- Физиология возбудимых тканей

Содержание
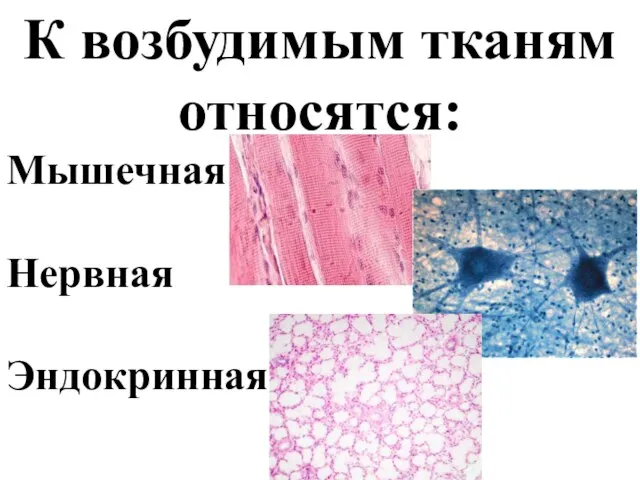
- 2. К возбудимым тканям относятся: Мышечная Нервная Эндокринная
- 3. Общие свойства возбудимых тканей: Раздражимость – способность органов, тканей, клеток активно реагировать на воздействие изменением своих
- 4. Раздражитель – агент внешней или внутренней среды организма, воздействуя на клетки, ткани, органы способен вызвать возбуждение.
- 5. При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии текущего метаболизма. Реакции клетки,
- 6. Возбуждение – реакция возбудимой ткани на действие раздражителя, проявляющаяся в совокупности физических, физико-химических и функциональных изменений.
- 7. Биоэлектрические явления в тканях Луиджи Гальвани (1871 г.) Мембранная теория возникновения мембранных потенциалов (теория натрий-калиевого насоса)
- 9. Мембранный электрогенез
- 10. Мембрана животной клетки состоит из двойного слоя фосфолипидов, трансмембранных и периферических белков, а также поверхностных гликолипидов
- 11. Мембранный электрогенез
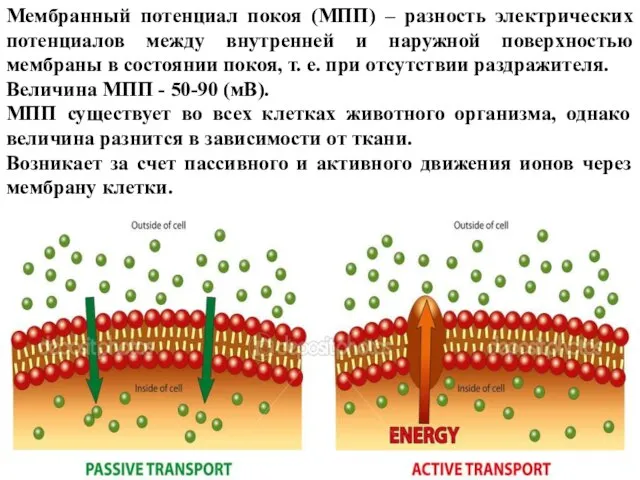
- 12. Мембранный потенциал покоя (МПП) – разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии
- 13. Пассивный транспорт
- 14. Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат энергии. Клеточная мембрана в состоянии
- 16. Ионов натрия в 10 раз больше во внеклеточной жидкости, чем в клетке, проницаемость их через мембрану
- 17. Активный транспорт
- 18. Активный перенос ионов осуществляется в результате деятельности ионного Na+-K+ насоса, представляющего собой фермент плазматической мембраны. Работа
- 19. Все клетки возбудимых тканей при действии различных раздражителей пороговой силы способны переходить в состояние возбуждения. Обязательный
- 20. Мембранный потенциал действия (МПД) – пикообразное колебание потенциала (спайк), возникающее в результате кратковременной перезарядки мембраны и
- 22. Пик потенциала действия связан с процессом возбуждения, он имеет восходящую и нисходящую фазы. Во время восходящей
- 23. При действии раздражителей пороговой или сверхпороговой силы наступает деполяризация клеточной мембраны. Когда деполяризация достигает критической величины,
- 24. Вследствие уменьшения положительных зарядов наружная поверхность мембраны за счет внеклеточных анионов заряжается отрицательно. Но увеличение натриевой
- 25. Вслед за этим в 10-15 раз повышается проницаемость мембраны для ионов калия. Они начинают усиленно выходить
- 26. Реполяризация мембраны бывает замедлена (следовые потенциалы), что обусловлено продолжающимся выходом из клетки ионов калия. Различают деполяризационный
- 27. Основные свойства возбудимых тканей Раздражимость - общее свойство любой живой ткани, клетки реагировать на раздражение изменением
- 28. Кривая «силы-времени» Гоорвега-Вейса
- 29. Изменение возбудимости ткани при возбуждении Абсолютная рефрактерность - состояние, когда ткань после раздражения временно не реагирует
- 31. Проводимость – способность ткани проводить возбуждение (Л. Герман, 1879 г., А. Ходжкин, 1937 г.). Между возбужденным
- 32. Лабильность (функциональная подвижность) - скорость, с которой в ткани возникает и успевает закончиться полный период отдельного
- 33. Усвоение ритма - понижение и повышение физиологической лабильности в связи с деятельностью, проявляющееся более высоким или
- 35. Скачать презентацию
Слайд 3Общие свойства возбудимых тканей:
Раздражимость – способность органов, тканей, клеток активно реагировать на
Общие свойства возбудимых тканей:
Раздражимость – способность органов, тканей, клеток активно реагировать на

