- Физические основы преобразования и аккумуляции энергии в биологических системах

Содержание
- 2. Клеточная мембрана –полупроницаемый барьер, отделяющий цитоплазму клеток от окружающей среды Плазматическая мембрана. Фосфолипид 1 R 2
- 3. Биофизика мембран. Основные задачи биофизики мембран: Строение мембран, динамические свойства мембранной структуры, определяющие ее функциональность Мембранный
- 4. Образование мембранных структур. Изотерма площадь-давление для монослоя длинноцепочечных липидов и схема расположения липидных фаз вода-воздух (рассматривается
- 5. Образование мембранных структур. Создание искусственных бислойных мембран позволяет моделировать и исследовать свойства биомембран Сферические бислойные мембраны
- 6. Образование мембранных структур. Этапы формирования бислойной мембраны: 1 2 3 Толстая многослойная мембрана (~ > 100
- 7. Образование мембранных структур. Поведение различных веществ в растворах подчиняется адсорбционному уравнению Гиббса: где σ – поверхностное
- 8. Фазовые переходы в мембранах. Природные и искусственные мембраны могут находиться в 2-х основных фазовых состояниях: Твердый
- 9. ΔG = ΔH– TΔS Фазовые переходы в мембранах. Фазовый переход первого рода, так как скачком изменяется
- 10. Кооперативность фазовых переходов в мембранах. Для оценки степени кооперативности сравнивают изменение внутренней энергии (ΔH) и измеряемую
- 11. Мембранный транспорт Мембранный транспорт Пассивный - транспорт веществ по градиенту концентрации, осуществляющийся за счет диффузии не
- 12. Классификация видов мембранного транспорта
- 13. Энергия пассивного транспорта создается различными градиентами: концентрационным осмотическим электрическим градиентом гидростатического давления жидкости электрохимическим (совокупность концентрационного
- 14. Диффузия – это самопроизвольный процесс проникновения вещества из области большей в область меньшей его концентрации в
- 15. Пассивный мембранный транспорт. Диффузия. Первый закон Фика: (поток вещества пропорционален градиенту концентрации) В случае стационарного потока
- 16. Уравнение Коллендера-Бернульда где С1 и С2 – концентрации вещества по разные стороны мембраны Р – коэффициент
- 17. График простой диффузии Скорость переноса Концентрация
- 18. Молекулы, имеющие примерно одинаковую массу и размер, поступают через мембрану тем легче, чем выше их растворимость
- 19. В ряде случаев скорость диффузии некоторых водорастворимых веществ во много раз выше, чем скорость диффузии этих
- 20. Схема облегченной диффузии, осуществляемой при помощи белка-переносчика.
- 21. Транспорт ионов. Движущей силой диффузии служит разность химических потенциалов данного вещества в двух областях. Стандартный химический
- 22. Ионные каналы биомембраны Ионные каналы биомембраны - это специализированные селективные поры, образованные белковыми молекулами, предназначенными для
- 23. Строение натриевого ионного канала мембраны
- 24. Активный мембранный транспорт. Натрий-калиевый насос. Рабочий цикл димера Na+ насоса в режиме Na+/K+ обмена Большая субъединица
- 25. E- фермент, Х – АТФ, Z- АДФ, Y(Ф)- фосфат, i- внутренняя сторона мембраны, e – внешняя
- 26. Активный мембранный транспорт. Натрий-калиевый насос.
- 27. Кинетика облегченной диффузии через мембраны при участии белка-переносчика и каналообразующего белка Скорость транспорта
- 28. Ферментативный перенос описывается уравнением Михаэлиса-Ментен . Поток вещества М равен возможной максимальной скорости Vmax реакции в
- 29. Обменная диффузия - это ферментативный перенос веществ через мембрану как с внешней, так и с внутренней
- 30. Уравнение, описывающее осмотический перенос воды. Где - количество воды, проходящей через мембрану площадью S за единицу
- 31. Где r – радиус поры l – длина поры η – вязкость жидкости Р1-Р2 – разность
- 32. Основные этапы Na+ - K+-насоса : образование комплекса фермента с АТФ на внутренней поверхности мембраны (эта
- 33. Внешняя среда Внутренняя среда МЕМБРАНА АТФ АДФ+Фн 2К + 3Na + 2Mg 2+ АТФ-аза
- 34. Основные этапы Са2+-насоса : Первый этап: связывание Са2+ с комплексом Mg-АТФазы на наружной поверхности саркоплазматической мембраны.
- 35. КЛЕТКА МЕМБРАНА МЕЖКЛЕТОЧНОЕ ПРОСТРАНСТВО АТФ АДФ+Фн Са 2+ Mg 2+ АТФ-аза
- 36. Образующийся в цикле Кребса восстановленный никотинамидадениндинуклеотид (NADH) подхватывается плавающим по мембране первым дыхательным комплексом (I) и
- 37. Внешняя среда Сопрягающая мембрана Внутренняя среда субстрат АТФ АДФ+Фн е е 2Н 2Н 2Н + +
- 38. Организация дыхательной цепи Комплекс I содержит FMN (флавинмононуклеотид) и не менее пяти железо-серных белков. Комплекс II
- 39. Организация дыхательной цепи
- 40. Физические основы преобразования и аккумуляции энергии в биологических системах Таблица 1. Компоненты дыхательной цепи В дыхательной
- 41. Константа равновесия К непосредственно связана с изменением свободной энергии реакции в стандартных условиях ΔG° (ΔG° =
- 42. Причины высоко экзоэргического характера гидролиза АТФ: сильное отталкивание отрицательно заряженных атомов кислорода, которое ослабевает при гидролитическом
- 43. Механизм синтеза АТФ АТФ-синтаза состоит из двух частей: встроенного в мембрану протонного канала и каталитической белковой
- 44. Синтез АТФ
- 45. Митохондрии, перенос электронов Митохондриальная цепь переноса электронов. Комплекс I содержит FMN и не менее пяти железо-серных
- 46. Направление процесса и сопряженные реакции Направление химической реакции определяется значением ΔG. Если эта величина отрицательна, то
- 47. Механизмы, обеспечивающие протекание сопряженных реакций. Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата является эндергонической: (1)
- 48. Свободная энергия гидролиза некоторых органических фосфатов
- 49. АТФ - молекула, богатая энергией, поскольку она содержит две высокоэнергетические (макроэргические) фосфоангидридные связи (β, γ). Они
- 50. Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифосфатов, аналогов АТФ; к ним относят
- 51. Биологическое окисление Под окислением понимают отщепление электронов, а под восстановлением - присоединение электронов. Окисление донора электронов
- 52. Стандартные окислительно-восстановительные потенциалы некоторых сопряжённых пар Величину Еº' выражают в вольтах; чем она меньше (отрицательнее), тем
- 54. Скачать презентацию
Слайд 2Клеточная мембрана –полупроницаемый барьер, отделяющий цитоплазму клеток от окружающей среды
Плазматическая мембрана.
Фосфолипид
1 R
2
Клеточная мембрана –полупроницаемый барьер, отделяющий цитоплазму клеток от окружающей среды
Плазматическая мембрана.
Фосфолипид
1 R
2
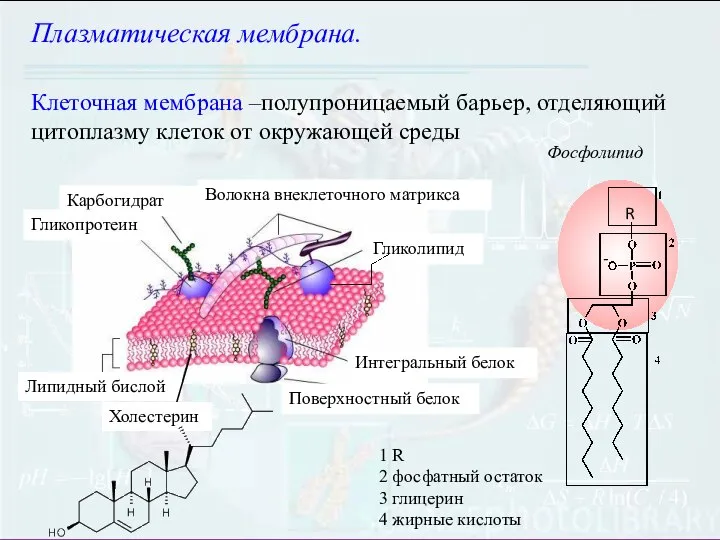
Слайд 3Биофизика мембран.
Основные задачи биофизики мембран:
Строение мембран, динамические свойства мембранной структуры, определяющие
Биофизика мембран.
Основные задачи биофизики мембран:
Строение мембран, динамические свойства мембранной структуры, определяющие

