Гормональная регуляция обменных процессов: белков, воды, натрия, калия, кальция, фосфатов. Лекция № 22
- Гормональная регуляция обменных процессов: белков, воды, натрия, калия, кальция, фосфатов. Лекция № 22

Содержание
- 2. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА БЕЛКОВ
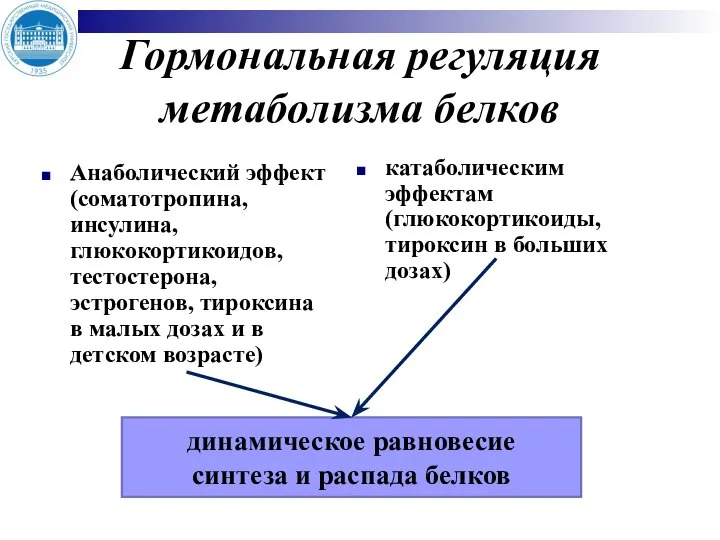
- 3. Гормональная регуляция метаболизма белков Анаболический эффект (соматотропина, инсулина, глюкокортикоидов, тестостерона, эстрогенов, тироксина в малых дозах и
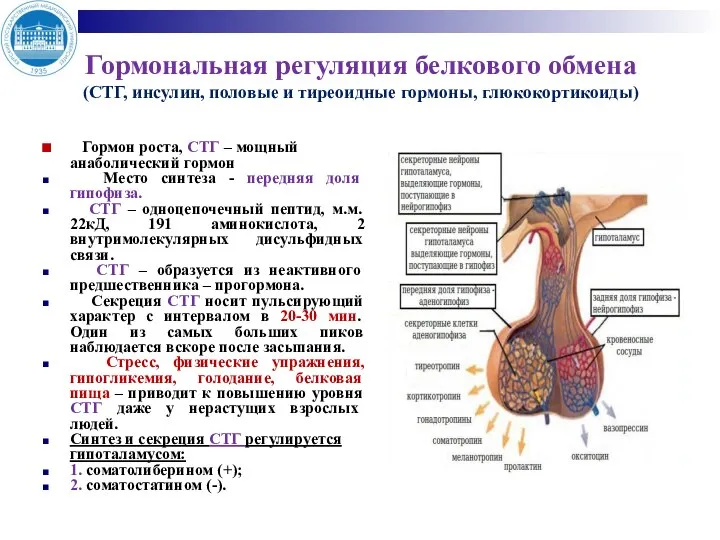
- 4. Гормональная регуляция белкового обмена (СТГ, инсулин, половые и тиреоидные гормоны, глюкокортикоиды) Гормон роста, СТГ – мощный
- 5. Рецепторы для СТГ расположены в плазматической мембране клеток 1. Гормозависимые ткани: скелетные мышцы, хрящи, кости, соединительная
- 6. Действие СТГ на рост скелета и мягких тканей опосредуется соматомединами Это соединения с инсулиноподобной активностью и
- 7. Биологическое действие СТГ 1. Начальные эффекты (мышцы, хрящи, кости, соединительная ткань) увеличение проницаемости клеточных мембран для
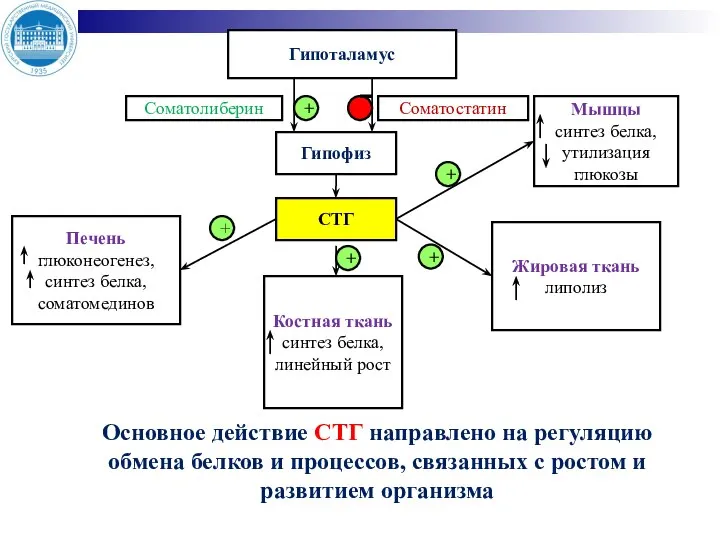
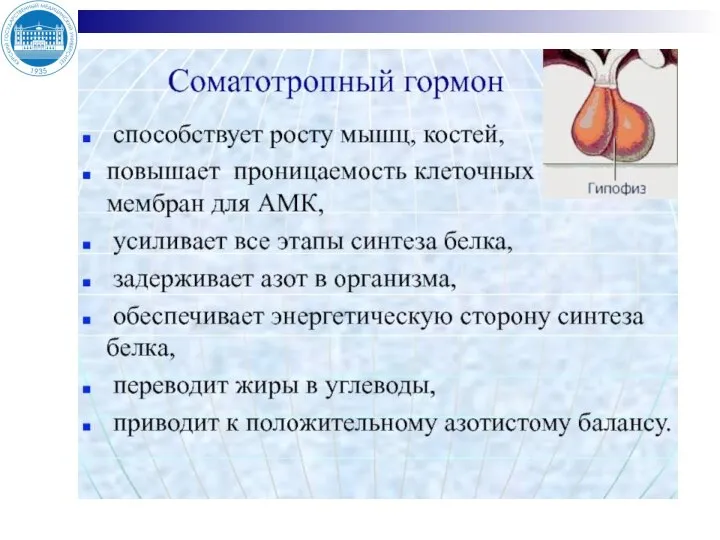
- 8. Гипоталамус Соматолиберин Гипофиз Соматостатин СТГ Печень глюконеогенез, синтез белка, соматомединов Костная ткань синтез белка, линейный рост
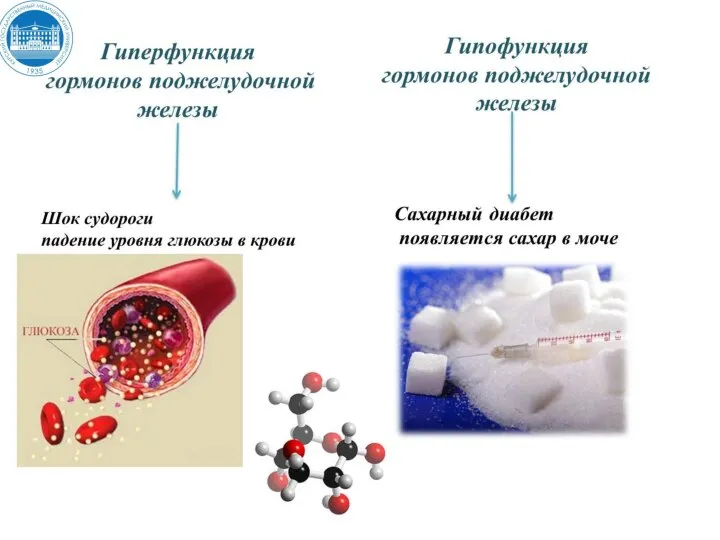
- 10. Патология синтеза и секреции СТГ Гипофункция с рождения - гипофизарная карликовость (дефицит СТГ из-за мутации гена
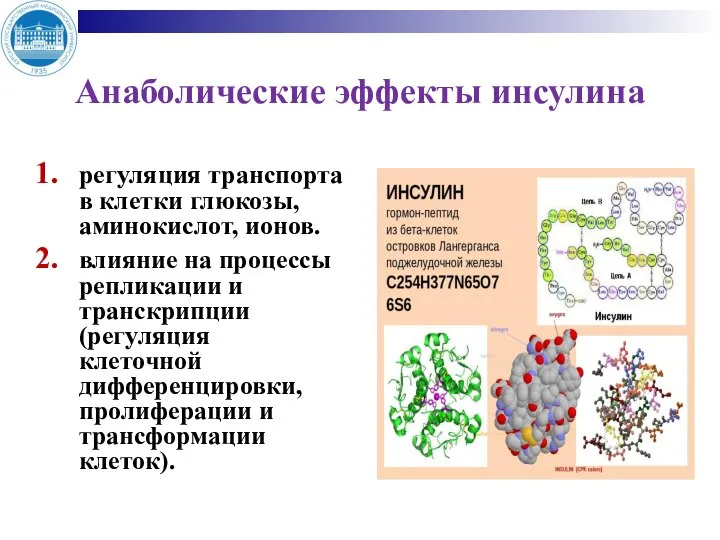
- 12. Анаболические эффекты инсулина регуляция транспорта в клетки глюкозы, аминокислот, ионов. влияние на процессы репликации и транскрипции
- 13. Анаболические эффекты инсулина Начальные эффекты (в течении нескольких секунд-минут, изменение трансмембранного транспорта, фосфорилирование и дефосфорилирование белков,
