- Фотосинтез. Лекция №9

Содержание
- 2. Вопросы с прошлой лекции Почему QH- так обозначен? Почему не 36–38 АТФ образуется всего из глюкозы?
- 3. Почему QH- так обозначен?
- 4. H+ нужен не только АТФ-синтазе АТФ-синтаза АТФ/АДФ-транслоказа фосфат-транслоказа Для синтеза 1 АТФ необходим еще 1 H+,
- 5. Итоги гликолиза, ПДК, ЦТК и ЦПЭ + 2 NAD+ + 2 АДФ + 2 HPO42- 2
- 6. Путь NADH из цитозоля в митохондрии малат-аспартатный челнок глицерол-3фосфатный челнок NADH FADH2
- 7. Итоги гликолиза, ПДК, ЦТК и ЦПЭ + 2 NAD+ + 2 АДФ + 2 HPO42- 2
- 8. Комплексы цепи переноса электронов Комплекс I NADH-дегидрогеназный Комплекс III цитохром с редуктазный Комплекс II сукцинат дегидрогеназный
- 9. https://www.kegg.jp/kegg-bin/highlight_pathway?scale=1.0&map=hsa00190&keyword=Mitochondria
- 10. План лекции Зачем мне фотосинтез, я же буду … Процесс перехода хлорофилла в возбужденное состояние Структура
- 11. Зачем мне нужен фотосинтез, я же буду… https://academic.oup.com/jxb/article/71/7/2219/5736454
- 12. Общая схема фотосинтеза (световые + темновые реакции) световые реакции темновые реакции H2O O2 NADPH ATP CO2
- 13. Переход молекулы хлорофилла в возбужденное состояние хлорофилл а Наличие цепочек чередующихся двойных и одинарных связей делает
- 14. Структура различных пигментов Lehninger Principles of Biochemistry (4th Ed.)
- 15. Спектры поглощения пигментами видимого света длинна волны, нм поглощение Молярный коэффициент экстинкции (ε) хлорофилла а ~
- 16. Строение хлоропластов свет антенные хлорофиллы каротионоиды и другие доп. пигменты Реакционный центр Конвертирует энергию фотона в
- 17. Превращение энергии света в поток электронов
- 18. Что собой представляет реакционный центр? Реакционный центр – это комплекс белков, включающий в себя: Акцептор электрона
- 19. Супрамолекулярный комплекс фотосистемы I с антенными пигментами (зеленые и желтые) и белками (серые) Пигменты в комплексе
- 20. Общая схема световых реакций У сосудистых растений, водорослей и цианобактерий по две фотосистемы (у бактерий –
- 21. Общая схема световых реакций Поток электронов в ФС2 доходит до комплекса, в котором в процессе перемещения
- 22. 1. Возбуждение P680 и транспорт электрона в ФС2 P680* за ~3 пс передаёт электрон на близко
- 23. 1. Возбуждение P680 и транспорт электрона в ФС2 Когда PQB дождался второго электрона, он присоединяет два
- 24. 3. Перенос электрона с пластохинола в комплексе цитохрома b6f Комплекс цитохрома b6f аналогичен комплексу III цепи
- 25. 2. Разложение воды и восстановление P680+ P680, отдавший электрон феофетину, должен быть восстановлен Первичным донором электрона