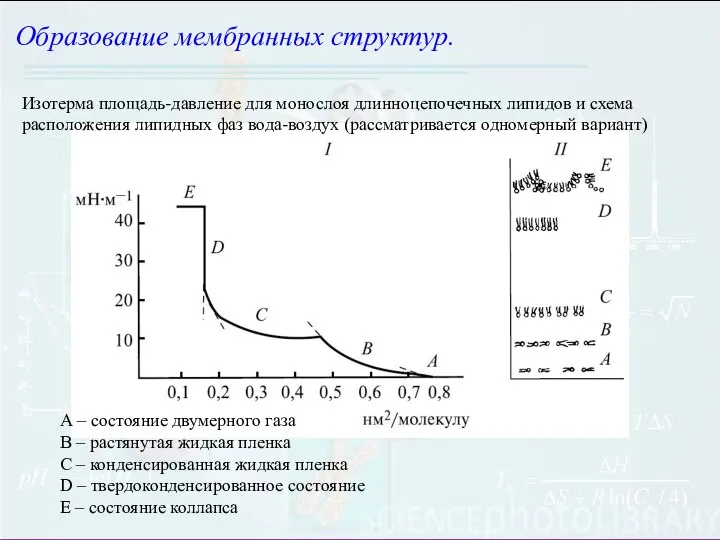
Слайд 4Образование мембранных структур.
Изотерма площадь-давление для монослоя длинноцепочечных липидов и схема расположения липидных
Образование мембранных структур.
Изотерма площадь-давление для монослоя длинноцепочечных липидов и схема расположения липидных
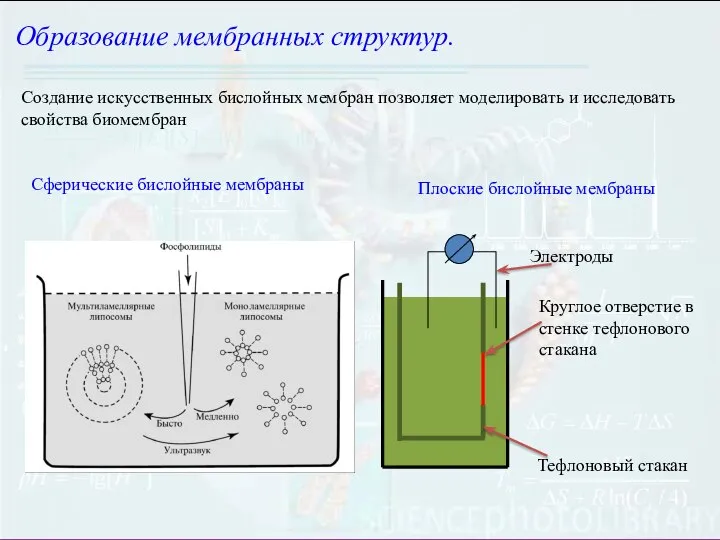
Слайд 5Образование мембранных структур.
Создание искусственных бислойных мембран позволяет моделировать и исследовать свойства биомембран
Образование мембранных структур.
Создание искусственных бислойных мембран позволяет моделировать и исследовать свойства биомембран
Слайд 6Образование мембранных структур.
Этапы формирования бислойной мембраны:
1
2
3
Толстая многослойная мембрана (~ > 100 нм)
Выпуклая
Образование мембранных структур.
Этапы формирования бислойной мембраны:
1
2
3
Толстая многослойная мембрана (~ > 100 нм)
Выпуклая