- 15. Йодтиронины 1. Синтез и секреция йодтиронинов (Т3 и Т4) регулируется гипоталамо-гипофизарной системой по принципу обратной связи.
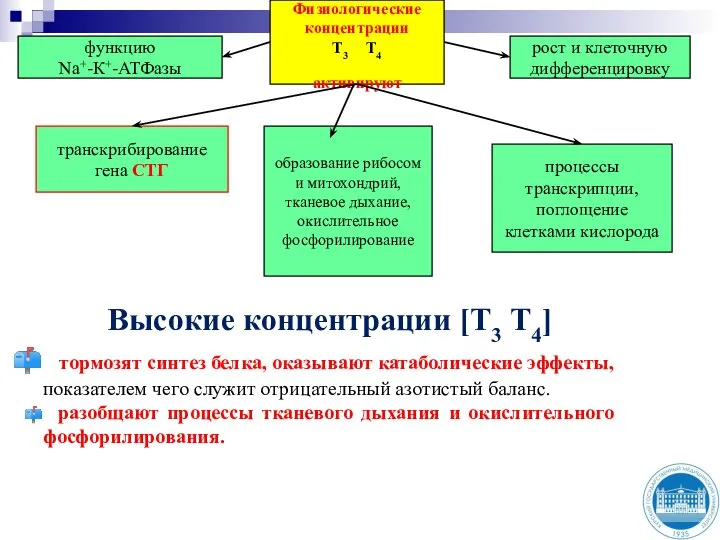
- 16. Физиологические концентрации Т3 Т4 активируют функцию Na+-К+-АТФазы рост и клеточную дифференцировку процессы транскрипции, поглощение клетками кислорода
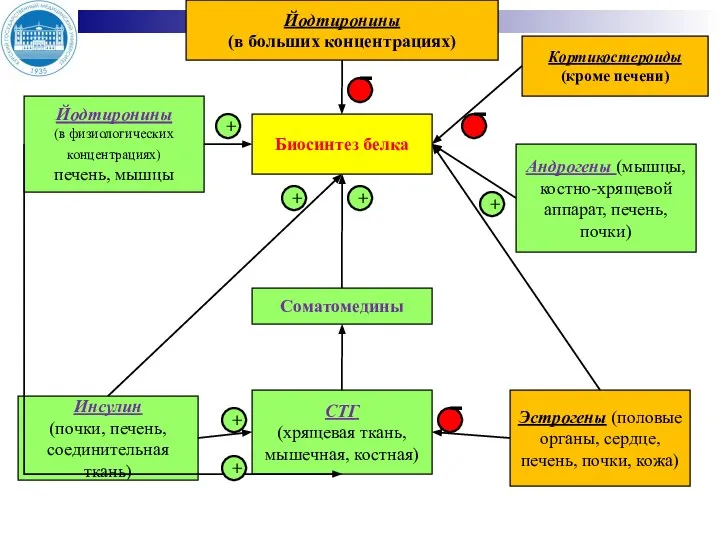
- 17. Йодтиронины (в больших концентрациях) Йодтиронины (в физиологических концентрациях) печень, мышцы Инсулин (почки, печень, соединительная ткань) Биосинтез
- 18. Заболевания щитовидной железы Гипотиреоз у новорожденных приводит к развитию кретинизма – тяжелое, необратимое нарушение умственного развития.
- 19. Заболевания щитовидной железы
- 21. Половые гормоны Андрогены обладают выраженным анаболическим эффектом в печени, почках, костно-хрящевом аппарате, покровных тканях, мужских половых
- 23. Влияние глюкокортикоидов на обмен белков Проявляется двояко: В печени кортизол в основном оказывает анаболический эффект (стимулирует
- 33. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВОДЫ, НАТРИЯ, КАЛИЯ
- 34. ПАРАМЕТРЫ ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА рН Осмотическое давление V внутриклеточной жидкости V внеклеточной жидкости Изменение этих параметров может
- 35. ГОРМОНЫ, УЧАСТВУЮЩИЕ В РЕГУЛЯЦИИ ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА АДГ (АНТИДИУРЕТИЧЕСКИЙ ГОРМОН, ВАЗОПРЕССИН), АЛЬДОСТЕРОН Предсердный натриуретический фактор (ПНФ).
- 36. Регуляция водно-солевого гомеостаза
- 38. ХАРАКТЕРИСТИКА АДГ АДГ – пептид, м.м. 1100 Д, содержит 9 АМК, одна дисульфидная связь Синтез и
- 39. СТИМУЛ ДЛЯ СЕКРЕЦИИ АДГ [Na+] осмотического давления внеклеточной жидкости Осморецепторы гипоталамуса регистрируют : Р осмотического нервные
- 40. Секреция и механизм действия антидиуретического гормона