- 26. Общая схема световых реакций
- 27. 4. Возбуждение P700 и транспорт электрона в ФС1 После цепочки антенных пигментов возбужденный экситоном P700*(два хлорофилла)
- 28. Суммарное уравнение световых реакций 2 H2O + 4 H+N → O2 + 4 H+ + 2
- 29. Темновые реакции фотосинтеза (Реакции, не требующие света) Цикл Кальвина (восстановительный пентозофосфатный цикл) Фотодыхание C4-фотосинтез CAM-фотосинтез
- 30. Цикл Кальвина Этап 1: фиксация Этап 3: регенерация акцептора Этап 2: восстановление Получение энергии в гликолизе;
- 31. Цикл Кальвина: 1. Фиксация CO2 Осуществляется ферментом рибулозо-1,5-бисфосфат карбоксилазой/оксигеназой (рубиско) Рубиско активируется путём карбамоилирования Lys201 с
- 32. Цикл Кальвина: 1. Фиксация CO2 Рубиско – очень медленный фермент (3 CO2/секунду), поэтому данный фермент составляет
- 33. Цикл Кальвина: 2. Восстановление 3-фосфоглицерата до глицеральдегид-3-фосфата
- 34. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата глицеральдегид-3-фосфат дигидроксиацетон-фосфат трансальдолаза фруктозо-1,6-бисфосфат фруктозо-1,6-бисфосфатаза фруктозо-6-фосфат Pi 1 2
- 35. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата глицеральдегид-3-фосфат фруктозо-6-фосфат транскетолаза ксилулозо-5-фосфат эритрозо-4-фосфат 3
- 36. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата эритрозо-4-фосфат дигидроксиацетон-фосфат трансальдолаза седогептулозо-1,7-бисфосфат седогептулозо-7-фосфат седогептулозо-1,7-бисфосфатаза 4 Pi
- 37. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата седогептулозо-7-фосфат глицеральдегид-3-фосфат транскетолаза рибозо-5-фосфат ксилулозо-5-фосфат 5
- 38. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата рибозо-5-фосфат рибозо-5-фосфат изомераза рибулозо-5-фосфат рибулозо-5-фосфат изомераза рибулозо-5-фосфат изомераза рибулозо-5-фосфат рибулозо-5-фосфат
- 39. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата рибулозо-5-фосфат 3× рибулозо-5-фосфат киназа рибулозо-1,5-бисфосфат ATP
- 40. Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата и суммарное уравнение 5 глицеральдегид-3-фосфат + 3 ATP → 3 рибулозо-1,5-бисфосфат
- 41. Фотодыхание O2 конкурирует с CO2 за активный центр рубиско. В 1 из 3 или 4 случаев
- 42. Метаболизм 2-фосфогликолята 2-фосфогликолят гликолят фосфатаза гликолят-оксидаза O2 глиоксилат глицин +глицин + NAD+ + H2O NADH +
- 43. C4-фотосинтез C4-растения живут при высоких температурах, где отношение концентраций растворенных O2 к CO2 еще выше У
- 45. Скачать презентацию

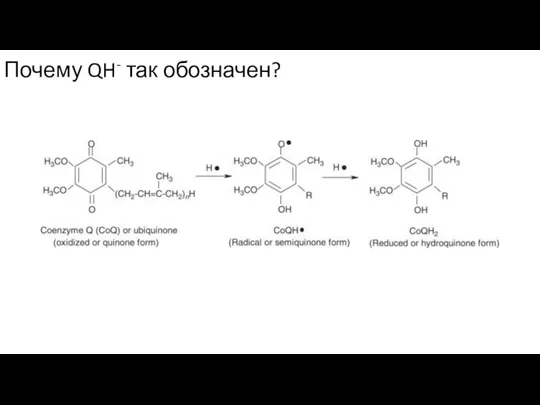
Слайд 3Почему QH- так обозначен?