Слайд 7Образование мембранных структур.
Поведение различных веществ в растворах подчиняется
адсорбционному уравнению Гиббса:
где σ
Образование мембранных структур.
Поведение различных веществ в растворах подчиняется
адсорбционному уравнению Гиббса:
где σ

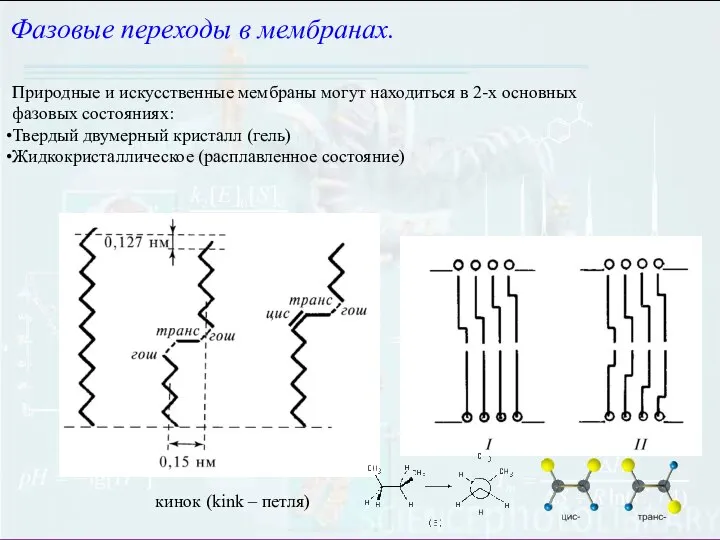
Слайд 8Фазовые переходы в мембранах.
Природные и искусственные мембраны могут находиться в 2-х основных
Фазовые переходы в мембранах.
Природные и искусственные мембраны могут находиться в 2-х основных
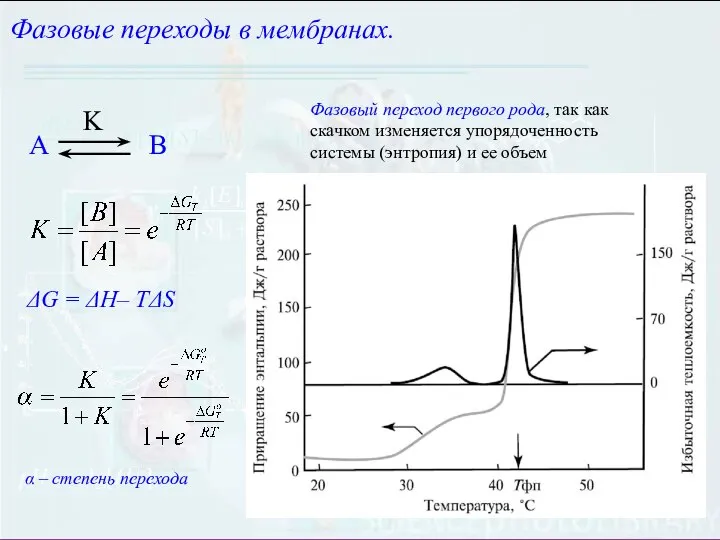
Слайд 9ΔG = ΔH– TΔS
Фазовые переходы в мембранах.
Фазовый переход первого рода, так как
ΔG = ΔH– TΔS
Фазовые переходы в мембранах.
Фазовый переход первого рода, так как
Слайд 10Кооперативность фазовых переходов в мембранах.
Для оценки степени кооперативности сравнивают изменение внутренней энергии
Кооперативность фазовых переходов в мембранах.
Для оценки степени кооперативности сравнивают изменение внутренней энергии

Слайд 11Мембранный транспорт
Мембранный транспорт
Пассивный - транспорт веществ по градиенту концентрации, осуществляющийся за счет
Мембранный транспорт
Мембранный транспорт
Пассивный - транспорт веществ по градиенту концентрации, осуществляющийся за счет

Слайд 12Классификация видов мембранного транспорта
Классификация видов мембранного транспорта

Слайд 13Энергия пассивного транспорта создается различными градиентами:
концентрационным
осмотическим
электрическим
градиентом гидростатического давления жидкости
электрохимическим (совокупность концентрационного и
Энергия пассивного транспорта создается различными градиентами:
концентрационным
осмотическим
электрическим
градиентом гидростатического давления жидкости
электрохимическим (совокупность концентрационного и

Слайд 14Диффузия –
это самопроизвольный процесс проникновения вещества из области большей в область
Диффузия –
это самопроизвольный процесс проникновения вещества из области большей в область

Слайд 15Пассивный мембранный транспорт. Диффузия.
Первый закон Фика:
(поток вещества пропорционален градиенту концентрации)
В случае стационарного
Пассивный мембранный транспорт. Диффузия.
Первый закон Фика:
(поток вещества пропорционален градиенту концентрации)
В случае стационарного

Слайд 16Уравнение Коллендера-Бернульда
где
С1 и С2 – концентрации вещества по разные стороны мембраны
Р –
Уравнение Коллендера-Бернульда
где
С1 и С2 – концентрации вещества по разные стороны мембраны
Р –