- 41. Секреция и механизм действия антидиуретического гормона А: 1 - супраоптический нейрон; 2 - паравентрикулярный нейрон; 3
- 42. МЕХАНИЗМ ДЕЙСТВИЯ АДГ Для АДГ существует 2 типа рецепторов: V1 и V2 V2 – опосредуют главный
- 43. ЭТАПЫ ДЕЙСТВИЯ АДГ НА ЭПИТЕЛИАЛЬНЫЕ КЛЕТКИ ДИСТАЛЬНЫХ КАНАЛЬЦЕВ И СОБИРАТЕЛЬНЫХ ТРУБОЧЕК ПОЧЕК
- 44. НЕСАХАРНЫЙ ДИАБЕТ Причиной является дефицит АДГ 1) генетические дефекты синтеза препро-АДГ в гипоталамусе; 2) дефекты процессинга
- 45. АЛЬДОСТЕРОН Синтез и секреция непосредственно стимулируются: понижением [Na+] и повышением [K+] в плазме крови простагландинами, АКТГ
- 46. МЕХАНИЗМ ДЕЙСТВИЯ АЛЬДОСТЕРОНА Альдостерон, взаимодействуя с внутриклеточными рецепторами, стимулирует синтез белков. Эти белки могут быть: 1
- 47. СУММАРНЫЙ БИОЛОГИЧЕСКИЙ ЭФФЕКТ АЛЬДОСТЕРОНА Увеличение реабсорбции ионов Na+ в канальцах нефронов Задержка NaCl в организме и
- 48. РОЛЬ СИСТЕМЫ РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОН В РЕГУЛЯЦИИ ВОДНО-СОЛЕВОГО ОБМЕНА Повышение осмотического давления
- 49. ГИПЕРАЛЬДОСТЕРОНИЗМ Заболевание вызванное гиперсекрецией альдостерона надпочечниками. Причина первичного гиперальдостеронизма (синдром Конна) – аденома надпочечников, диффузная гипертрофия
- 50. ВТОРИЧНЫЙ ГИПЕРАЛЬДОСТЕРОНИЗМ Встречается чаще, чем первичный. Причины: сердечная недостаточность, хронические заболевания почек, опухоли секретирующие ренин. Клиника:
- 52. ХАРАКТЕРИСТИКА ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ) Физиологический антагонист ангиотензина II. Этот пептид, 28 АМК, с одним дисульфидным
- 53. СТИМУЛЫ ДЛЯ СЕКРЕЦИИ ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
- 54. ОСНОВНЫЕ КЛЕТКИ-МИШЕНИ ПНФ: 1.почки, 2.периферические артерии. расширение приносящих артериол. усиление почечного кровотока. увеличение скорости фильтрации и