Почему QH- так обозначен?
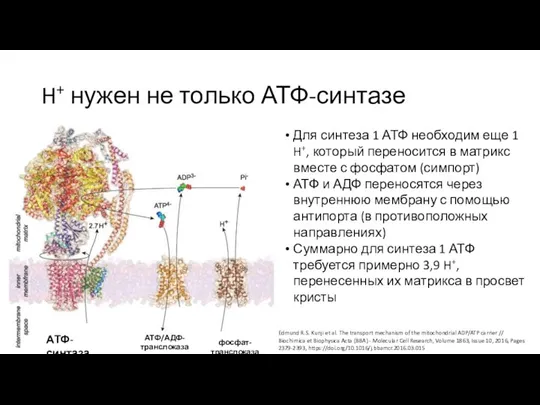
Слайд 4H+ нужен не только АТФ-синтазе
АТФ-синтаза
АТФ/АДФ-транслоказа
фосфат-транслоказа
Для синтеза 1 АТФ необходим еще 1 H+,
H+ нужен не только АТФ-синтазе
АТФ-синтаза
АТФ/АДФ-транслоказа
фосфат-транслоказа
Для синтеза 1 АТФ необходим еще 1 H+,
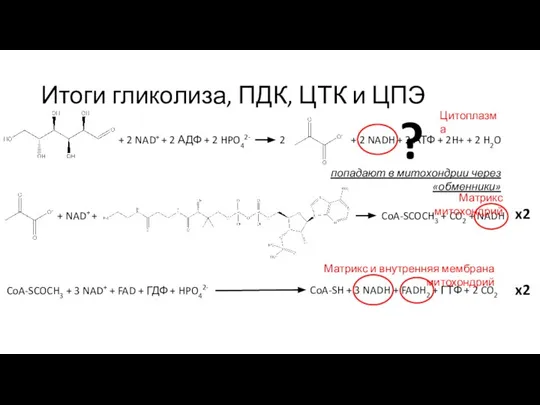
Слайд 5Итоги гликолиза, ПДК, ЦТК и ЦПЭ
+ 2 NAD+ + 2 АДФ +
Итоги гликолиза, ПДК, ЦТК и ЦПЭ
+ 2 NAD+ + 2 АДФ +
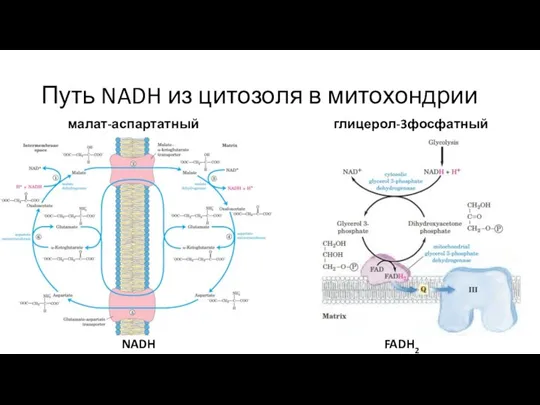
Слайд 6Путь NADH из цитозоля в митохондрии
малат-аспартатный челнок
глицерол-3фосфатный челнок
NADH
FADH2
Путь NADH из цитозоля в митохондрии
малат-аспартатный челнок
глицерол-3фосфатный челнок
NADH
FADH2
Слайд 7Итоги гликолиза, ПДК, ЦТК и ЦПЭ
+ 2 NAD+ + 2 АДФ +
Итоги гликолиза, ПДК, ЦТК и ЦПЭ
+ 2 NAD+ + 2 АДФ +
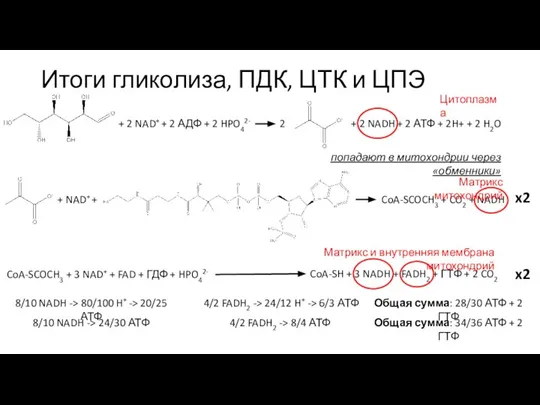
Слайд 8Комплексы цепи переноса электронов
Комплекс I
NADH-дегидрогеназный
Комплекс III
цитохром с редуктазный
Комплекс II
сукцинат дегидрогеназный
Комплекс IV
цитохром с
Комплексы цепи переноса электронов
Комплекс I
NADH-дегидрогеназный
Комплекс III
цитохром с редуктазный
Комплекс II
сукцинат дегидрогеназный
Комплекс IV
цитохром с
Слайд 9https://www.kegg.jp/kegg-bin/highlight_pathway?scale=1.0&map=hsa00190&keyword=Mitochondria
https://www.kegg.jp/kegg-bin/highlight_pathway?scale=1.0&map=hsa00190&keyword=Mitochondria

Слайд 10План лекции
Зачем мне фотосинтез, я же буду …
Процесс перехода хлорофилла в возбужденное
План лекции
Зачем мне фотосинтез, я же буду …
Процесс перехода хлорофилла в возбужденное

Слайд 11Зачем мне нужен фотосинтез, я же буду…
https://academic.oup.com/jxb/article/71/7/2219/5736454
Зачем мне нужен фотосинтез, я же буду…
https://academic.oup.com/jxb/article/71/7/2219/5736454
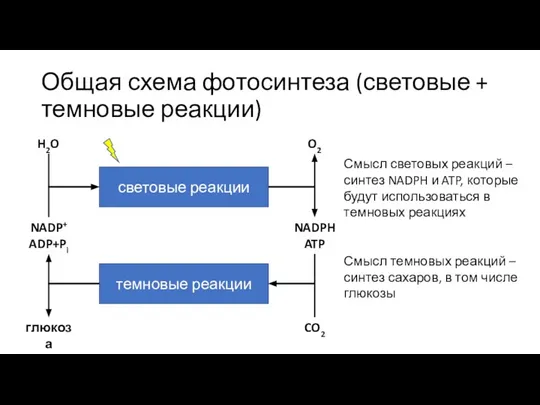
Слайд 12Общая схема фотосинтеза (световые + темновые реакции)
световые реакции
темновые реакции
H2O
O2
NADPH
ATP
CO2
NADP+
ADP+Pi
глюкоза
Смысл световых реакций –
Общая схема фотосинтеза (световые + темновые реакции)
световые реакции
темновые реакции
H2O
O2
NADPH
ATP
CO2
NADP+
ADP+Pi
глюкоза
Смысл световых реакций –
Слайд 13Переход молекулы хлорофилла в возбужденное состояние
хлорофилл а
Наличие цепочек чередующихся двойных и одинарных
Переход молекулы хлорофилла в возбужденное состояние
хлорофилл а
Наличие цепочек чередующихся двойных и одинарных

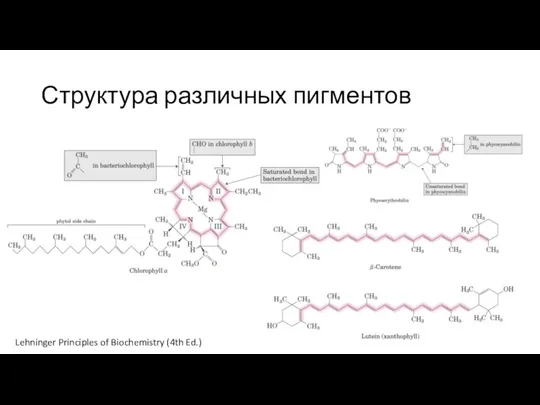
Слайд 14Структура различных пигментов
Lehninger Principles of Biochemistry (4th Ed.)
Структура различных пигментов
Lehninger Principles of Biochemistry (4th Ed.)
