, так

Слайд 17
Функции мембраны обеспечиваются благодаря избирательной проницаемости
Транспортная
диффузия (от меньшей концентрации к большей)
Функции мембраны обеспечиваются благодаря избирательной проницаемости
Транспортная
диффузия (от меньшей концентрации к большей)

Слайд 18D) Активный транспорт - двусторонний
перенос ( калий – натриевый насос)
Натрий-калий зависимая
D) Активный транспорт - двусторонний
перенос ( калий – натриевый насос)
Натрий-калий зависимая
Слайд 19Эндоцитоз (фаго – и пиноцитоз)
Экзоцитоз
Рецепторная функция мембраны
Межклеточные контакты
Эндоцитоз (фаго – и пиноцитоз)
Экзоцитоз
Рецепторная функция мембраны
Межклеточные контакты

Слайд 20
Гликопротеид
Каталитическая
внутриклеточная часть-
фермент аденилатциклаза
АТФ
цАМФ
реакции
фосфорилирования
Гликопротеид
Каталитическая
внутриклеточная часть-
фермент аденилатциклаза
АТФ
цАМФ
реакции
фосфорилирования
Слайд 22ЯДРО
Самая крупная органелла (3-10мкм)
Содержимое ядра отделено от цитоплазмы оболочкой, состоящей из двух(8нм)
ЯДРО
Самая крупная органелла (3-10мкм)
Содержимое ядра отделено от цитоплазмы оболочкой, состоящей из двух(8нм)

Слайд 23Ядро
В ядре находятся хромосомы – Днк в комп-
лексе с гистоновыми и негистоновыми
Ядро
В ядре находятся хромосомы – Днк в комп-
лексе с гистоновыми и негистоновыми

Слайд 24Эндоплазматическая сеть -ЭПС
Система соединенных между собой канальцев и полостей разной величины. Их
Эндоплазматическая сеть -ЭПС
Система соединенных между собой канальцев и полостей разной величины. Их

Слайд 25ЭПС
Шероховатая-содержит прикрепленные рибосомы, где идет синтез белка
Гладкая ЭПС
Здесь находятся ферменты синтеза
ЭПС
Шероховатая-содержит прикрепленные рибосомы, где идет синтез белка
Гладкая ЭПС
Здесь находятся ферменты синтеза

Слайд 26Аппарат Гольджи
Состоит из окруженных мембранами полостей, уложенных в стопку, в которых происходит
Аппарат Гольджи
Состоит из окруженных мембранами полостей, уложенных в стопку, в которых происходит

Слайд 27Аппарат Гольджи
Представляет собой образованную мембраной систему плоских цистерн, вакуолей и мелких пузырьков.
Аппарат Гольджи
Представляет собой образованную мембраной систему плоских цистерн, вакуолей и мелких пузырьков.

Слайд 28Синтез сложных углеводов, осуществляет их связь с белками, образуя сложные комплексы -гликопротеины
Здесь
Синтез сложных углеводов, осуществляет их связь с белками, образуя сложные комплексы -гликопротеины
Здесь

Слайд 29В растительных клетках обнаруживается ряд
отдельных стопок называемых дактилосомами
В растительных клетках обнаруживается ряд
отдельных стопок называемых дактилосомами
Слайд 30Митохондрии
Энергетические станции клетки функции которых сводятся к окислению органических соединений и использованию
Митохондрии
Энергетические станции клетки функции которых сводятся к окислению органических соединений и использованию