- 55. Эффекты действия ПНФ 1 - ингибирует выделение ренина; 2 - ингибирует секрецию альдостерона; 3 - ингибирует
- 56. Гормональные нарущения водно-солевого обмена
- 58. Гормональные нарушения обмена натрия и калия
- 61. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА КАЛЬЦИЯ И ФОСФАТОВ
- 62. РОЛЬ ИОНОВ КАЛЬЦИЯ В ОБМЕННЫХ ПРОЦЕССАХ: структурный компонент костной ткани; играет ключевую роль в мышечном сокращении;
- 63. В организме взрослого человека содержится до 1,5 кг Ca2+. Пределы колебаний Ca2+ в крови: 2,12 –
- 66. ХАРАКТЕРИСТИКА ПАРАТГОРМОНА ПТГ – одноцепочечный полипептид, 84 АМК. Действие ПТГ: 1. Повышение [Ca2+]; 2. Снижение [фосфатов]
- 67. ЭФФЕКТЫ ПАРАТГОРМОНА
- 68. БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ПАРАТГОРМОНА 1 - ПТГ стимулирует мобилизацию кальция из кости; 2 - ПТГ стимулирует реабсорбцию
- 70. Схема синтеза кальцитриола Кожа: Провитамин Д3 (7-дегидрохолестерол) Витамин Д3 (холекальциферол) (связывается с белком-транскальциферином, альбумином и поступает
- 71. Обмен витамина D (D2 – эрго -; D3 – холекальциферол) Пища (молочные продукты, печень) Кожа Ультрафиолетовое
- 72. Обмен витамина D Трансферрин (Транскальциферрин) Печень D2(3) Кровь 25-окси D (кальцидиол) 25-окси D (кальцидиол) Гидроксилирование в
- 73. Витамин D Почки Клетки-мишени 25-окси D2(3) Гидроксилирование в 1 положении-активация паратгормоном (при снижении уровня в крови
- 74. ЭФФЕКТЫ КАЛЬЦИТРИОЛА [1,25 (ОН)2 Д3] Клетки кишечника: Индуцируют синтез Ca2+ - переносящих белков, которые обеспечивают всасывание
- 75. ХАРАКТЕРИСТИКА КАЛЬЦИТОНИНА Кальцитонин – полипептид, 32 АМК. Секретируется: 1. парафолликулярными К – клетками щитовидной железы 2.
- 76. Ингибирует высвобождение Са2+ из костной ткани; Стимулирует экскрецию Са2+ и фосфатов с мочой; Снижение содержания кальция
- 78. Гормональные нарушения обмена кальция
- 81. Скачать презентацию