Слайд 4Раздражитель – агент внешней или внутренней среды организма, воздействуя на клетки, ткани,
Раздражитель – агент внешней или внутренней среды организма, воздействуя на клетки, ткани,

Слайд 5При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии
При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии

Слайд 6Возбуждение – реакция возбудимой ткани на действие раздражителя, проявляющаяся в совокупности физических,
Возбуждение – реакция возбудимой ткани на действие раздражителя, проявляющаяся в совокупности физических,

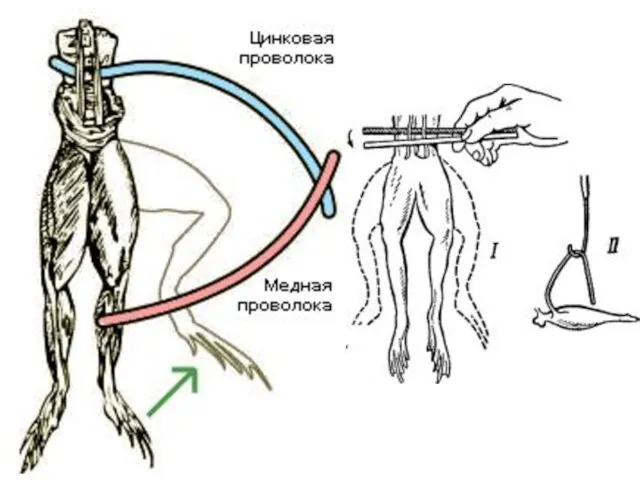
Слайд 7Биоэлектрические явления в тканях
Луиджи Гальвани (1871 г.)
Мембранная теория возникновения мембранных потенциалов (теория
Биоэлектрические явления в тканях
Луиджи Гальвани (1871 г.)
Мембранная теория возникновения мембранных потенциалов (теория

Слайд 9Мембранный электрогенез
Мембранный электрогенез
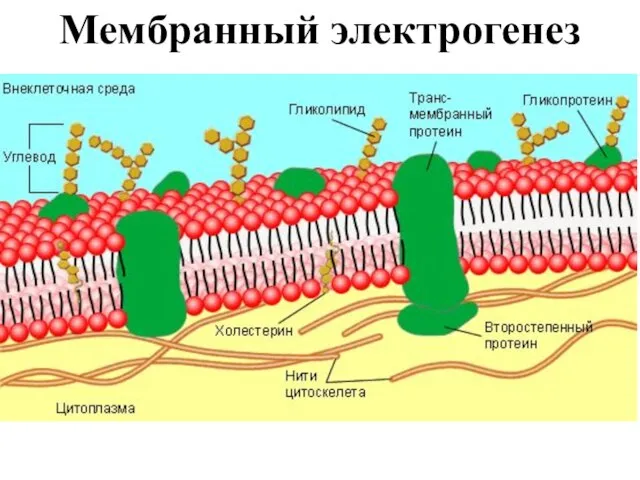
Слайд 10Мембрана животной клетки состоит из двойного слоя фосфолипидов, трансмембранных и периферических белков,
Мембрана животной клетки состоит из двойного слоя фосфолипидов, трансмембранных и периферических белков,

