- Физиология микроорганизмов

Содержание
- 2. ЛИТЕРАТУРА Основная Готтшалк, Г. Метаболизм бактерий / Г. Готтшалк. М.: Мир, 1982. Гусев, М. В. Микробиология
- 3. ЛИТЕРАТУРА Дополнительная Белясова, Н.А. Микробиология / Н.А. Белясова. Минск: Выш. шк., 2012. Гутина, В.Н. Очерки по
- 4. Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
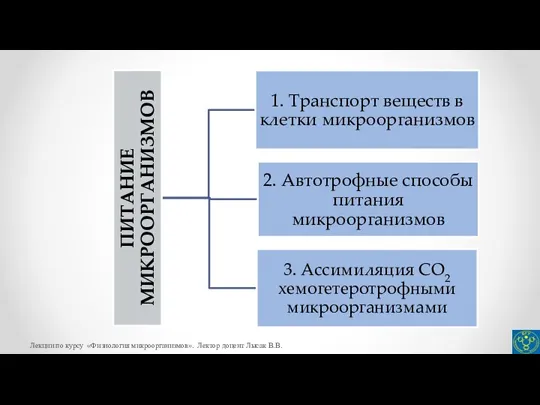
- 5. Питание микроорганизмов Питание микроорганизмов – включение в метаболические реакции любого характера тех или иных соединений внешней
- 6. Питание микроорганизмов К микроэлементам относятся металлы переходной группы (V, Mn, Fe, Co, Ni, Cu, Zn, Mo,
- 7. Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
- 8. Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
- 9. Механизмы, обеспечивающие детоксикацию сильных окислителей у аэробных микроорганизмов Многие клетки синтезируют ферменты каталазу и пероксидазу, защищающие
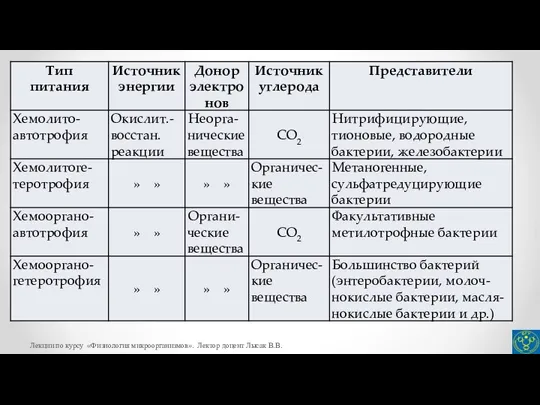
- 10. Источники азота для микроорганизмов Одним из основных элементов, из которых построены клетки микроорганизмов, является азот. Большинство
- 11. Железо как элемент питания Микроорганизмам для осуществления жизнедеятельности требуется также в относительно больших количествах железо. Оно
- 12. Железо как элемент питания Поэтому большинство аэробных микроорганизмов для поглощения железа синтезируют и секретируют в среду
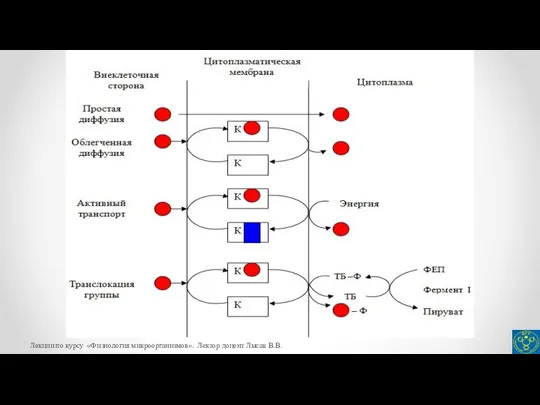
- 13. Транспорт веществ в клетку бактерий Различают следующие способы поступления веществ в клетку бактерий: простая, или пассивная
- 14. Транспорт веществ в клетку бактерий Таким путем осуществляется транcпорт газов, таких как О2, N2, CO2, H2
- 15. Облегченная диффузия (унипорт) Перенос веществ при облегченной диффузии (унипорте) также происходит по градиенту концентрации переносимого вещества.
- 16. Облегченная диффузия (унипорт) Путем облегченной диффузии происходит транспорт в клетку: глюкозы у бактерий Zymomonas mobilis, лизина
- 17. Активный транспорт Активный транспорт – основной механизм избирательного переноса веществ через цитоплазматическую мембрану в клетку против
- 18. Активный транспорт Различают активный транспорт первого и второго класса. Наиболее распространенным механизмом активного поглощения субстратов является
- 19. Активный транспорт Примерами симпорта является перенос протона (Н+) и молекулы лактозы у бактерий E.coli, перенос пролина