Слайд 3Гормональная регуляция
метаболизма белков
Анаболический эффект (соматотропина, инсулина, глюкокортикоидов, тестостерона, эстрогенов, тироксина в
Гормональная регуляция
метаболизма белков
Анаболический эффект (соматотропина, инсулина, глюкокортикоидов, тестостерона, эстрогенов, тироксина в
Слайд 4Гормональная регуляция белкового обмена
(СТГ, инсулин, половые и тиреоидные гормоны, глюкокортикоиды)
Гормон
Гормональная регуляция белкового обмена
(СТГ, инсулин, половые и тиреоидные гормоны, глюкокортикоиды)
Гормон
Слайд 5Рецепторы для СТГ расположены в плазматической мембране клеток
1. Гормозависимые ткани:
скелетные мышцы,
хрящи,
Рецепторы для СТГ расположены в плазматической мембране клеток
1. Гормозависимые ткани:
скелетные мышцы,
хрящи,

Слайд 6Действие СТГ на рост скелета и мягких тканей опосредуется соматомединами
Это соединения с
Действие СТГ на рост скелета и мягких тканей опосредуется соматомединами
Это соединения с

Слайд 7Биологическое действие СТГ
1. Начальные эффекты
(мышцы, хрящи, кости, соединительная ткань)
увеличение проницаемости клеточных
Биологическое действие СТГ
1. Начальные эффекты
(мышцы, хрящи, кости, соединительная ткань)
увеличение проницаемости клеточных

Слайд 8Гипоталамус
Соматолиберин
Гипофиз
Соматостатин
СТГ
Печень
глюконеогенез, синтез белка, соматомединов
Костная ткань
синтез белка, линейный рост
Жировая ткань липолиз
Мышцы
синтез
Гипоталамус
Соматолиберин
Гипофиз
Соматостатин
СТГ
Печень
глюконеогенез, синтез белка, соматомединов
Костная ткань
синтез белка, линейный рост
Жировая ткань липолиз
Мышцы
синтез
Слайд 10Патология синтеза и секреции СТГ
Гипофункция с рождения - гипофизарная карликовость (дефицит СТГ
Патология синтеза и секреции СТГ
Гипофункция с рождения - гипофизарная карликовость (дефицит СТГ


Слайд 12Анаболические эффекты инсулина
регуляция транспорта в клетки глюкозы, аминокислот, ионов.
влияние на процессы репликации
Анаболические эффекты инсулина
регуляция транспорта в клетки глюкозы, аминокислот, ионов.
влияние на процессы репликации
Слайд 13Анаболические эффекты инсулина
Начальные эффекты (в течении нескольких секунд-минут, изменение трансмембранного транспорта, фосфорилирование
Анаболические эффекты инсулина
Начальные эффекты (в течении нескольких секунд-минут, изменение трансмембранного транспорта, фосфорилирование

Слайд 15Йодтиронины
1. Синтез и секреция йодтиронинов (Т3 и Т4) регулируется гипоталамо-гипофизарной системой по
Йодтиронины
1. Синтез и секреция йодтиронинов (Т3 и Т4) регулируется гипоталамо-гипофизарной системой по

Слайд 16Физиологические концентрации
Т3 Т4
активируют
функцию
Na+-К+-АТФазы
рост и клеточную дифференцировку
процессы транскрипции, поглощение клетками кислорода
образование
Физиологические концентрации
Т3 Т4
активируют
функцию
Na+-К+-АТФазы
рост и клеточную дифференцировку
процессы транскрипции, поглощение клетками кислорода
образование
Слайд 17Йодтиронины
(в больших концентрациях)
Йодтиронины
(в физиологических концентрациях)
печень, мышцы
Инсулин
(почки, печень, соединительная
Йодтиронины
(в больших концентрациях)
Йодтиронины
(в физиологических концентрациях)
печень, мышцы
Инсулин
(почки, печень, соединительная
Слайд 18Заболевания щитовидной железы
Гипотиреоз у новорожденных приводит к развитию кретинизма – тяжелое, необратимое
Заболевания щитовидной железы
Гипотиреоз у новорожденных приводит к развитию кретинизма – тяжелое, необратимое