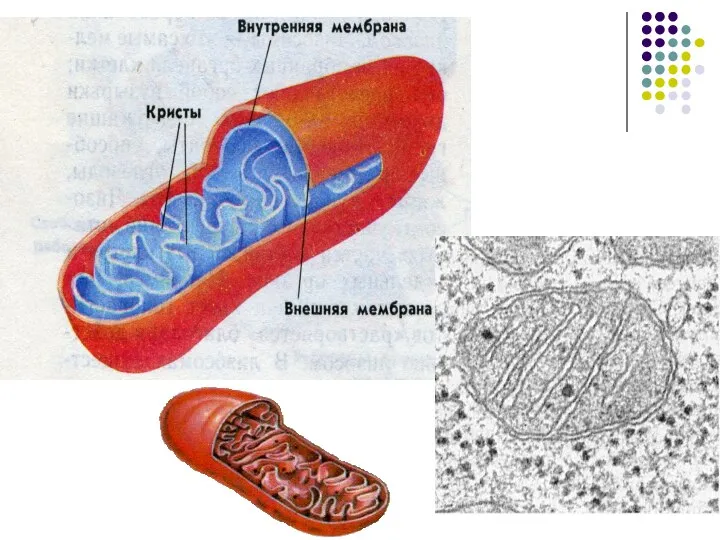
Слайд 32В наружной мембране присутствуют большое число каналообразующего белка порина – наружная мембрана
В наружной мембране присутствуют большое число каналообразующего белка порина – наружная мембрана

Слайд 33В матриксе находятся ферментные системы окисления глюкозы и жирных кислот, а также
В матриксе находятся ферментные системы окисления глюкозы и жирных кислот, а также

Слайд 34митохондрии
Белок синтезирующий аппарат
Мелкие рибосомы (Кs-55S)
ДНК – 16500 нуклеотидный последовательностей
Есть крупные рибосомы, но
митохондрии
Белок синтезирующий аппарат
Мелкие рибосомы (Кs-55S)
ДНК – 16500 нуклеотидный последовательностей
Есть крупные рибосомы, но

Слайд 35Процесс энергообразования
Превращение образовавшегося в цитозоле пирувата и жирных кислот в ацетил –СО
Процесс энергообразования
Превращение образовавшегося в цитозоле пирувата и жирных кислот в ацетил –СО

Слайд 36На кристах: окислительное
фосфорилирование
Служит для превращения энергии запасенной в процессе гликолиза и цикла
На кристах: окислительное
фосфорилирование
Служит для превращения энергии запасенной в процессе гликолиза и цикла

Слайд 37Потребление кислорода в качестве окислителя называют «внутриклеточным дыханием» – поэтому электронно –
Потребление кислорода в качестве окислителя называют «внутриклеточным дыханием» – поэтому электронно –

Слайд 38Заключительный этап:
Синтез АТФ идет в АТФ –сомах с участием АТФ – синтетаз
Заключительный этап:
Синтез АТФ идет в АТФ –сомах с участием АТФ – синтетаз

Слайд 39пластиды
Это полуавтономные структуры (могут существовать относительно автономно от ядерной ДНК клетки), которые
пластиды
Это полуавтономные структуры (могут существовать относительно автономно от ядерной ДНК клетки), которые

Слайд 40Выделяют три группы пластид:
1) лейкопласты. Имеют округлую форму, не окрашены и содержат
Выделяют три группы пластид: 1) лейкопласты. Имеют округлую форму, не окрашены и содержат