Слайд 17График простой диффузии
Скорость переноса
Концентрация
График простой диффузии
Скорость переноса
Концентрация
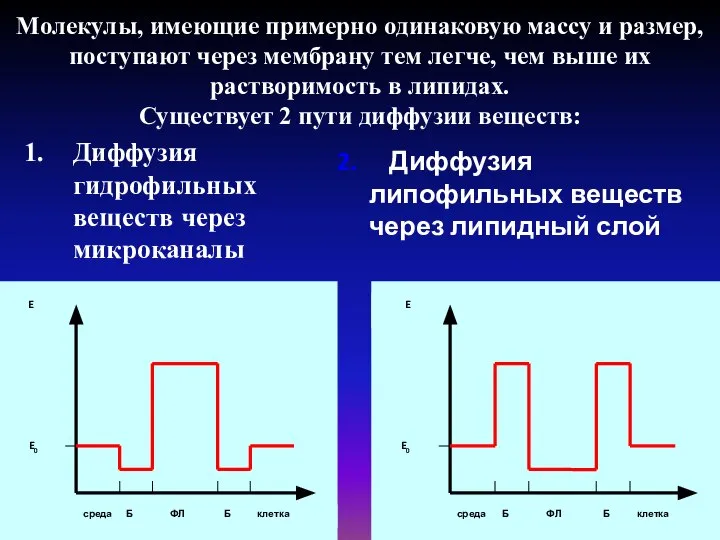
Слайд 18Молекулы, имеющие примерно одинаковую массу и размер, поступают через мембрану тем легче,
Молекулы, имеющие примерно одинаковую массу и размер, поступают через мембрану тем легче,
Слайд 19 В ряде случаев скорость диффузии некоторых
водорастворимых веществ во много раз
В ряде случаев скорость диффузии некоторых водорастворимых веществ во много раз

Слайд 20
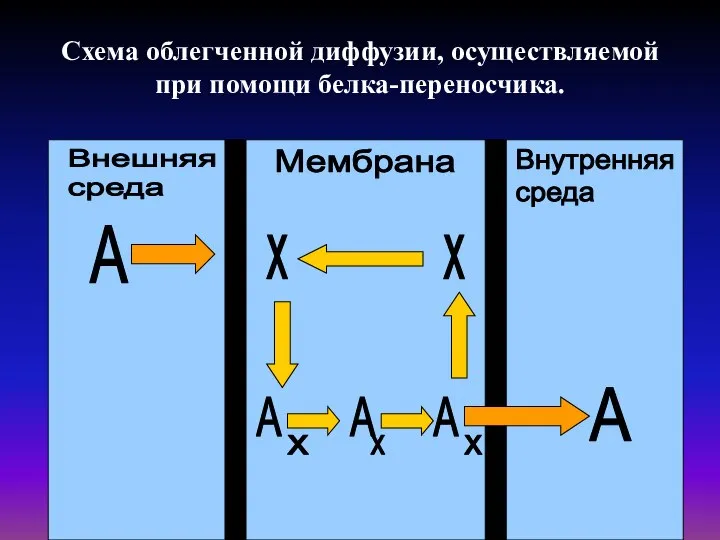
Схема облегченной диффузии, осуществляемой при помощи белка-переносчика.
Схема облегченной диффузии, осуществляемой при помощи белка-переносчика.
Слайд 21Транспорт ионов.
Движущей силой диффузии служит разность химических потенциалов данного вещества в двух
Транспорт ионов.
Движущей силой диффузии служит разность химических потенциалов данного вещества в двух

Слайд 22Ионные каналы биомембраны
Ионные каналы биомембраны - это специализированные селективные поры, образованные
Ионные каналы биомембраны
Ионные каналы биомембраны - это специализированные селективные поры, образованные

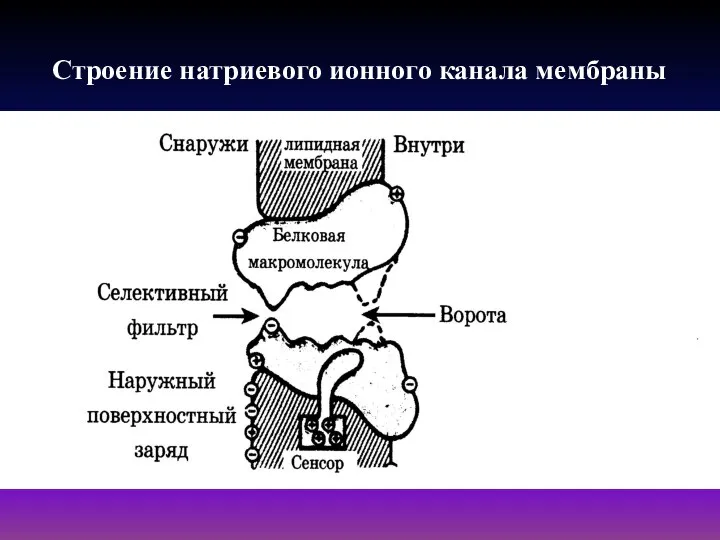
Слайд 23Строение натриевого ионного канала мембраны
Строение натриевого ионного канала мембраны
Слайд 24Активный мембранный транспорт. Натрий-калиевый насос.
Рабочий цикл димера Na+ насоса в режиме Na+/K+
Активный мембранный транспорт. Натрий-калиевый насос.
Рабочий цикл димера Na+ насоса в режиме Na+/K+
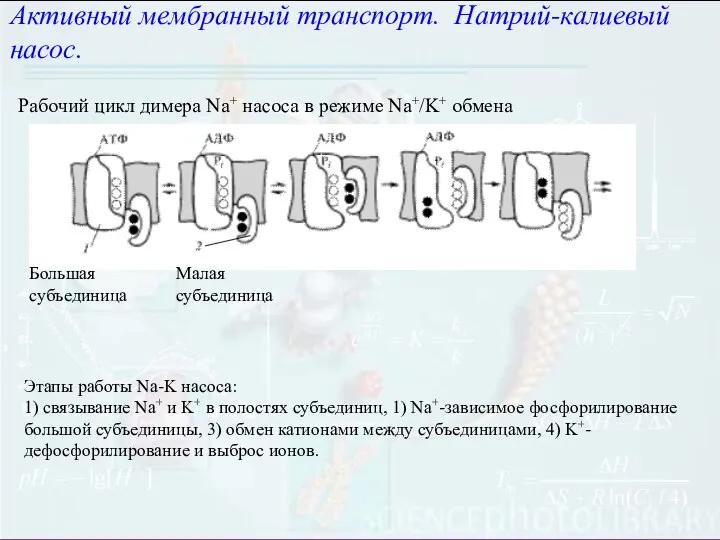
Слайд 25E- фермент, Х – АТФ, Z- АДФ, Y(Ф)- фосфат, i- внутренняя сторона
E- фермент, Х – АТФ, Z- АДФ, Y(Ф)- фосфат, i- внутренняя сторона

Слайд 26Активный мембранный транспорт. Натрий-калиевый насос.
Активный мембранный транспорт. Натрий-калиевый насос.