- 20. Активный транспорт Хорошо изучены механизмы антипорта субстрата и продукта реакции, например, антипорт малат/лактат при сбраживании малата
- 21. Активный транспорт В системах активного транспорта второго класса перенос растворенного вещества сопряжен с синтезом или гидролизом
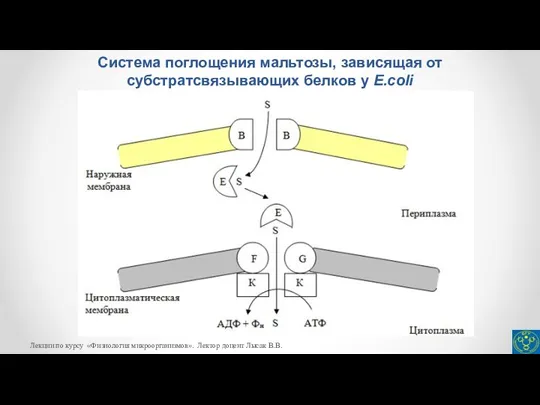
- 22. Активный транспорт Ко второму классу транспортных систем относится также транспорт веществ, зависимый от периплазматических субстратсвязывающих белков.
- 23. Система поглощения мальтозы, зависящая от субстратсвязывающих белков у E.coli Лекции по курсу «Физиология микроорганизмов». Лектор доцент
- 24. К третьему классу активного транспорта относятся специфические транспортные системы, которые осуществляют выделение из клеток ионов натрия,
- 25. При транслокации группы происходит химическая модификация переносимых молекул, тогда как при пассивной диффузии, облегченной диффузии и
- 26. Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
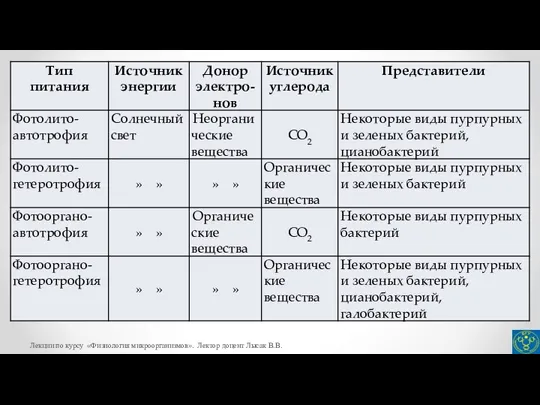
- 27. Автотрофные способы питания микроорганизмов Известно четыре различных механизма фиксации СО2. Наиболее распространен у прокариот цикл Кальвина,
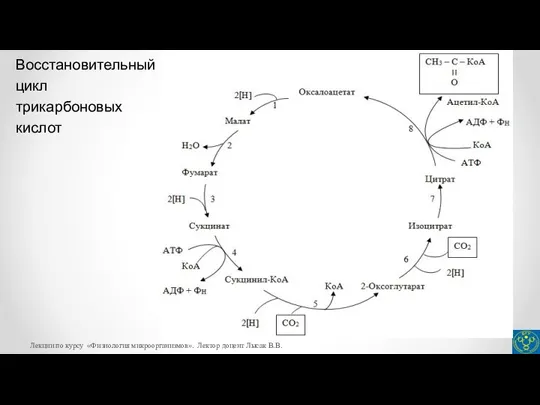
- 28. Восстановительный цикл трикарбоновых кислот Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
- 29. Автотрофные способы питания микроорганизмов На рисунке представлены реакции восстановительного цикла трикарбоновых кислот: 1 – малатдегидрогеназа; 2
- 30. Третий механизм автотрофной фиксации СО2 – восстановительный ацетил-КоА путь осуществляется у некоторых сульфатредуцирующих бактерий (род Desulfobacterium)
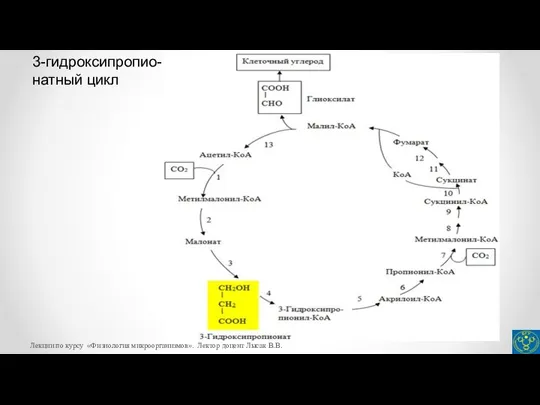
- 31. Глиоксилат относится к метаболитам-предшественникам, используемым в биосинтетических реакциях. На рисунке представлены реакции 3-гидроксипропионатного цикла. 1 –
- 32. 3-гидроксипропио- натный цикл Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
- 33. Ассимиляция СО2 хемогетеротрофами Все гетеротрофные микроорганизмы с помощью определенных ферментативных реакций активно включают диоксид углерода в
- 34. Ассимиляция СО2 хемогетеротрофами В этом случае, СО2, участвующий в реакциях энергетического метаболизма, не включается в вещества
- 35. Реакции включения СО2 в метаболиты клеток Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
- 37. Скачать презентацию