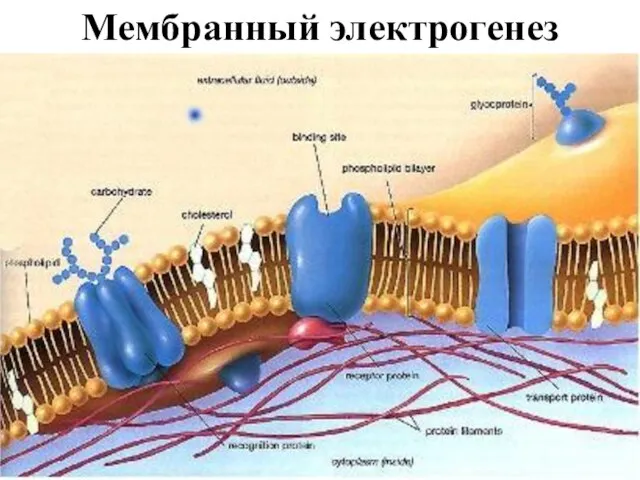
Слайд 11Мембранный электрогенез
Мембранный электрогенез
Слайд 12Мембранный потенциал покоя (МПП) – разность электрических потенциалов между внутренней и наружной
Мембранный потенциал покоя (МПП) – разность электрических потенциалов между внутренней и наружной
Слайд 13Пассивный транспорт
Пассивный транспорт
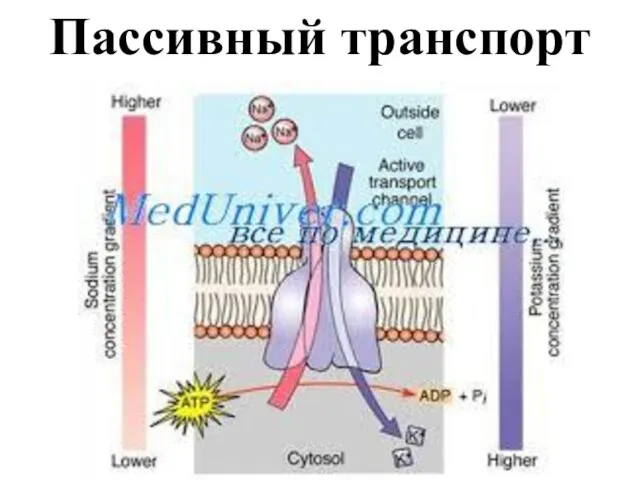
Слайд 14Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат энергии.
Пассивное движение ионов осуществляется по градиенту концентрации и не требует затрат энергии.

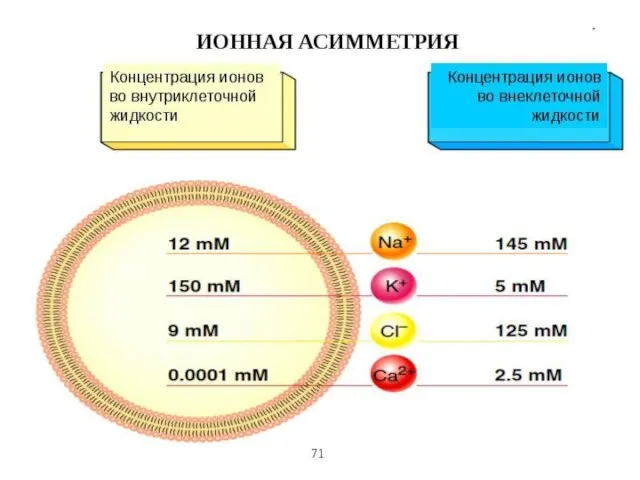
Слайд 16Ионов натрия в 10 раз больше во внеклеточной жидкости, чем в клетке,
Ионов натрия в 10 раз больше во внеклеточной жидкости, чем в клетке,