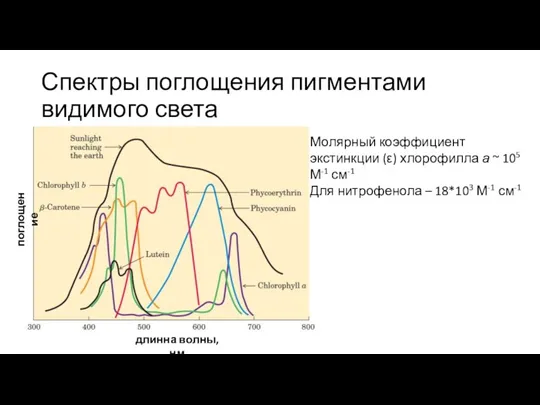
Слайд 15Спектры поглощения пигментами видимого света
длинна волны, нм
поглощение
Молярный коэффициент экстинкции (ε) хлорофилла а
Спектры поглощения пигментами видимого света
длинна волны, нм
поглощение
Молярный коэффициент экстинкции (ε) хлорофилла а
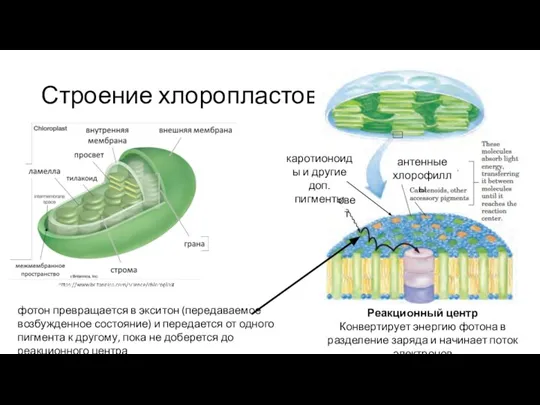
Слайд 16Строение хлоропластов
свет
антенные хлорофиллы
каротионоиды и другие доп. пигменты
Реакционный центр
Конвертирует энергию фотона в разделение
Строение хлоропластов
свет
антенные хлорофиллы
каротионоиды и другие доп. пигменты
Реакционный центр
Конвертирует энергию фотона в разделение
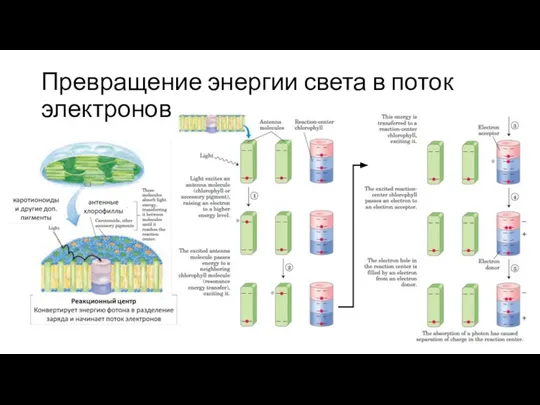
Слайд 17Превращение энергии света в поток электронов
Превращение энергии света в поток электронов
Слайд 18Что собой представляет реакционный центр?
Реакционный центр – это комплекс белков, включающий в
Что собой представляет реакционный центр?
Реакционный центр – это комплекс белков, включающий в

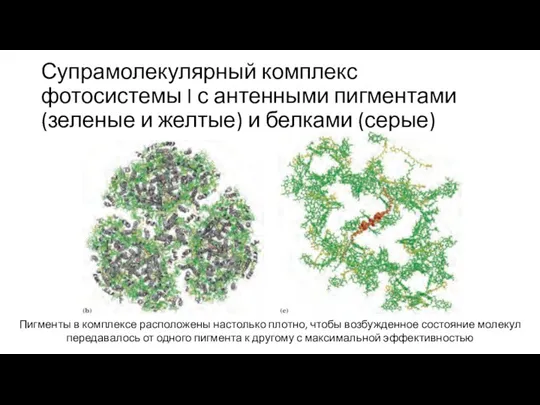
Слайд 19Супрамолекулярный комплекс фотосистемы I с антенными пигментами (зеленые и желтые) и белками
Супрамолекулярный комплекс фотосистемы I с антенными пигментами (зеленые и желтые) и белками
Слайд 20Общая схема световых реакций
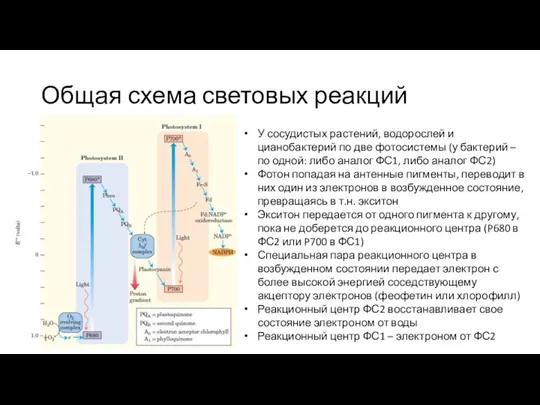
У сосудистых растений, водорослей и цианобактерий по две фотосистемы
Общая схема световых реакций
У сосудистых растений, водорослей и цианобактерий по две фотосистемы
Слайд 21Общая схема световых реакций
Поток электронов в ФС2 доходит до комплекса, в котором
Общая схема световых реакций
Поток электронов в ФС2 доходит до комплекса, в котором

Слайд 221. Возбуждение P680 и транспорт электрона в ФС2
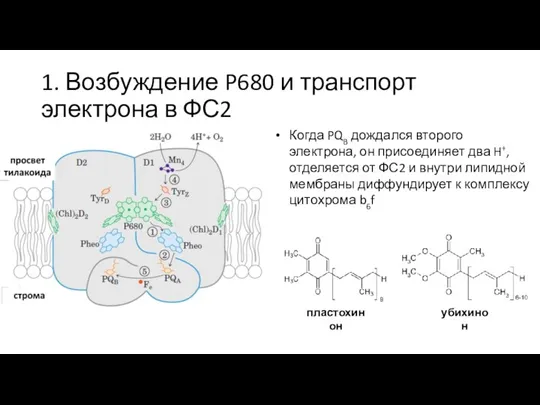
P680* за ~3 пс передаёт
1. Возбуждение P680 и транспорт электрона в ФС2
P680* за ~3 пс передаёт

Слайд 231. Возбуждение P680 и транспорт электрона в ФС2
Когда PQB дождался второго электрона,