Слайд 3ЛИТЕРАТУРА
Дополнительная
Белясова, Н.А. Микробиология / Н.А. Белясова. Минск: Выш. шк., 2012.
Гутина, В.Н. Очерки
ЛИТЕРАТУРА
Дополнительная
Белясова, Н.А. Микробиология / Н.А. Белясова. Минск: Выш. шк., 2012.
Гутина, В.Н. Очерки

Слайд 4
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Слайд 5Питание микроорганизмов
Питание микроорганизмов – включение в метаболические реакции любого характера тех или
Питание микроорганизмов
Питание микроорганизмов – включение в метаболические реакции любого характера тех или

Слайд 6Питание микроорганизмов
К микроэлементам относятся металлы переходной группы (V, Mn, Fe, Co, Ni,
Питание микроорганизмов
К микроэлементам относятся металлы переходной группы (V, Mn, Fe, Co, Ni,

Слайд 7Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Слайд 8Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Слайд 9Механизмы, обеспечивающие детоксикацию сильных окислителей у аэробных микроорганизмов
Многие клетки синтезируют ферменты каталазу
Механизмы, обеспечивающие детоксикацию сильных окислителей у аэробных микроорганизмов
Многие клетки синтезируют ферменты каталазу

Слайд 10Источники азота для микроорганизмов
Одним из основных элементов, из которых построены клетки микроорганизмов,
Источники азота для микроорганизмов
Одним из основных элементов, из которых построены клетки микроорганизмов,

Слайд 11Железо как элемент питания
Микроорганизмам для осуществления жизнедеятельности требуется также в относительно больших
Железо как элемент питания
Микроорганизмам для осуществления жизнедеятельности требуется также в относительно больших

Слайд 12Железо как элемент питания
Поэтому большинство аэробных микроорганизмов для поглощения железа синтезируют и
Железо как элемент питания
Поэтому большинство аэробных микроорганизмов для поглощения железа синтезируют и

Слайд 13Транспорт веществ в клетку бактерий
Различают следующие способы поступления веществ в клетку бактерий:
Транспорт веществ в клетку бактерий
Различают следующие способы поступления веществ в клетку бактерий:

Слайд 14Транспорт веществ в клетку бактерий
Таким путем осуществляется транcпорт газов, таких как О2,
Транспорт веществ в клетку бактерий
Таким путем осуществляется транcпорт газов, таких как О2,

Слайд 15Облегченная диффузия (унипорт)
Перенос веществ при облегченной диффузии (унипорте) также происходит по градиенту
Облегченная диффузия (унипорт)
Перенос веществ при облегченной диффузии (унипорте) также происходит по градиенту

Слайд 16Облегченная диффузия (унипорт)
Путем облегченной диффузии происходит транспорт в клетку:
глюкозы у бактерий
Облегченная диффузия (унипорт)
Путем облегченной диффузии происходит транспорт в клетку:
глюкозы у бактерий

Слайд 17Активный транспорт
Активный транспорт – основной механизм избирательного переноса веществ через цитоплазматическую мембрану
Активный транспорт
Активный транспорт – основной механизм избирательного переноса веществ через цитоплазматическую мембрану

Слайд 18Активный транспорт
Различают активный транспорт первого и второго класса.
Наиболее распространенным механизмом активного поглощения
Активный транспорт
Различают активный транспорт первого и второго класса.
Наиболее распространенным механизмом активного поглощения

Слайд 19Активный транспорт
Примерами симпорта является перенос протона (Н+) и молекулы лактозы у бактерий
Активный транспорт
Примерами симпорта является перенос протона (Н+) и молекулы лактозы у бактерий

Слайд 20Активный транспорт
Хорошо изучены механизмы антипорта субстрата и продукта реакции, например,
антипорт малат/лактат
Активный транспорт
Хорошо изучены механизмы антипорта субстрата и продукта реакции, например,
антипорт малат/лактат

Слайд 21Активный транспорт
В системах активного транспорта второго класса перенос растворенного вещества сопряжен с
Активный транспорт
В системах активного транспорта второго класса перенос растворенного вещества сопряжен с

Слайд 22Активный транспорт
Ко второму классу транспортных систем относится также транспорт веществ, зависимый от
Активный транспорт
Ко второму классу транспортных систем относится также транспорт веществ, зависимый от