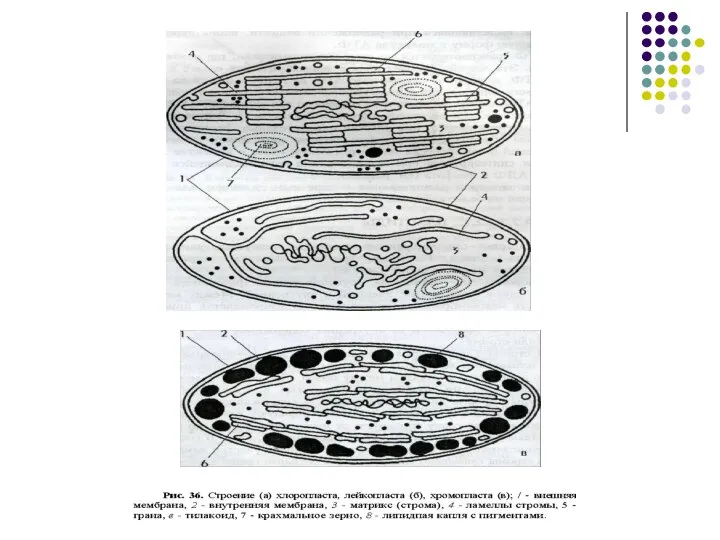
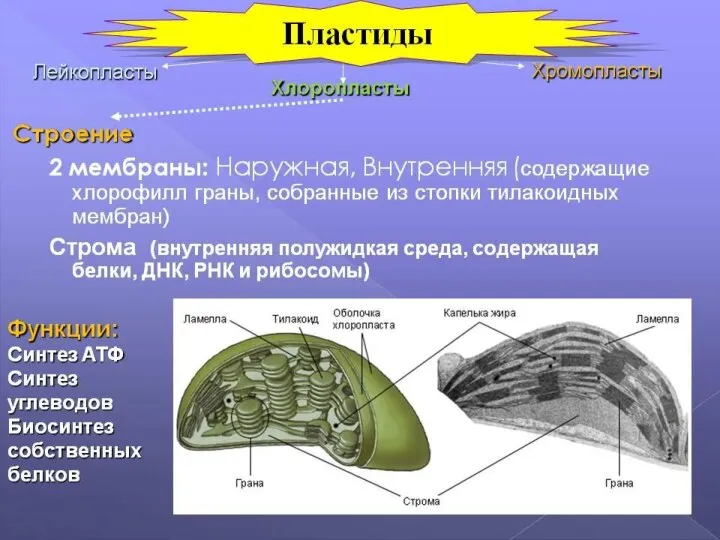
Слайд 43Хлоропласты представляют собой структуры, ограниченные двумя мембранами — внутренней и внешней. Внешняя
Хлоропласты представляют собой структуры, ограниченные двумя мембранами — внутренней и внешней. Внешняя
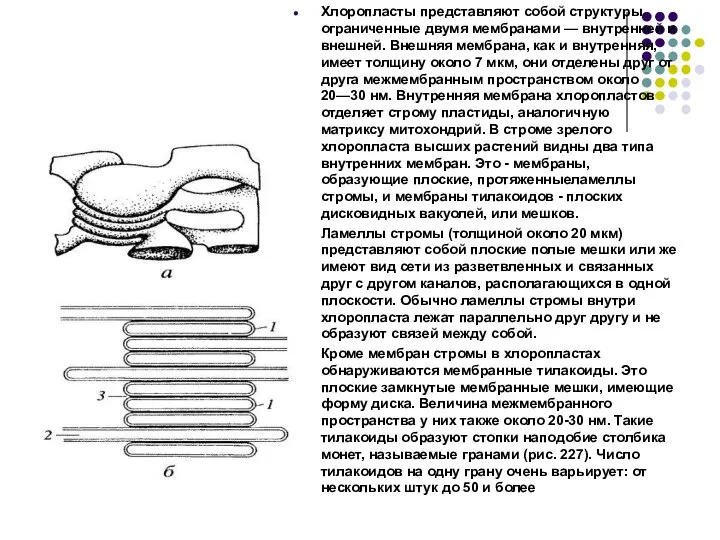
Слайд 44Белоксинтезирующий аппарат- находится в матриксе. Представлен кольцевой ДНК – 130 -160тыс. пар
Белоксинтезирующий аппарат- находится в матриксе. Представлен кольцевой ДНК – 130 -160тыс. пар

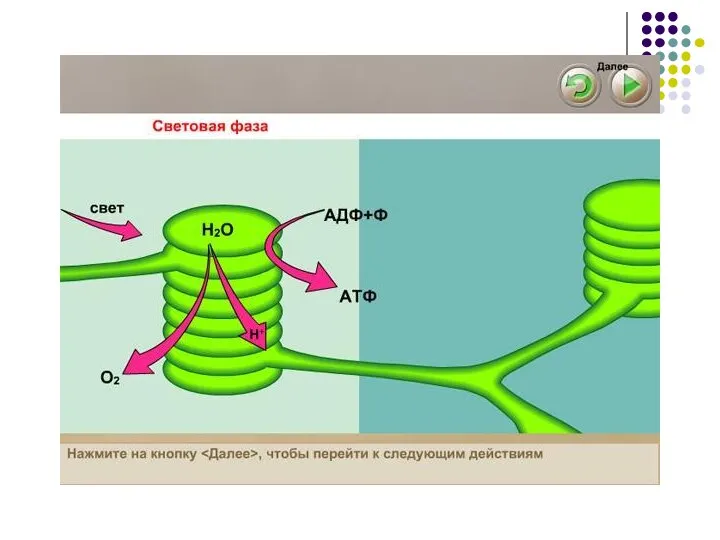
Слайд 46Фотосинтез; Световая фаза
Энергия света поглощается хлорофиллом, который фиксирован в мембранах тилакоидов и
Фотосинтез; Световая фаза
Энергия света поглощается хлорофиллом, который фиксирован в мембранах тилакоидов и