1. Возбуждение P680 и транспорт электрона в ФС2
Когда PQB дождался второго электрона,
Слайд 243. Перенос электрона с пластохинола в комплексе цитохрома b6f
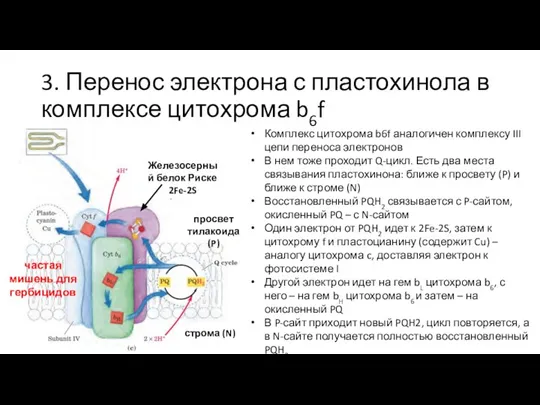
Комплекс цитохрома b6f аналогичен
3. Перенос электрона с пластохинола в комплексе цитохрома b6f
Комплекс цитохрома b6f аналогичен
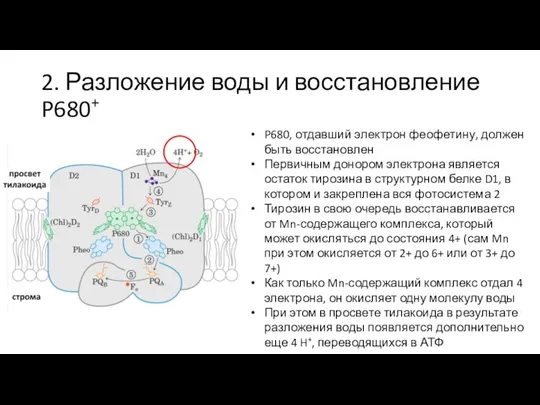
Слайд 252. Разложение воды и восстановление P680+
P680, отдавший электрон феофетину, должен быть восстановлен
Первичным
2. Разложение воды и восстановление P680+
P680, отдавший электрон феофетину, должен быть восстановлен
Первичным
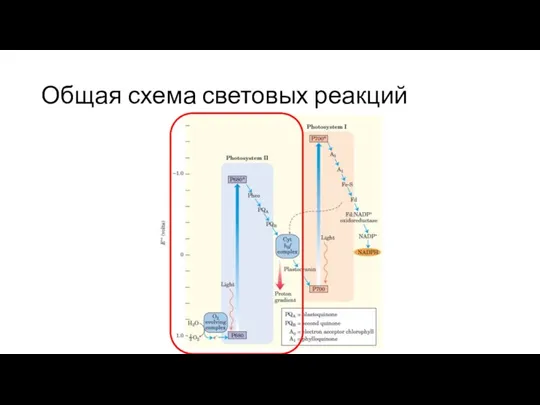
Слайд 26Общая схема световых реакций
Общая схема световых реакций
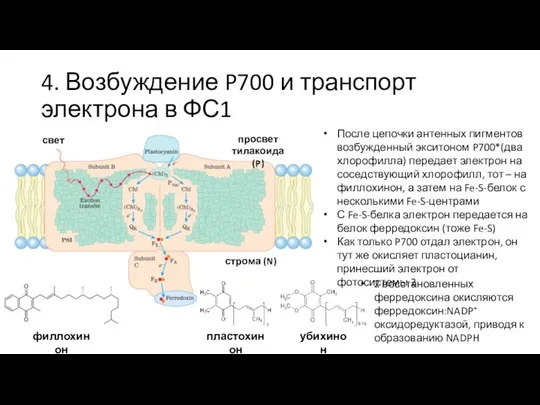
Слайд 274. Возбуждение P700 и транспорт электрона в ФС1
После цепочки антенных пигментов возбужденный
4. Возбуждение P700 и транспорт электрона в ФС1
После цепочки антенных пигментов возбужденный
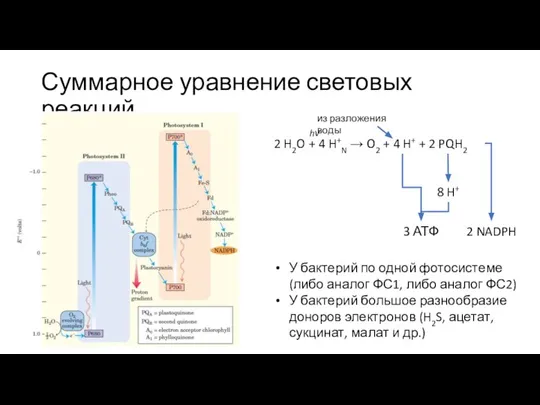
Слайд 28Суммарное уравнение световых реакций
2 H2O + 4 H+N → O2 + 4
Суммарное уравнение световых реакций
2 H2O + 4 H+N → O2 + 4
Слайд 29Темновые реакции фотосинтеза
(Реакции, не требующие света)
Цикл Кальвина (восстановительный пентозофосфатный цикл)
Фотодыхание
C4-фотосинтез
CAM-фотосинтез
Темновые реакции фотосинтеза
(Реакции, не требующие света)
Цикл Кальвина (восстановительный пентозофосфатный цикл)
Фотодыхание
C4-фотосинтез
CAM-фотосинтез

Слайд 30Цикл Кальвина
Этап 1:
фиксация
Этап 3:
регенерация акцептора
Этап 2:
восстановление
Получение энергии в гликолизе; синтез крахмала и
Цикл Кальвина
Этап 1:
фиксация
Этап 3:
регенерация акцептора
Этап 2:
восстановление
Получение энергии в гликолизе; синтез крахмала и
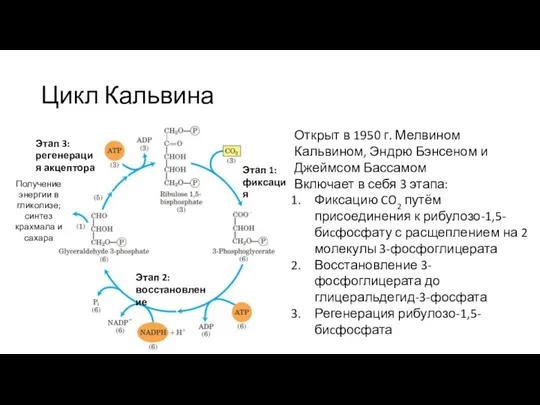
Слайд 31Цикл Кальвина: 1. Фиксация CO2
Осуществляется ферментом рибулозо-1,5-бисфосфат карбоксилазой/оксигеназой (рубиско)
Рубиско активируется путём карбамоилирования
Цикл Кальвина: 1. Фиксация CO2