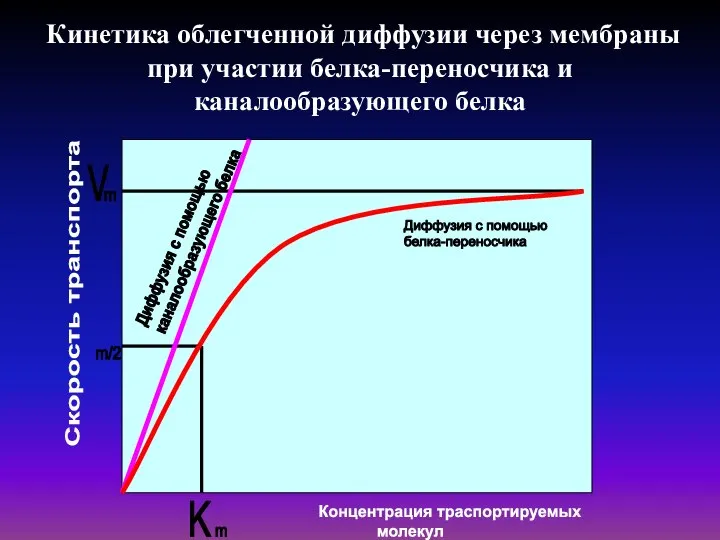
Слайд 27Кинетика облегченной диффузии через мембраны при участии белка-переносчика и каналообразующего белка
Скорость транспорта
Кинетика облегченной диффузии через мембраны при участии белка-переносчика и каналообразующего белка
Скорость транспорта
Слайд 28Ферментативный перенос описывается уравнением Михаэлиса-Ментен .
Поток вещества М равен возможной
Ферментативный перенос описывается уравнением Михаэлиса-Ментен .
Поток вещества М равен возможной

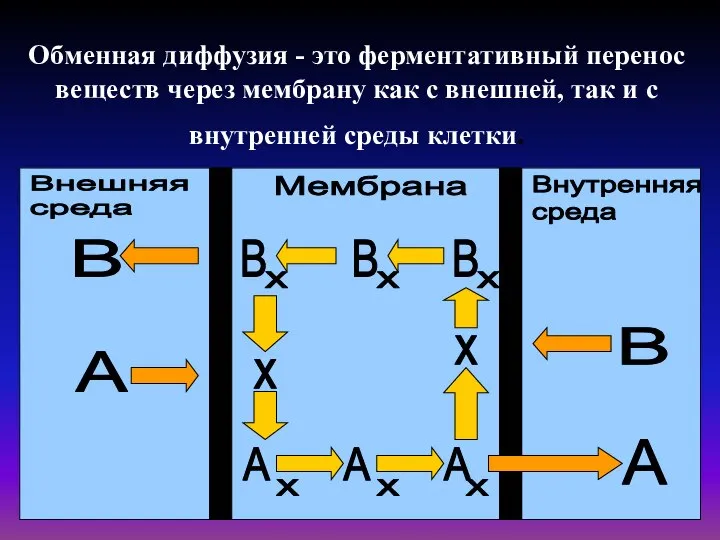
Слайд 29Обменная диффузия - это ферментативный перенос веществ через мембрану как с внешней,
Обменная диффузия - это ферментативный перенос веществ через мембрану как с внешней,
Слайд 30Уравнение, описывающее осмотический перенос воды.
Где
- количество воды, проходящей через мембрану площадью
Уравнение, описывающее осмотический перенос воды.
Где
- количество воды, проходящей через мембрану площадью

Слайд 31Где
r – радиус поры
l – длина поры
η – вязкость
Где
r – радиус поры
l – длина поры
η – вязкость

Слайд 32Основные этапы Na+ - K+-насоса :
образование комплекса фермента с АТФ на внутренней
Основные этапы Na+ - K+-насоса :
образование комплекса фермента с АТФ на внутренней

Слайд 33Внешняя
среда
Внутренняя
среда
МЕМБРАНА
АТФ
АДФ+Фн
2К
+
3Na
+
2Mg
2+
АТФ-аза
Внешняя
среда
Внутренняя
среда
МЕМБРАНА
АТФ
АДФ+Фн
2К
+
3Na
+
2Mg
2+
АТФ-аза
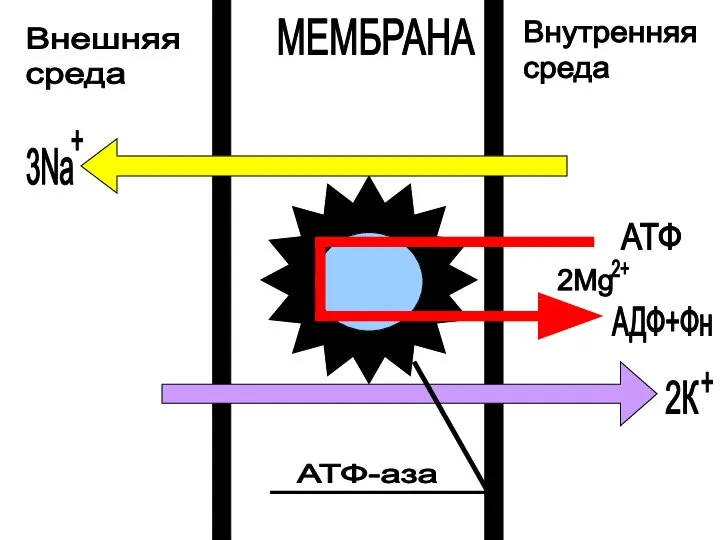
Слайд 34Основные этапы Са2+-насоса :
Первый этап: связывание Са2+ с комплексом Mg-АТФазы на наружной
Основные этапы Са2+-насоса :
Первый этап: связывание Са2+ с комплексом Mg-АТФазы на наружной

