- Genetic load in human population

Содержание
- 2. Genetic load Genetic load: the extent to which the fitness of an individual is below the
- 3. Genetic load: the difference between the average fitness of the population and the fitness of the
- 4. Types of Genetic Load Three main kinds of genetic load may be recognized: A.Input Load: in
- 5. C. Substitutional Load: Which is generated by selection favouring the replacement of an existing allele by
- 7. Genetic load an Example… Selective death (or genetic death): the chance that an individual will die
- 8. Causes of Genetic Load 1.Deleterious mutation 2.Beneficial mutation 3.Inbreeding 4.Recombination/segregation load
- 9. DELETERIOUS MUTATIONS Deleterious mutation load is the main contributing factor to genetic load overall. Most mutations
- 10. High load can lead to a small population size, which in turn increases the accumulation of
- 11. Beneficial mutation New beneficial mutations create fitter genotypes than those previously present in the population. When
- 12. Motoo Kimura's original argument for the neutral theory of molecular evolution was that if most differences
- 14. Inbreeding Inbreeding increases homozygosity. In the short run, an increase in inbreeding increases the probability with
- 15. Recombination/segregation load Combinations of alleles that have evolved to work well together may not work when
- 16. Recombination load arises through unfavorable combinations across multiple loci that appear when favorable linkage disequilibria are
- 17. Genetic load : Mutation
- 18. Genetic load: segregational Segregational load is a big problem for the balance school: Well known examples
- 19. Balance school would extend this to most polymorphic loci in the genome. Let’s see if this
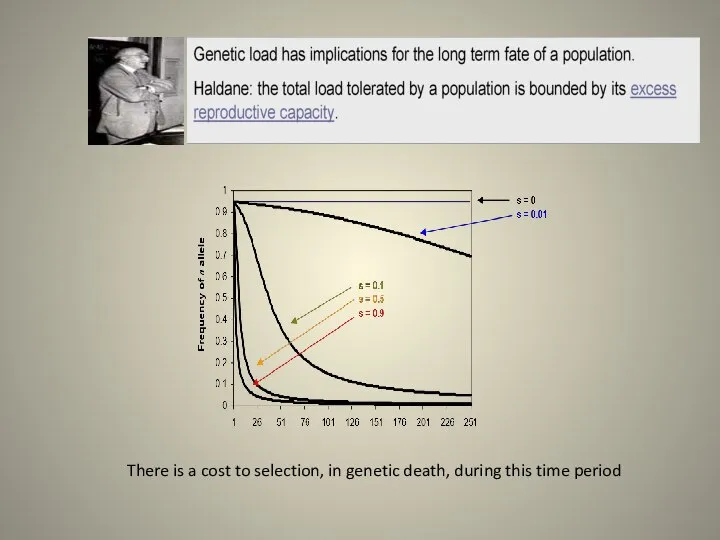
- 20. There is a cost to selection, in genetic death, during this time period
- 21. Migration load Migration load is the result of nonnative organisms that aren’t adapted to a particular
- 22. “It is altogether unlikely that two genes would have identical selective values under all the conditions
- 23. Neo-Darwinism 1930’s: ⎯ no way to test the predictions of different schools. ⎯arguments centered on mathematical
- 24. Defining Directional Section Directional selection: selection that favours the phenotype at an extreme of the range
- 25. Defining two types directional selection Type 1: Positive Darwinian selection: directional selection for fixation of a
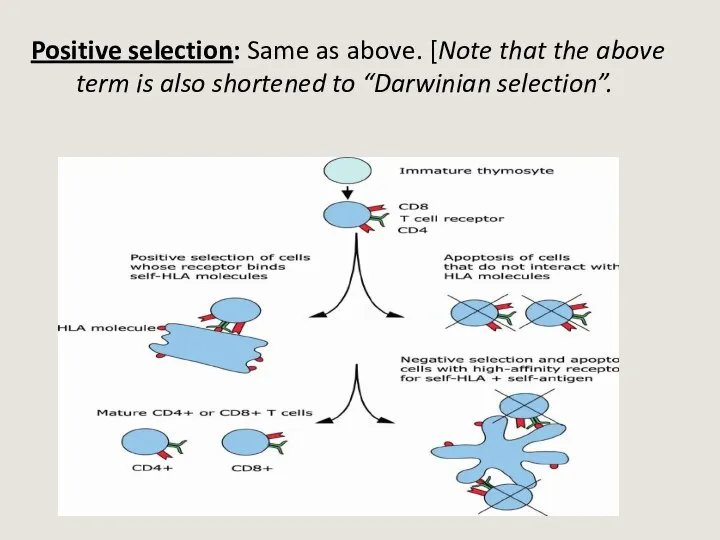
- 26. Positive selection: Same as above. [Note that the above term is also shortened to “Darwinian selection”.
- 27. Type 2: Negative Darwinian selection: directional selection for removal of a new and deleterious mutation from
- 30. Скачать презентацию