Осуществляется ферментом рибулозо-1,5-бисфосфат карбоксилазой/оксигеназой (рубиско)
Рубиско активируется путём карбамоилирования
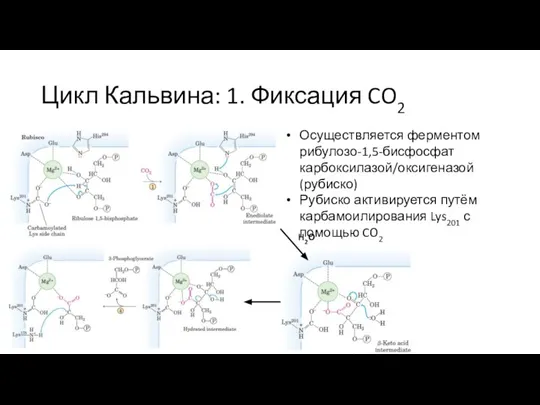
Слайд 32Цикл Кальвина: 1. Фиксация CO2
Рубиско – очень медленный фермент (3 CO2/секунду), поэтому
Цикл Кальвина: 1. Фиксация CO2
Рубиско – очень медленный фермент (3 CO2/секунду), поэтому

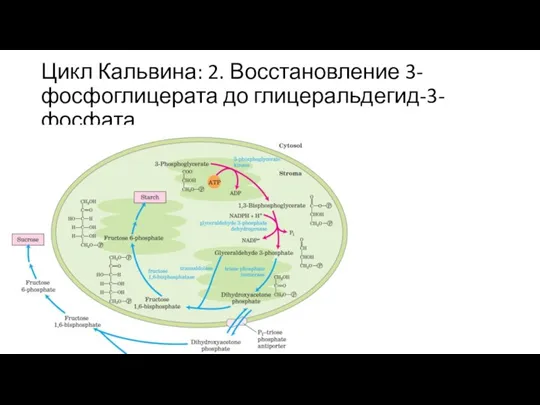
Слайд 33Цикл Кальвина: 2. Восстановление 3-фосфоглицерата до глицеральдегид-3-фосфата
Цикл Кальвина: 2. Восстановление 3-фосфоглицерата до глицеральдегид-3-фосфата
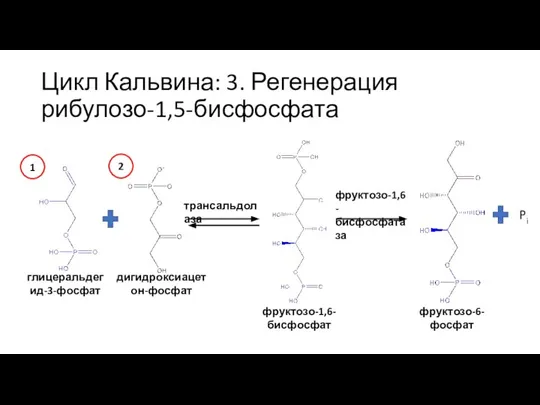
Слайд 34Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
глицеральдегид-3-фосфат
дигидроксиацетон-фосфат
трансальдолаза
фруктозо-1,6-бисфосфат
фруктозо-1,6-бисфосфатаза
фруктозо-6-фосфат
Pi
1
2
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
глицеральдегид-3-фосфат
дигидроксиацетон-фосфат
трансальдолаза
фруктозо-1,6-бисфосфат
фруктозо-1,6-бисфосфатаза
фруктозо-6-фосфат
Pi
1
2
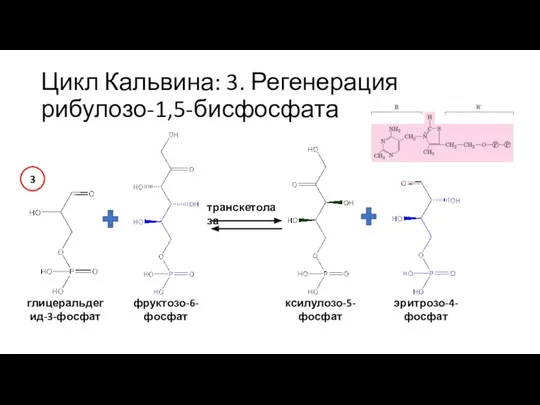
Слайд 35Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
глицеральдегид-3-фосфат
фруктозо-6-фосфат
транскетолаза
ксилулозо-5-фосфат
эритрозо-4-фосфат
3
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
глицеральдегид-3-фосфат
фруктозо-6-фосфат
транскетолаза
ксилулозо-5-фосфат
эритрозо-4-фосфат
3
Слайд 36Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
эритрозо-4-фосфат
дигидроксиацетон-фосфат
трансальдолаза
седогептулозо-1,7-бисфосфат
седогептулозо-7-фосфат
седогептулозо-1,7-бисфосфатаза
4
Pi
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
эритрозо-4-фосфат
дигидроксиацетон-фосфат
трансальдолаза
седогептулозо-1,7-бисфосфат
седогептулозо-7-фосфат
седогептулозо-1,7-бисфосфатаза
4
Pi
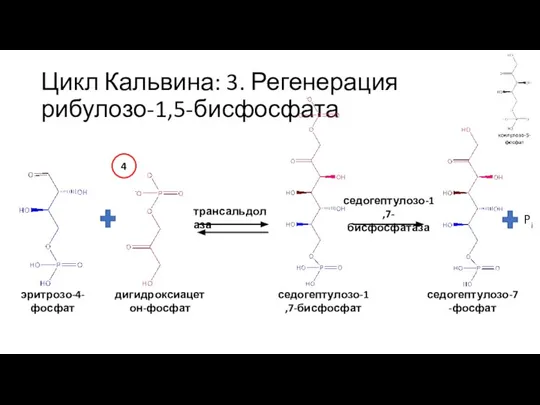
Слайд 37Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
седогептулозо-7-фосфат
глицеральдегид-3-фосфат
транскетолаза