Слайд 23Система поглощения мальтозы, зависящая от субстратсвязывающих белков у E.coli
Лекции по курсу «Физиология
Система поглощения мальтозы, зависящая от субстратсвязывающих белков у E.coli
Лекции по курсу «Физиология
Слайд 24К третьему классу активного транспорта относятся специфические транспортные системы, которые осуществляют выделение
К третьему классу активного транспорта относятся специфические транспортные системы, которые осуществляют выделение

Слайд 25При транслокации группы происходит химическая модификация переносимых молекул, тогда как при пассивной
При транслокации группы происходит химическая модификация переносимых молекул, тогда как при пассивной

Слайд 26Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Слайд 27Автотрофные способы питания микроорганизмов
Известно четыре различных механизма фиксации СО2. Наиболее распространен у
Автотрофные способы питания микроорганизмов
Известно четыре различных механизма фиксации СО2. Наиболее распространен у

Слайд 28Восстановительный
цикл
трикарбоновых
кислот
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Восстановительный
цикл
трикарбоновых
кислот
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Слайд 29Автотрофные способы питания микроорганизмов
На рисунке представлены реакции восстановительного цикла трикарбоновых кислот:
1
Автотрофные способы питания микроорганизмов
На рисунке представлены реакции восстановительного цикла трикарбоновых кислот:
1

Слайд 30Третий механизм автотрофной фиксации СО2 – восстановительный ацетил-КоА путь осуществляется у некоторых
Третий механизм автотрофной фиксации СО2 – восстановительный ацетил-КоА путь осуществляется у некоторых

Слайд 31Глиоксилат относится к метаболитам-предшественникам, используемым в биосинтетических реакциях.
На рисунке представлены реакции 3-гидроксипропионатного
На рисунке представлены реакции 3-гидроксипропионатного

Слайд 323-гидроксипропио-
натный цикл
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
3-гидроксипропио-
натный цикл
Лекции по курсу «Физиология микроорганизмов». Лектор доцент Лысак В.В.
Слайд 33Ассимиляция СО2 хемогетеротрофами
Все гетеротрофные микроорганизмы с помощью определенных ферментативных реакций активно включают
Ассимиляция СО2 хемогетеротрофами
Все гетеротрофные микроорганизмы с помощью определенных ферментативных реакций активно включают

Слайд 34Ассимиляция СО2 хемогетеротрофами
В этом случае, СО2, участвующий в реакциях энергетического метаболизма, не
Ассимиляция СО2 хемогетеротрофами
В этом случае, СО2, участвующий в реакциях энергетического метаболизма, не

Слайд 35Реакции включения СО2 в метаболиты клеток
Лекции по курсу «Физиология микроорганизмов». Лектор доцент
Реакции включения СО2 в метаболиты клеток
Лекции по курсу «Физиология микроорганизмов». Лектор доцент

 Растительный и животный мир Башкортостана
Растительный и животный мир Башкортостана Моллюски в циклах развития паразитов
Моллюски в циклах развития паразитов Строение эукариотических клеток
Строение эукариотических клеток Пищеварение в желудке и 12-перстной кишке
Пищеварение в желудке и 12-перстной кишке Презентация на тему История красной книги
Презентация на тему История красной книги  Причины акселерации
Причины акселерации Рыба язь
Рыба язь Коровы
Коровы Презентация на тему Основные направления эволюции
Презентация на тему Основные направления эволюции  Влияние поллютантов на возникновение мутаций у бактерий
Влияние поллютантов на возникновение мутаций у бактерий Регуляция метаболизма у бактерий
Регуляция метаболизма у бактерий Характер строения и окраска ствола и ветвей
Характер строения и окраска ствола и ветвей Безусловные рефлексы
Безусловные рефлексы Эволюция органического мира
Эволюция органического мира Фармакология процессов обмена веществ и системы крови
Фармакология процессов обмена веществ и системы крови Организм человека, как единая биологическая система
Организм человека, как единая биологическая система Наследственность и изменчивость. Роль наследственности и изменчивости в эволюции
Наследственность и изменчивость. Роль наследственности и изменчивости в эволюции Устройство микроскопа
Устройство микроскопа Строение и функции головного мозга
Строение и функции головного мозга Ландыш майский
Ландыш майский Пуповина (пупочный канатик)
Пуповина (пупочный канатик) Особенности внутреннего строения рыб (часть 2)
Особенности внутреннего строения рыб (часть 2) Современные методы палеонтологических исследований
Современные методы палеонтологических исследований Кожа и её производные
Кожа и её производные Бром и его биологическое значение
Бром и его биологическое значение Что такое метаболизм?
Что такое метаболизм? Познавательные процессы
Познавательные процессы Презентация на тему Железо внутри нас
Презентация на тему Железо внутри нас