Слайд 47Освободившаяся энергия служит для фосфорилирования АДФ в АТФ.
Часть возбужденных светом электронов используется
Освободившаяся энергия служит для фосфорилирования АДФ в АТФ.
Часть возбужденных светом электронов используется

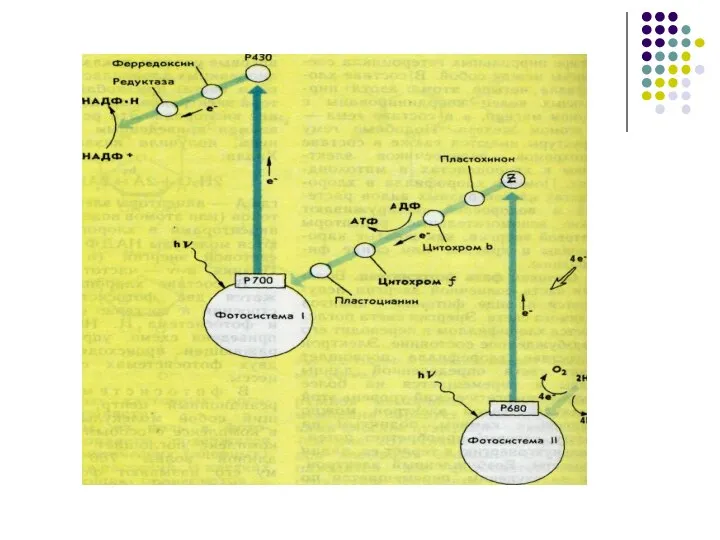
Слайд 48Под действием солнечного света в хлоропластах происходит фотолиз воды при этом возникают
Под действием солнечного света в хлоропластах происходит фотолиз воды при этом возникают

Слайд 51Темновая фаза
В составе хлоропласта есть фермент, который катализирует соединение молекулы углекислого газа
Темновая фаза
В составе хлоропласта есть фермент, который катализирует соединение молекулы углекислого газа

Слайд 52Лизосомы и вакуоли- учить самостоятельно
Лизосомы и вакуоли- учить самостоятельно