Слайд 17Активный транспорт
Активный транспорт
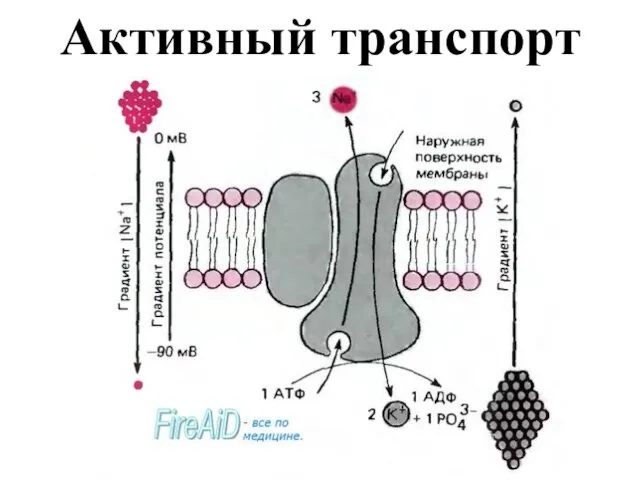
Слайд 18Активный перенос ионов осуществляется в результате деятельности ионного Na+-K+ насоса, представляющего собой
Активный перенос ионов осуществляется в результате деятельности ионного Na+-K+ насоса, представляющего собой

Слайд 19Все клетки возбудимых тканей при действии различных раздражителей пороговой силы способны переходить
Все клетки возбудимых тканей при действии различных раздражителей пороговой силы способны переходить

Слайд 20Мембранный потенциал действия (МПД) – пикообразное колебание потенциала (спайк), возникающее в результате
Мембранный потенциал действия (МПД) – пикообразное колебание потенциала (спайк), возникающее в результате

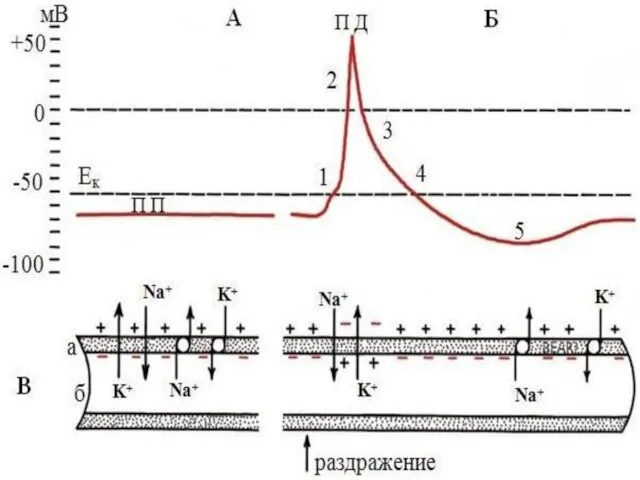
Слайд 22Пик потенциала действия связан с процессом возбуждения, он имеет восходящую и нисходящую
Пик потенциала действия связан с процессом возбуждения, он имеет восходящую и нисходящую

Слайд 23При действии раздражителей пороговой или сверхпороговой силы наступает деполяризация клеточной мембраны. Когда
При действии раздражителей пороговой или сверхпороговой силы наступает деполяризация клеточной мембраны. Когда