Слайд 35КЛЕТКА
МЕМБРАНА
МЕЖКЛЕТОЧНОЕ
ПРОСТРАНСТВО
АТФ
АДФ+Фн
Са
2+
Mg
2+
АТФ-аза
КЛЕТКА
МЕМБРАНА
МЕЖКЛЕТОЧНОЕ
ПРОСТРАНСТВО
АТФ
АДФ+Фн
Са
2+
Mg
2+
АТФ-аза
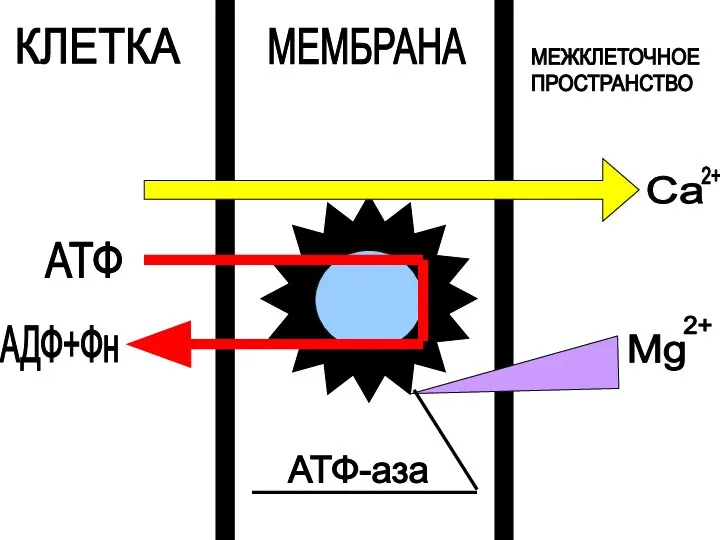
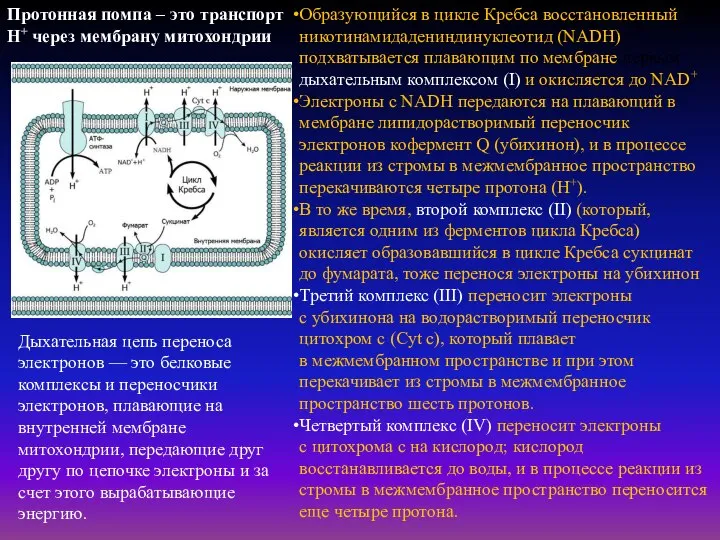
Слайд 36Образующийся в цикле Кребса восстановленный никотинамидадениндинуклеотид (NADH) подхватывается плавающим по мембране первым дыхательным
Образующийся в цикле Кребса восстановленный никотинамидадениндинуклеотид (NADH) подхватывается плавающим по мембране первым дыхательным
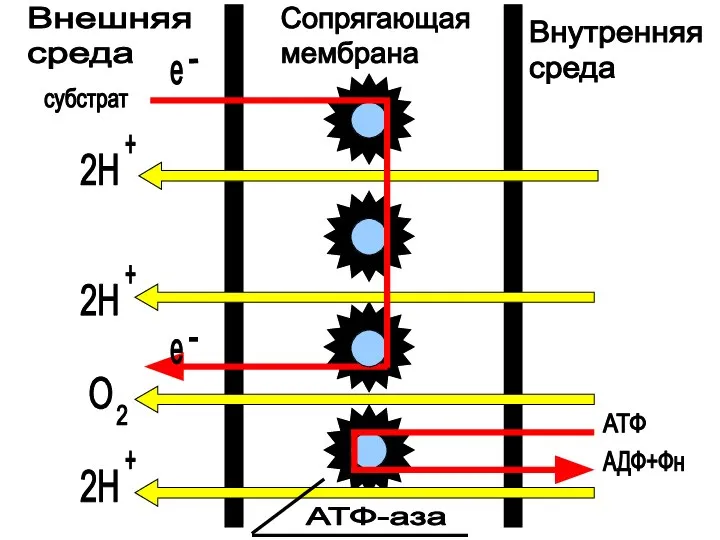
Слайд 37Внешняя
среда
Сопрягающая
мембрана
Внутренняя
среда
субстрат
АТФ
АДФ+Фн
е
е
2Н
2Н
2Н
+
+
+
О
2
АТФ-аза
Внешняя
среда
Сопрягающая
мембрана
Внутренняя
среда
субстрат
АТФ
АДФ+Фн
е
е
2Н
2Н
2Н
+
+
+
О
2
АТФ-аза
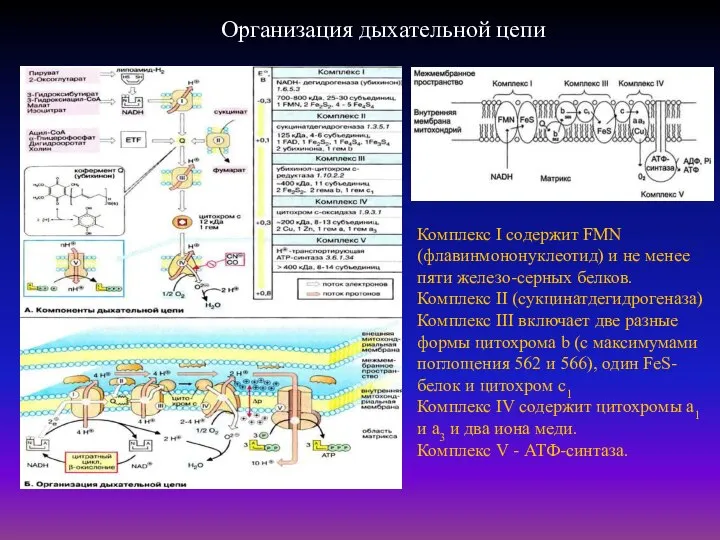
Слайд 38Организация дыхательной цепи
Комплекс I содержит FMN (флавинмононуклеотид) и не менее пяти железо-серных
Организация дыхательной цепи
Комплекс I содержит FMN (флавинмононуклеотид) и не менее пяти железо-серных
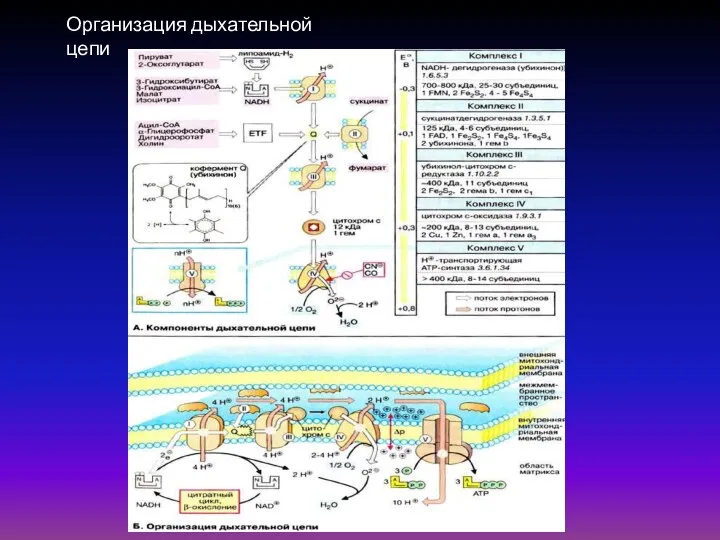
Слайд 39Организация дыхательной цепи
Организация дыхательной цепи
Слайд 40Физические основы преобразования и аккумуляции
энергии в биологических системах
Таблица 1. Компоненты дыхательной
Физические основы преобразования и аккумуляции
энергии в биологических системах
Таблица 1. Компоненты дыхательной