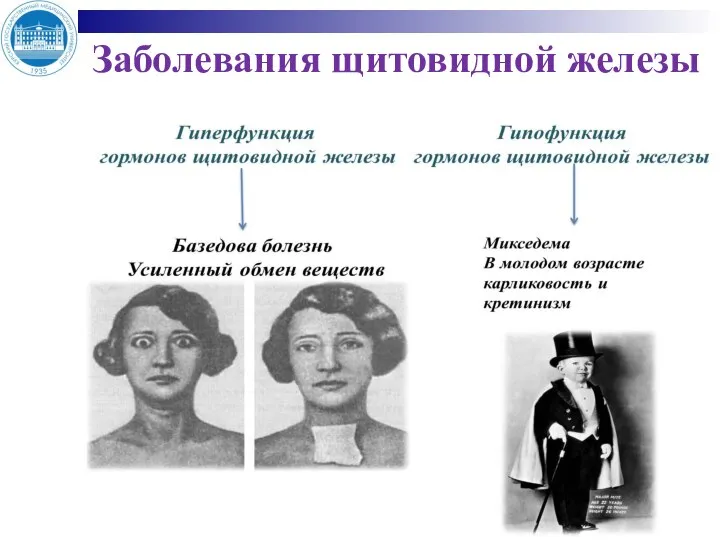
Слайд 19Заболевания щитовидной железы
Заболевания щитовидной железы
Слайд 21Половые гормоны
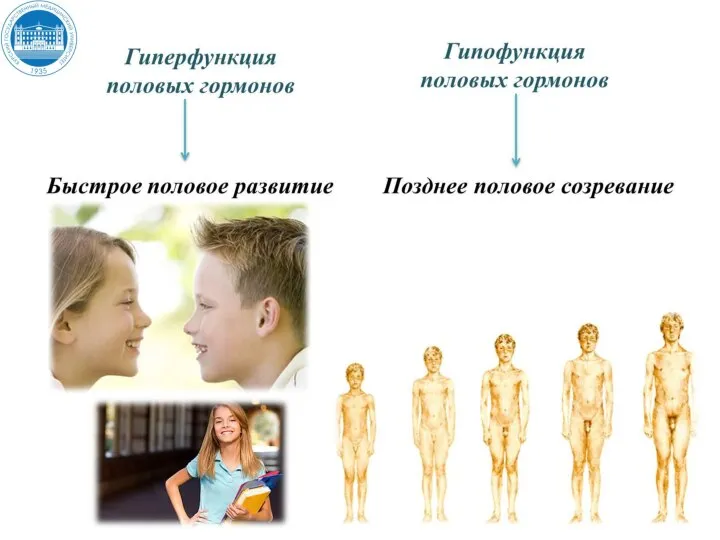
Андрогены обладают выраженным анаболическим эффектом в печени, почках, костно-хрящевом аппарате, покровных
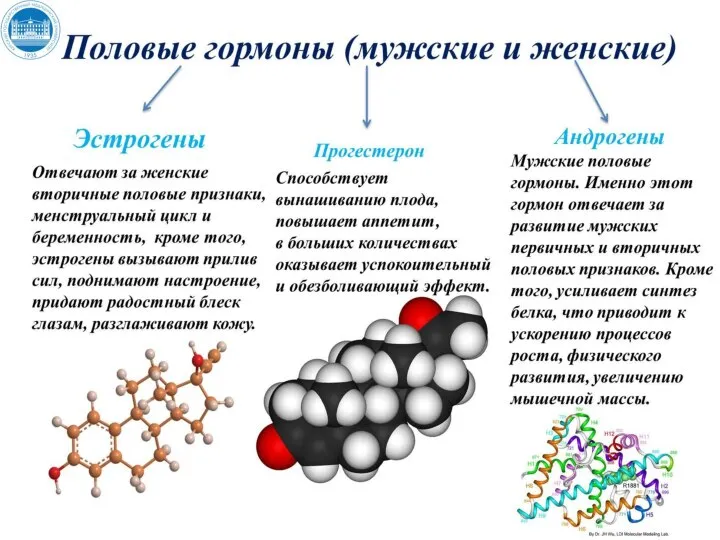
Половые гормоны
Андрогены обладают выраженным анаболическим эффектом в печени, почках, костно-хрящевом аппарате, покровных

Слайд 23Влияние глюкокортикоидов
на обмен белков
Проявляется двояко:
В печени кортизол в основном оказывает анаболический
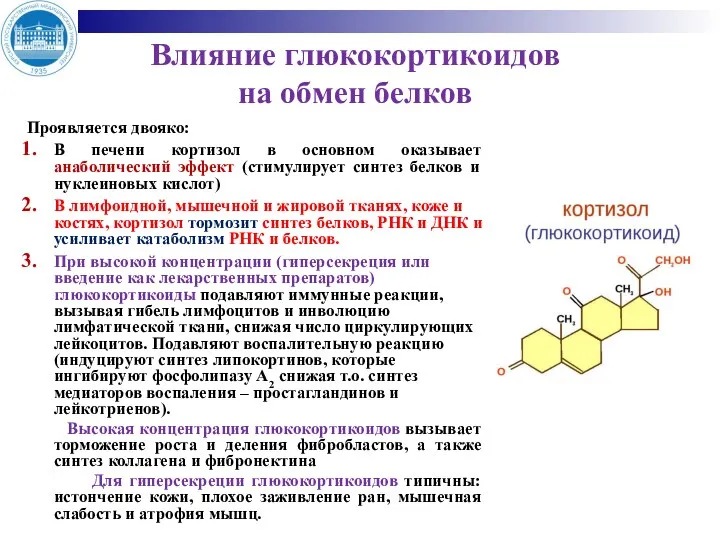
Влияние глюкокортикоидов
на обмен белков
Проявляется двояко:
В печени кортизол в основном оказывает анаболический



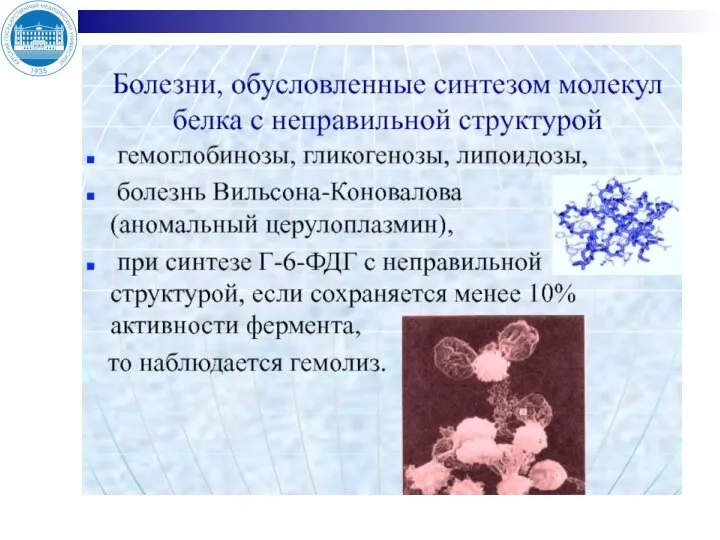




Слайд 33ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВОДЫ, НАТРИЯ, КАЛИЯ
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВОДЫ, НАТРИЯ, КАЛИЯ