Слайд 53Немембранные органеллы клетки
Немембранные органеллы клетки

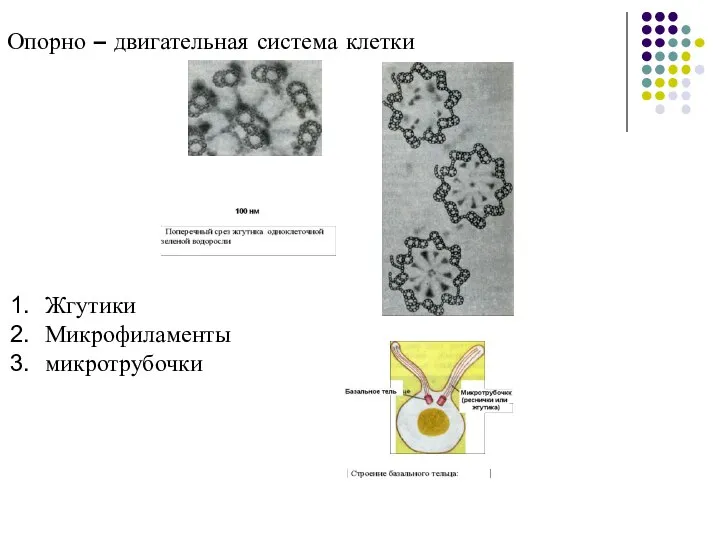
Слайд 54Опорно – двигательная система клетки
Жгутики
Микрофиламенты
микротрубочки
Опорно – двигательная система клетки
Жгутики
Микрофиламенты
микротрубочки
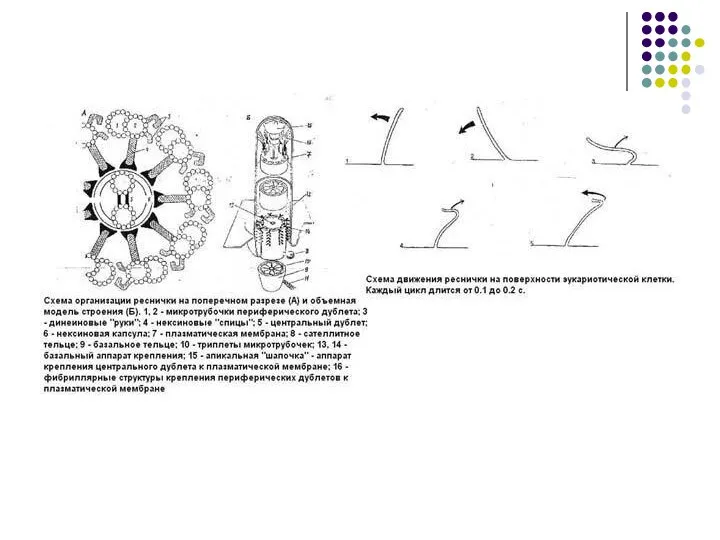
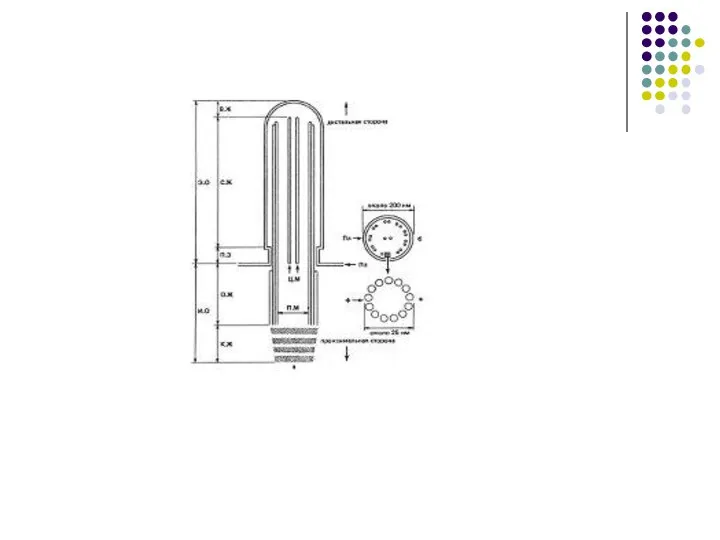
Слайд 57МИКРОТРУБОЧКА(microtubula), полая цилиндрич. структура клеток эукариотных организмов. Дл. от 100 нм до
МИКРОТРУБОЧКА(microtubula), полая цилиндрич. структура клеток эукариотных организмов. Дл. от 100 нм до

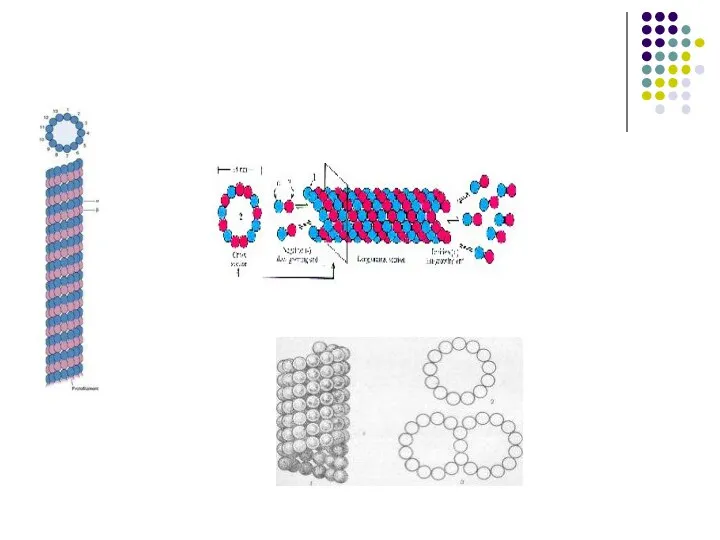
Слайд 59Микрофиламенты – тонкие белковые нити диаметром 5-8 нм, встречающиеся практически во всех
Микрофиламенты – тонкие белковые нити диаметром 5-8 нм, встречающиеся практически во всех