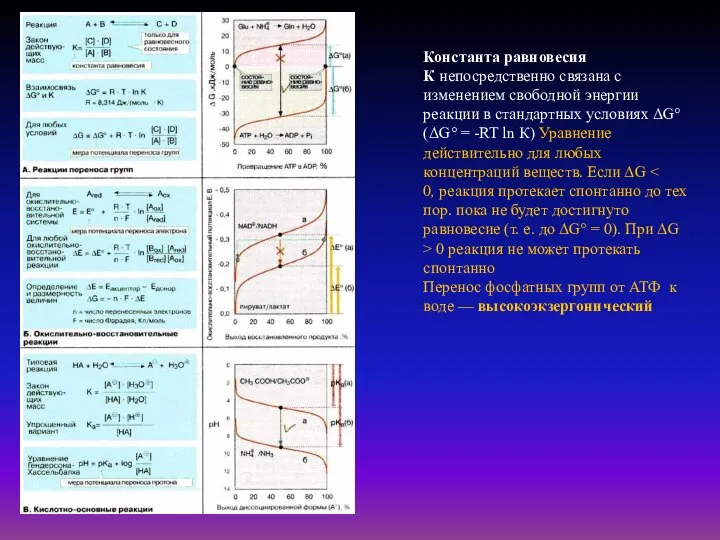
Слайд 41Константа равновесия К непосредственно связана с изменением свободной энергии реакции в стандартных условиях
Константа равновесия К непосредственно связана с изменением свободной энергии реакции в стандартных условиях
Слайд 42Причины высоко экзоэргического характера гидролиза АТФ: сильное отталкивание отрицательно заряженных атомов кислорода, которое
Причины высоко экзоэргического характера гидролиза АТФ: сильное отталкивание отрицательно заряженных атомов кислорода, которое

Слайд 43Механизм синтеза АТФ
АТФ-синтаза состоит из двух частей: встроенного в мембрану протонного канала
Механизм синтеза АТФ
АТФ-синтаза состоит из двух частей: встроенного в мембрану протонного канала

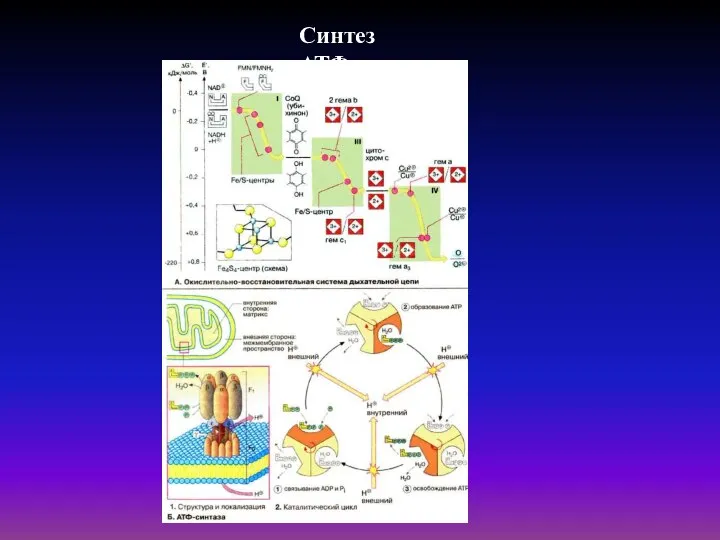
Слайд 44Синтез АТФ
Синтез АТФ
Слайд 45Митохондрии, перенос электронов
Митохондриальная цепь переноса электронов. Комплекс I содержит FMN и не
Митохондрии, перенос электронов
Митохондриальная цепь переноса электронов. Комплекс I содержит FMN и не