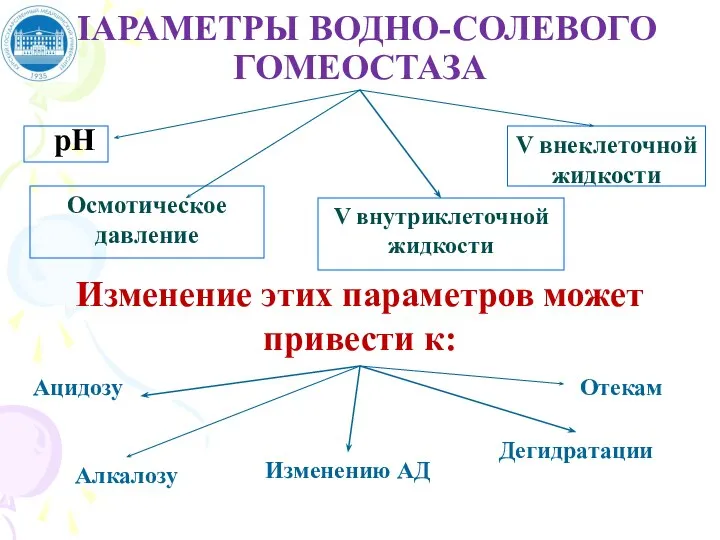
Слайд 34ПАРАМЕТРЫ ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА
рН
Осмотическое давление
V внутриклеточной жидкости
V внеклеточной жидкости
Изменение этих параметров может
ПАРАМЕТРЫ ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА
рН
Осмотическое давление
V внутриклеточной жидкости
V внеклеточной жидкости
Изменение этих параметров может
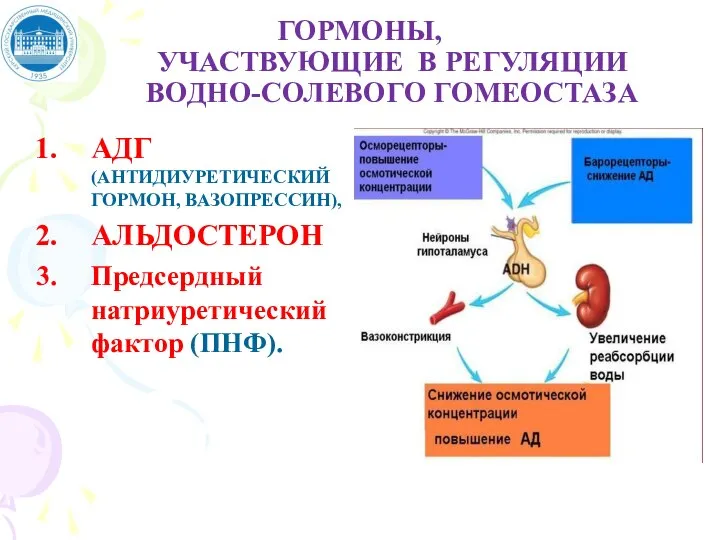
Слайд 35ГОРМОНЫ,
УЧАСТВУЮЩИЕ В РЕГУЛЯЦИИ
ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА
АДГ (АНТИДИУРЕТИЧЕСКИЙ ГОРМОН, ВАЗОПРЕССИН),
АЛЬДОСТЕРОН
Предсердный натриуретический фактор (ПНФ).
ГОРМОНЫ,
УЧАСТВУЮЩИЕ В РЕГУЛЯЦИИ
ВОДНО-СОЛЕВОГО ГОМЕОСТАЗА
АДГ (АНТИДИУРЕТИЧЕСКИЙ ГОРМОН, ВАЗОПРЕССИН),
АЛЬДОСТЕРОН
Предсердный натриуретический фактор (ПНФ).
Слайд 36Регуляция водно-солевого гомеостаза
Регуляция водно-солевого гомеостаза