Слайд 3Genetic load: the difference between the average fitness of the population and
Genetic load: the difference between the average fitness of the population and

Слайд 4Types of Genetic Load
Three main kinds of genetic load may be recognized:
A.Input
Types of Genetic Load
Three main kinds of genetic load may be recognized:
A.Input

Слайд 5C. Substitutional Load: Which is generated by selection favouring the replacement of
C. Substitutional Load: Which is generated by selection favouring the replacement of

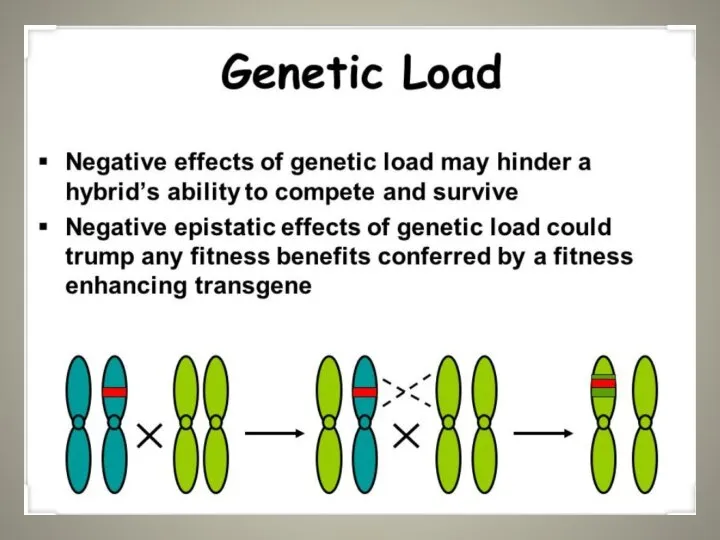
Слайд 7Genetic load an Example…
Selective death (or genetic death): the chance that an
Genetic load an Example… Selective death (or genetic death): the chance that an

Слайд 8Causes of Genetic Load
1.Deleterious mutation
2.Beneficial mutation
3.Inbreeding
4.Recombination/segregation load
Causes of Genetic Load
1.Deleterious mutation
2.Beneficial mutation
3.Inbreeding
4.Recombination/segregation load

Слайд 9DELETERIOUS MUTATIONS
Deleterious mutation load is the main contributing factor to genetic load
DELETERIOUS MUTATIONS
Deleterious mutation load is the main contributing factor to genetic load

Слайд 10High load can lead to a small population size, which in turn
High load can lead to a small population size, which in turn

Слайд 11Beneficial mutation
New beneficial mutations create fitter genotypes than those previously present in
Beneficial mutation
New beneficial mutations create fitter genotypes than those previously present in

Слайд 12Motoo Kimura's original argument for the neutral theory of molecular evolution was
Motoo Kimura's original argument for the neutral theory of molecular evolution was


Слайд 14Inbreeding
Inbreeding increases homozygosity.
In the short run, an increase in inbreeding increases
Inbreeding
Inbreeding increases homozygosity.
In the short run, an increase in inbreeding increases

Слайд 15Recombination/segregation load
Combinations of alleles that have evolved to work well together may
Recombination/segregation load
Combinations of alleles that have evolved to work well together may

Слайд 16Recombination load arises through unfavorable combinations across multiple loci that appear when
Recombination load arises through unfavorable combinations across multiple loci that appear when

Слайд 17Genetic load : Mutation
Genetic load : Mutation

Слайд 18Genetic load: segregational
Segregational load is a big problem for the balance school:
Well
Genetic load: segregational
Segregational load is a big problem for the balance school:
Well

Слайд 19Balance school would extend this to most polymorphic loci in the genome.
Balance school would extend this to most polymorphic loci in the genome.