Слайд 46Направление процесса и сопряженные реакции
Направление химической реакции определяется значением ΔG. Если эта
Направление процесса и сопряженные реакции
Направление химической реакции определяется значением ΔG. Если эта

Слайд 47Механизмы, обеспечивающие протекание сопряженных реакций.
Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата
Механизмы, обеспечивающие протекание сопряженных реакций.
Реакция фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата

Слайд 48Свободная энергия гидролиза некоторых органических фосфатов
Свободная энергия гидролиза некоторых органических фосфатов
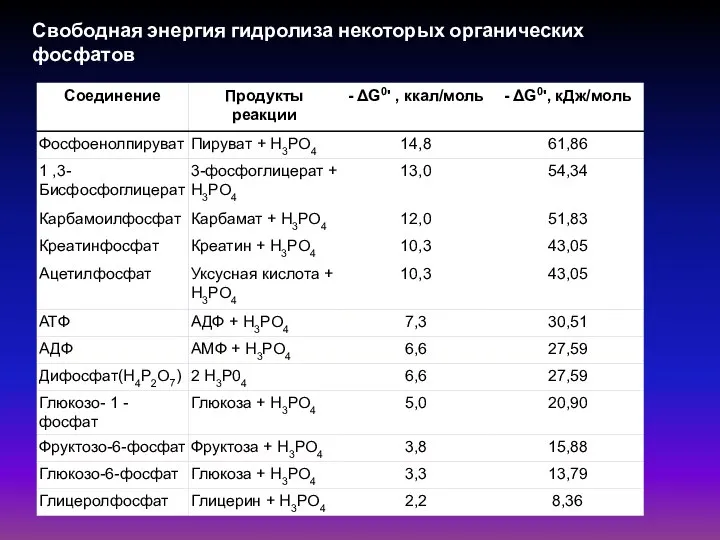
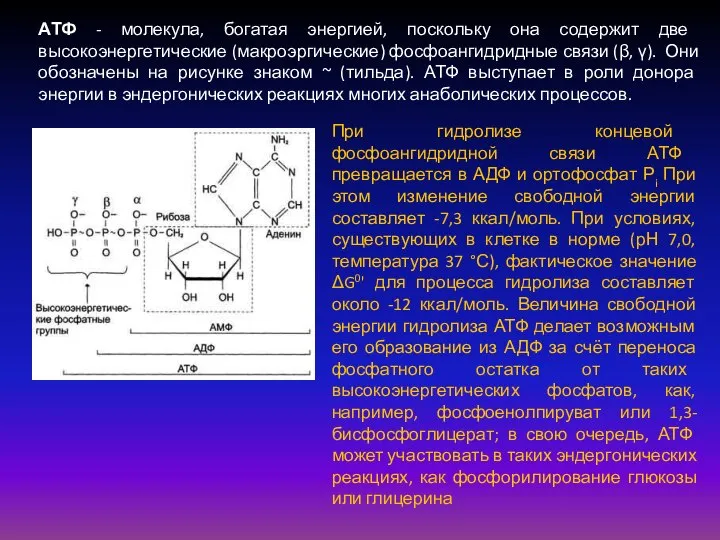
Слайд 49АТФ - молекула, богатая энергией, поскольку она содержит две высокоэнергетические (макроэргические) фосфоангидридные
АТФ - молекула, богатая энергией, поскольку она содержит две высокоэнергетические (макроэргические) фосфоангидридные
Слайд 50Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифосфатов, аналогов
Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифосфатов, аналогов

Слайд 51Биологическое окисление
Под окислением понимают отщепление электронов, а под восстановлением - присоединение электронов.
Биологическое окисление
Под окислением понимают отщепление электронов, а под восстановлением - присоединение электронов.

Слайд 52Стандартные окислительно-восстановительные потенциалы
некоторых сопряжённых пар
Величину Еº' выражают в вольтах; чем
Стандартные окислительно-восстановительные потенциалы
некоторых сопряжённых пар
Величину Еº' выражают в вольтах; чем

 Рептилии. Распространение и виды
Рептилии. Распространение и виды Колорад қоңызы
Колорад қоңызы Биоформы в художественном конструировании
Биоформы в художественном конструировании Естественный отбор
Естественный отбор Крылатые насекомые
Крылатые насекомые Презентация на тему Кровь и кровообращение
Презентация на тему Кровь и кровообращение  Lektsia_2 (1)
Lektsia_2 (1) Анатомия верхней челюсти
Анатомия верхней челюсти Животные Оренбуржья
Животные Оренбуржья Удивительные орхидеи
Удивительные орхидеи Ошибки в биологических олимпиадах
Ошибки в биологических олимпиадах Осенний КВН для учащихся 5 классов
Осенний КВН для учащихся 5 классов Места/среды обитания живых организмов
Места/среды обитания живых организмов Основные положения теории Чарльза Дарвина об эволюции органического мира. 9 класс
Основные положения теории Чарльза Дарвина об эволюции органического мира. 9 класс Микробиологический анализ морской воды платного и муниципального пляжей г. Сочи
Микробиологический анализ морской воды платного и муниципального пляжей г. Сочи Мучнистая роса
Мучнистая роса Биологические понятия и термины
Биологические понятия и термины Обмен и потери витаминов
Обмен и потери витаминов Грунт в аквариуме
Грунт в аквариуме Календарь сезонных овощей и фруктов
Календарь сезонных овощей и фруктов Семь основных таксономических категорий
Семь основных таксономических категорий Увеличение органических веществ почвы. Поддержание жизни на Земле
Увеличение органических веществ почвы. Поддержание жизни на Земле Полезные и вредные насекомые
Полезные и вредные насекомые Презентация на тему Веселая биология
Презентация на тему Веселая биология  Пути утилизации кислорода в организме
Пути утилизации кислорода в организме Антропогенез
Антропогенез Подтип Жгутиковые
Подтип Жгутиковые Мозазавр - род вымерших морских пресмыкающихся
Мозазавр - род вымерших морских пресмыкающихся