рибозо-5-фосфат
ксилулозо-5-фосфат
5
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
седогептулозо-7-фосфат
глицеральдегид-3-фосфат
транскетолаза
рибозо-5-фосфат
ксилулозо-5-фосфат
5
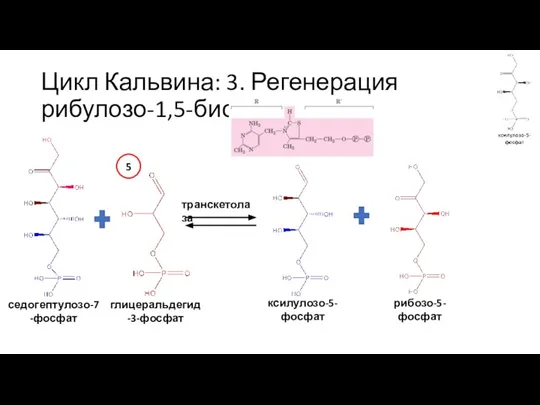
Слайд 38Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
рибозо-5-фосфат
рибозо-5-фосфат изомераза
рибулозо-5-фосфат
рибулозо-5-фосфат изомераза
рибулозо-5-фосфат изомераза
рибулозо-5-фосфат
рибулозо-5-фосфат
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
рибозо-5-фосфат
рибозо-5-фосфат изомераза
рибулозо-5-фосфат
рибулозо-5-фосфат изомераза
рибулозо-5-фосфат изомераза
рибулозо-5-фосфат
рибулозо-5-фосфат
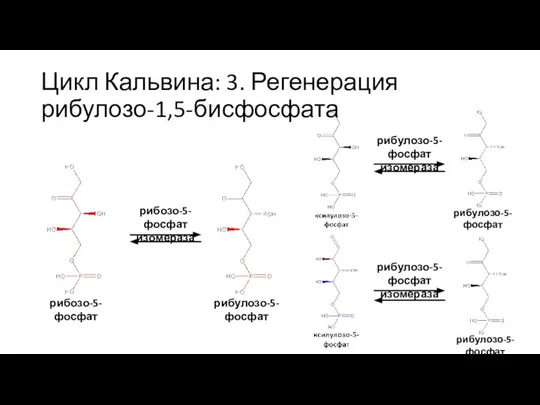
Слайд 39Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
рибулозо-5-фосфат
3×
рибулозо-5-фосфат киназа
рибулозо-1,5-бисфосфат
ATP
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата
рибулозо-5-фосфат
3×
рибулозо-5-фосфат киназа
рибулозо-1,5-бисфосфат
ATP

Слайд 40Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата и суммарное уравнение
5 глицеральдегид-3-фосфат + 3 ATP
Цикл Кальвина: 3. Регенерация рибулозо-1,5-бисфосфата и суммарное уравнение
5 глицеральдегид-3-фосфат + 3 ATP

Слайд 41Фотодыхание
O2 конкурирует с CO2 за активный центр рубиско. В 1 из 3
Фотодыхание
O2 конкурирует с CO2 за активный центр рубиско. В 1 из 3

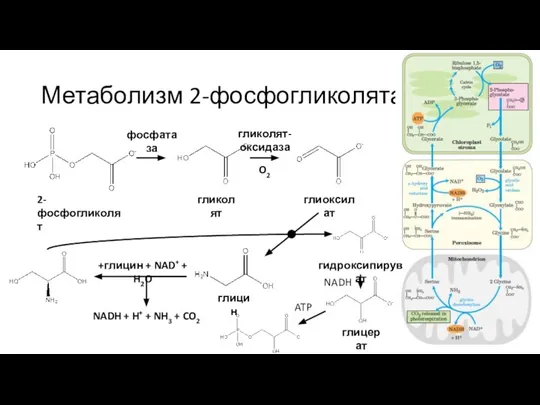
Слайд 42Метаболизм 2-фосфогликолята
2-фосфогликолят
гликолят
фосфатаза
гликолят-оксидаза
O2
глиоксилат
глицин
+глицин + NAD+ + H2O
NADH + H+ + NH3 + CO2
гидроксипируват
глицерат
NADH
ATP
Метаболизм 2-фосфогликолята
2-фосфогликолят
гликолят
фосфатаза
гликолят-оксидаза
O2
глиоксилат
глицин
+глицин + NAD+ + H2O
NADH + H+ + NH3 + CO2
гидроксипируват
глицерат
NADH
ATP
Слайд 43C4-фотосинтез
C4-растения живут при высоких температурах, где отношение концентраций растворенных O2 к CO2
C4-фотосинтез
C4-растения живут при высоких температурах, где отношение концентраций растворенных O2 к CO2