Слайд 20There is a cost to selection, in genetic death, during this time
There is a cost to selection, in genetic death, during this time
Слайд 21 Migration load
Migration load is the result of nonnative organisms that aren’t adapted
Migration load
Migration load is the result of nonnative organisms that aren’t adapted

Слайд 22“It is altogether unlikely that two genes would have identical selective values
“It is altogether unlikely that two genes would have identical selective values

Слайд 23Neo-Darwinism
1930’s:
⎯ no way to test the predictions of different schools. ⎯arguments
Neo-Darwinism
1930’s:
⎯ no way to test the predictions of different schools. ⎯arguments

Слайд 24Defining Directional Section
Directional selection: selection that favours the phenotype at an extreme
Defining Directional Section
Directional selection: selection that favours the phenotype at an extreme

Слайд 25Defining two types directional selection
Type 1:
Positive Darwinian selection: directional selection for
Defining two types directional selection
Type 1:
Positive Darwinian selection: directional selection for

Слайд 26Positive selection: Same as above. [Note that the above term is also
Positive selection: Same as above. [Note that the above term is also
Слайд 27 Type 2:
Negative Darwinian selection: directional selection for removal of a new
Type 2:
Negative Darwinian selection: directional selection for removal of a new


 Законы Менделя
Законы Менделя Био-арт. Генная инженерия
Био-арт. Генная инженерия Головной мозг
Головной мозг Метаболическая биохимия
Метаболическая биохимия 2
2 Разнообразие растительного мира
Разнообразие растительного мира Презентация на тему Выделение
Презентация на тему Выделение  В мире углеводов
В мире углеводов Проектная работа по биологии Условия прорастания семян
Проектная работа по биологии Условия прорастания семян Круговорот кислорода в природе
Круговорот кислорода в природе Классификация и строение микроорганизмов
Классификация и строение микроорганизмов Панда
Панда Изучение человека. Ученые-анатомы
Изучение человека. Ученые-анатомы Опорно-двигательная система клетки (цитоскелет). Микротрубочки
Опорно-двигательная система клетки (цитоскелет). Микротрубочки Мейоз
Мейоз Пищеварение в желудке
Пищеварение в желудке Мхи, хвощи, плауны, папоротники
Мхи, хвощи, плауны, папоротники Подтип Жгутиконосцы
Подтип Жгутиконосцы Витамины и их роль в организме человека. 8 класс
Витамины и их роль в организме человека. 8 класс Разнообразие лишайников
Разнообразие лишайников Свойства биогеоценозов
Свойства биогеоценозов Современная ззология
Современная ззология Введення в гастроентерологію. Методи обстеження
Введення в гастроентерологію. Методи обстеження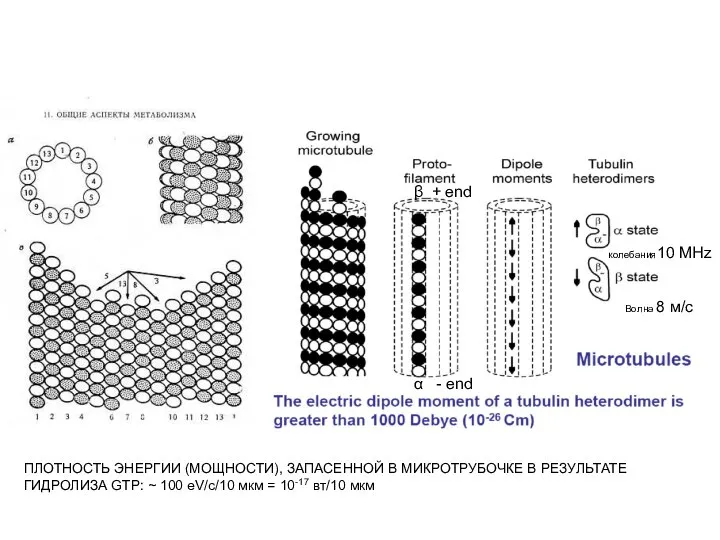 Энергизация микротрубочек
Энергизация микротрубочек Где живут эмоции
Где живут эмоции Хронобиология. Основные термины. Биоритм и его составляющие. Его значение для медицины
Хронобиология. Основные термины. Биоритм и его составляющие. Его значение для медицины Поэзия цветочного мира
Поэзия цветочного мира Вестибулярный анализатор
Вестибулярный анализатор