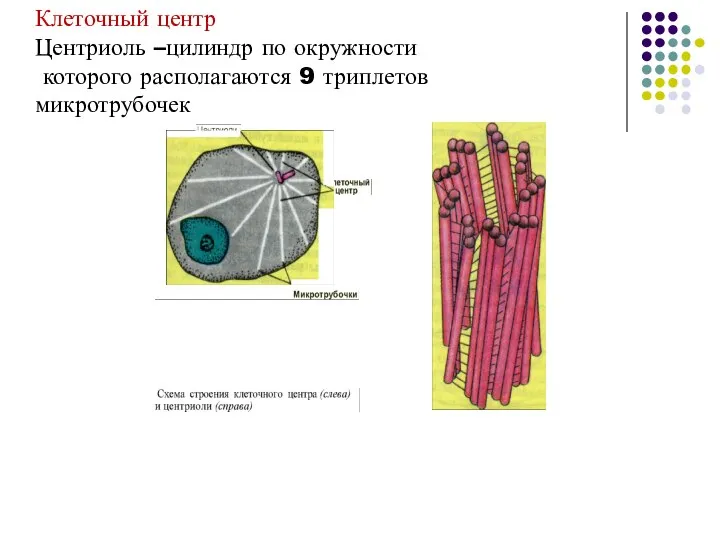
Слайд 60Клеточный центр
Центриоль –цилиндр по окружности
которого располагаются 9 триплетов
микротрубочек
Клеточный центр
Центриоль –цилиндр по окружности
которого располагаются 9 триплетов
микротрубочек
 Белый медведь
Белый медведь Клещи. Cпособы защиты от клещей
Клещи. Cпособы защиты от клещей Плоские черви
Плоские черви Биотические факторы среды. Типы взаимодействий между организмами
Биотические факторы среды. Типы взаимодействий между организмами Индукция толерогенных свойств дендритных клеток мышей линий BALB/C и C57BL/6
Индукция толерогенных свойств дендритных клеток мышей линий BALB/C и C57BL/6 Morphogical Characteristics
Morphogical Characteristics Внешнее строение птиц. Опорно-двигательная система птиц
Внешнее строение птиц. Опорно-двигательная система птиц Презентация на тему ХИЩНИКИ
Презентация на тему ХИЩНИКИ  Тварини Євразії
Тварини Євразії Клеточный цикл. Митоз. Наблюдение митоза в клетках кончика корешка лука на готовых микропрепаратах
Клеточный цикл. Митоз. Наблюдение митоза в клетках кончика корешка лука на готовых микропрепаратах MINPIN
MINPIN Презентация на тему Особенности высшей нервной деятельности человека
Презентация на тему Особенности высшей нервной деятельности человека  Центральный отдел слухового анализатора. Физиология слухового анализатора
Центральный отдел слухового анализатора. Физиология слухового анализатора Общая микробиология. Лекция 1
Общая микробиология. Лекция 1 Нервная система
Нервная система Обмен веществ и энергии
Обмен веществ и энергии Галофобы
Галофобы Влияние окружающей среды на развивающийся организм в эмбриональный период
Влияние окружающей среды на развивающийся организм в эмбриональный период Презентация на тему КЛАССИФИКАЦИЯ ЖИВОТНЫХ
Презентация на тему КЛАССИФИКАЦИЯ ЖИВОТНЫХ  Основные принципы проектирования архивов клонов
Основные принципы проектирования архивов клонов Международный День Прав Животных
Международный День Прав Животных Строение белковой молекулы
Строение белковой молекулы Принципы и механизмы регуляции онтогенеза. Лекция 4
Принципы и механизмы регуляции онтогенеза. Лекция 4 Подготовка к ВПР (окружающий мир, 4 класс)
Подготовка к ВПР (окружающий мир, 4 класс) Галерея. История развития биологии
Галерея. История развития биологии Фотосинтез и дыхание растений
Фотосинтез и дыхание растений Фосфорамидный метод
Фосфорамидный метод Жизнь леса
Жизнь леса