 Биологический алфавит
Биологический алфавит Признаки живых организмов. 5 класс
Признаки живых организмов. 5 класс Технология развивающего обучения на уроках биологии
Технология развивающего обучения на уроках биологии prez_evolyutsia_rasteniy
prez_evolyutsia_rasteniy Презентация на тему БОЛЬШОЙ МИР МАЛЕНЬКИХ КЛЕТОК
Презентация на тему БОЛЬШОЙ МИР МАЛЕНЬКИХ КЛЕТОК  Презентация на тему ЗАГРЯЗНЕНИЕ ГИДРОСФЕРЫ
Презентация на тему ЗАГРЯЗНЕНИЕ ГИДРОСФЕРЫ  Презентация на тему "Стрекоза" - презентации по Биологии
Презентация на тему "Стрекоза" - презентации по Биологии Эволюция выделительной системы
Эволюция выделительной системы Процессы в живой природе
Процессы в живой природе Методика исследования
Методика исследования Презентация на тему Дикие - дикие кошки
Презентация на тему Дикие - дикие кошки  Деревья
Деревья Исәнме, куянкай! Исәнме, керпе!
Исәнме, куянкай! Исәнме, керпе! Жизнедеятельность клетки, ее деление и рост
Жизнедеятельность клетки, ее деление и рост Презентация на тему Класс Круглоротые
Презентация на тему Класс Круглоротые  Вегетативная нервная система
Вегетативная нервная система Презентация на тему Методы изучения клетки
Презентация на тему Методы изучения клетки  Метаболическая биохимия
Метаболическая биохимия Царство грибы. Обобщение и систематизация знаний
Царство грибы. Обобщение и систематизация знаний Шестиногие в фотообъективе. Таинственный мир насекомых
Шестиногие в фотообъективе. Таинственный мир насекомых Что растет на подоконнике
Что растет на подоконнике Органы дыхания
Органы дыхания Двухмембранные органоиды. Митохондрии
Двухмембранные органоиды. Митохондрии 0767PP
0767PP Строение клетки
Строение клетки Экологические системы
Экологические системы Наши органы чувств
Наши органы чувств Анализаторы. Строение и общие закономерности развития сенсорных систем
Анализаторы. Строение и общие закономерности развития сенсорных систем