Слайд 24Вследствие уменьшения положительных зарядов наружная поверхность мембраны за счет внеклеточных анионов заряжается
Вследствие уменьшения положительных зарядов наружная поверхность мембраны за счет внеклеточных анионов заряжается
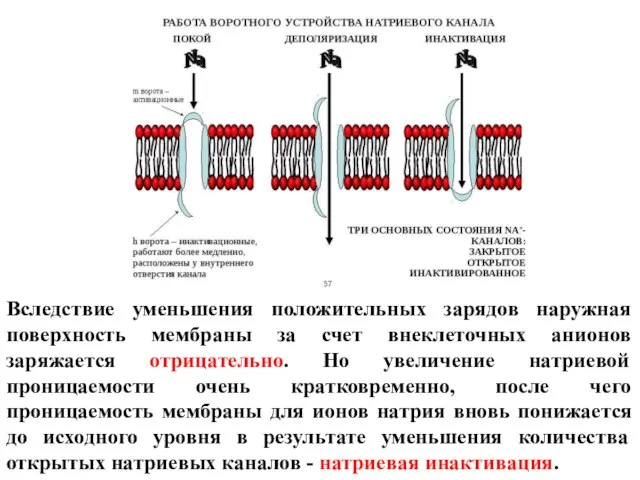
Слайд 25Вслед за этим в 10-15 раз повышается проницаемость мембраны для ионов калия.
Вслед за этим в 10-15 раз повышается проницаемость мембраны для ионов калия.
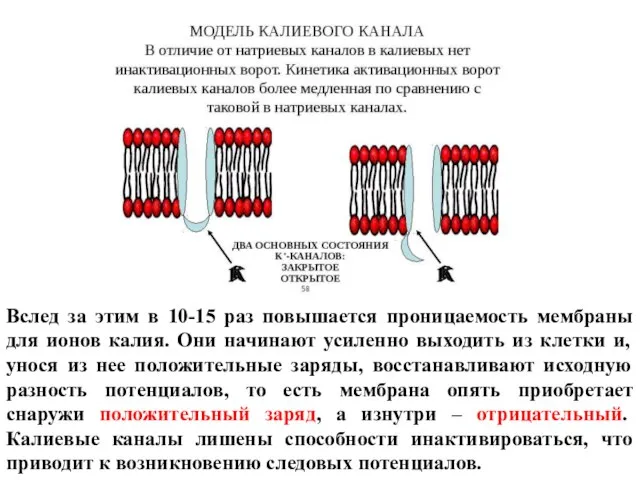
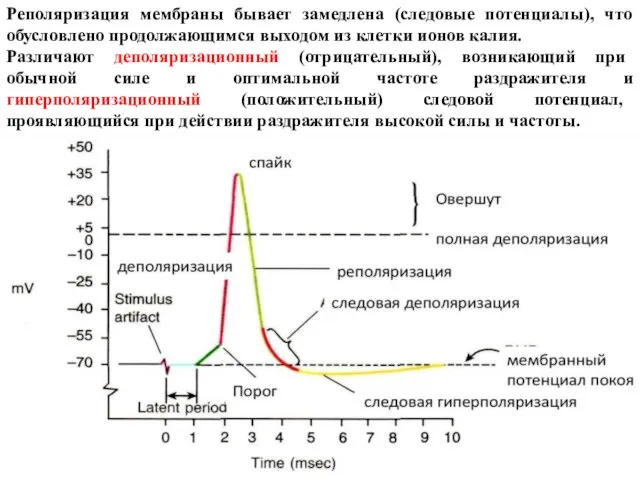
Слайд 26Реполяризация мембраны бывает замедлена (следовые потенциалы), что обусловлено продолжающимся выходом из клетки
Реполяризация мембраны бывает замедлена (следовые потенциалы), что обусловлено продолжающимся выходом из клетки
Слайд 27Основные свойства возбудимых тканей
Раздражимость - общее свойство любой живой ткани, клетки реагировать
Основные свойства возбудимых тканей
Раздражимость - общее свойство любой живой ткани, клетки реагировать

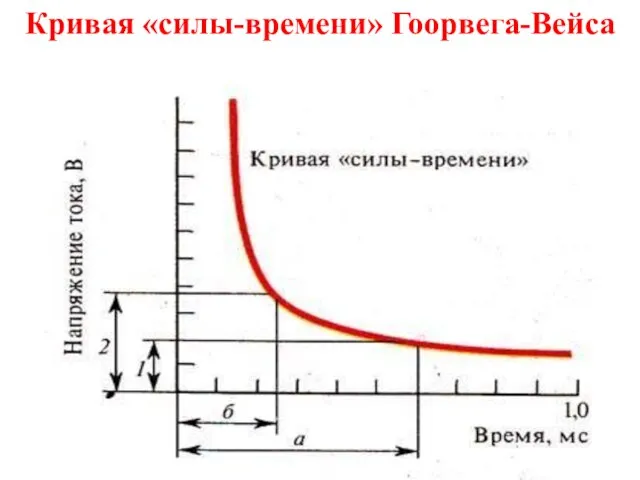
Слайд 28Кривая «силы-времени» Гоорвега-Вейса
Кривая «силы-времени» Гоорвега-Вейса
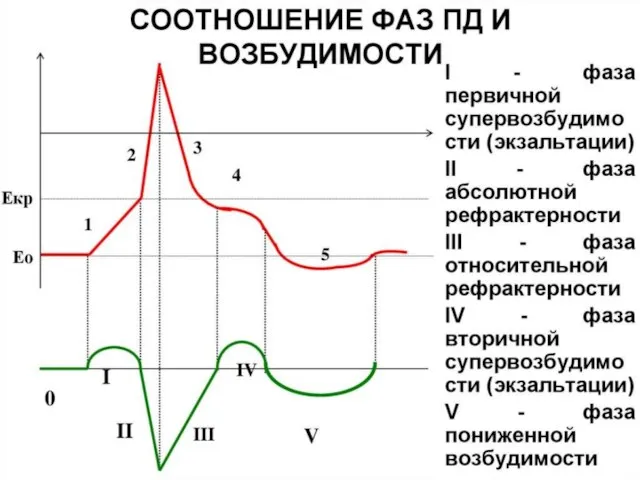
Слайд 29Изменение возбудимости ткани при возбуждении
Абсолютная рефрактерность - состояние, когда ткань после раздражения
Изменение возбудимости ткани при возбуждении
Абсолютная рефрактерность - состояние, когда ткань после раздражения

Слайд 31Проводимость – способность ткани проводить возбуждение (Л. Герман, 1879 г., А. Ходжкин,
Проводимость – способность ткани проводить возбуждение (Л. Герман, 1879 г., А. Ходжкин,

Слайд 32Лабильность (функциональная подвижность) - скорость, с которой в ткани возникает и успевает
Лабильность (функциональная подвижность) - скорость, с которой в ткани возникает и успевает

Слайд 33Усвоение ритма - понижение и повышение физиологической лабильности в связи с деятельностью,
Усвоение ритма - понижение и повышение физиологической лабильности в связи с деятельностью,

 Презентация на тему Бактерии помогают или вредят человеку
Презентация на тему Бактерии помогают или вредят человеку  Поиск оптимальных условий непосредственно в процессе выращивания микроорганизмов
Поиск оптимальных условий непосредственно в процессе выращивания микроорганизмов Белки. Протеиногенные аминокислоты
Белки. Протеиногенные аминокислоты Строение органа слуха
Строение органа слуха Тип Плоские черви
Тип Плоские черви Тип Моллюски. 7 класс
Тип Моллюски. 7 класс Бионеорганическая химия. Комплексообразование в организме. (Лекция 3)
Бионеорганическая химия. Комплексообразование в организме. (Лекция 3) Дигибридное скрещивание
Дигибридное скрещивание Fox lives in the forest
Fox lives in the forest Зимующие птицы городов
Зимующие птицы городов Наружные покровы человека
Наружные покровы человека Вегетативное размножение (стеблевое)
Вегетативное размножение (стеблевое) Заяц-беляк. Интересные факты о зайцах
Заяц-беляк. Интересные факты о зайцах Значение живых организмов в природе
Значение живых организмов в природе Размножение цветковых растений
Размножение цветковых растений embiol_1.Spermatogenesis
embiol_1.Spermatogenesis Презентация на тему История генетики
Презентация на тему История генетики  Физиология гиппокампа
Физиология гиппокампа Периферична нервова система. Соматична й вегетативна нервова система
Периферична нервова система. Соматична й вегетативна нервова система Покровы тела животных. 7 класс
Покровы тела животных. 7 класс Мозг млекопитающих
Мозг млекопитающих Основы зоогигиены
Основы зоогигиены Механизмы проведения возбуждения
Механизмы проведения возбуждения Они рядом с нами – редкие и исчезающие животные
Они рядом с нами – редкие и исчезающие животные Строение и функции половой системы человека
Строение и функции половой системы человека Биомеханическая характеристика ходьбы и бега
Биомеханическая характеристика ходьбы и бега Сравнительно анатомические доказательства инволюции
Сравнительно анатомические доказательства инволюции Кое-что о приспособлениях. Рыбы. Защита. 5
Кое-что о приспособлениях. Рыбы. Защита. 5