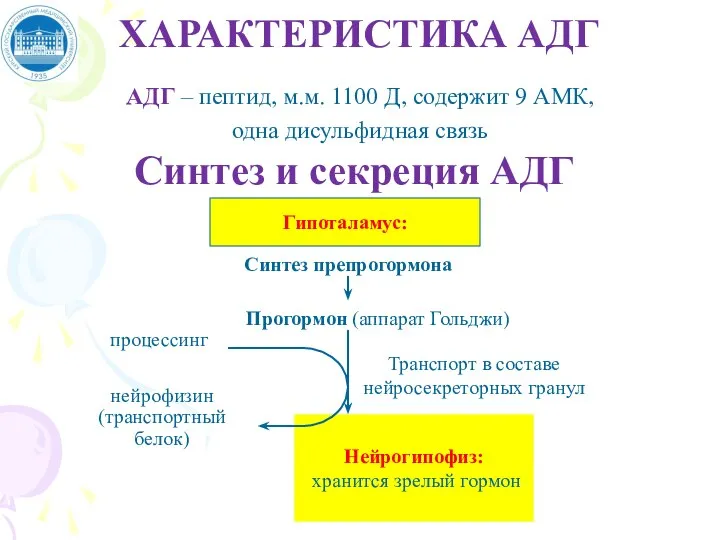
Слайд 38ХАРАКТЕРИСТИКА АДГ
АДГ – пептид, м.м. 1100 Д, содержит 9 АМК,
одна дисульфидная
ХАРАКТЕРИСТИКА АДГ
АДГ – пептид, м.м. 1100 Д, содержит 9 АМК,
одна дисульфидная
Слайд 39
СТИМУЛ
ДЛЯ СЕКРЕЦИИ АДГ
[Na+]
осмотического давления внеклеточной жидкости
Осморецепторы гипоталамуса регистрируют :
Р
СТИМУЛ
ДЛЯ СЕКРЕЦИИ АДГ
[Na+]
осмотического давления внеклеточной жидкости
Осморецепторы гипоталамуса регистрируют :
Р
![СТИМУЛ ДЛЯ СЕКРЕЦИИ АДГ [Na+] осмотического давления внеклеточной жидкости Осморецепторы гипоталамуса регистрируют](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1046461/slide-38.jpg)
Слайд 40Секреция и механизм действия антидиуретического гормона
Секреция и механизм действия антидиуретического гормона
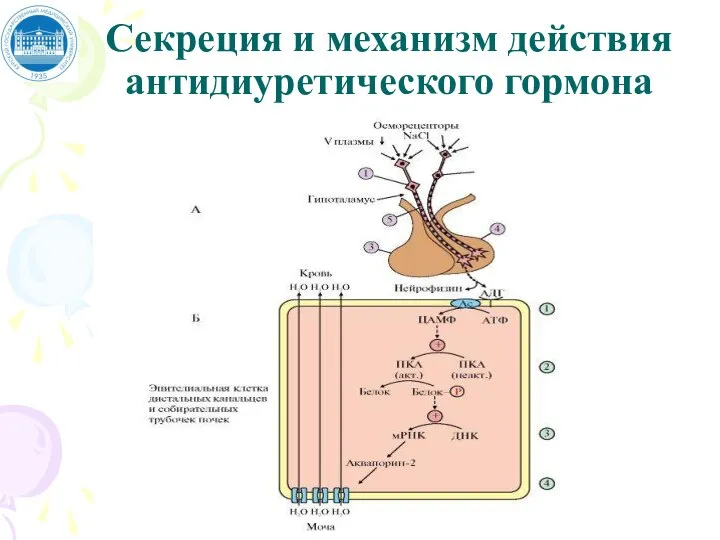
Слайд 41Секреция и механизм действия антидиуретического гормона
А: 1 - супраоптический нейрон; 2 -
Секреция и механизм действия антидиуретического гормона
А: 1 - супраоптический нейрон; 2 -

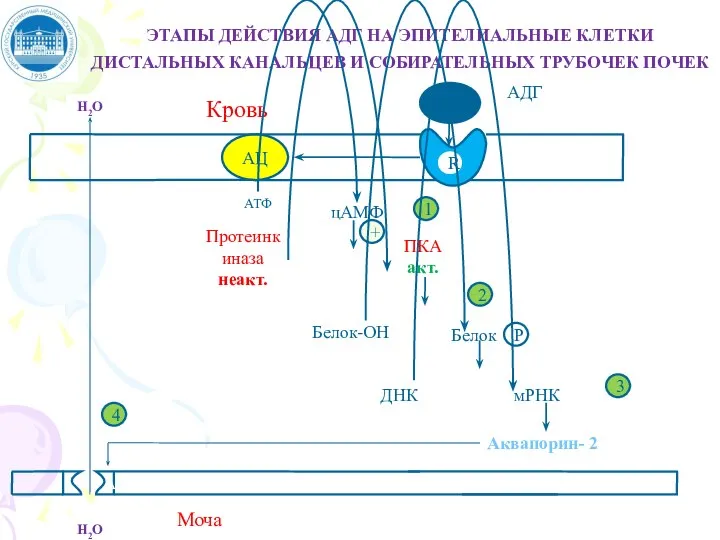
Слайд 42МЕХАНИЗМ ДЕЙСТВИЯ АДГ
Для АДГ существует 2 типа рецепторов: V1 и V2
V2 –
МЕХАНИЗМ ДЕЙСТВИЯ АДГ
Для АДГ существует 2 типа рецепторов: V1 и V2
V2 –

Слайд 43ЭТАПЫ ДЕЙСТВИЯ АДГ НА ЭПИТЕЛИАЛЬНЫЕ КЛЕТКИ ДИСТАЛЬНЫХ КАНАЛЬЦЕВ И СОБИРАТЕЛЬНЫХ ТРУБОЧЕК ПОЧЕК
ЭТАПЫ ДЕЙСТВИЯ АДГ НА ЭПИТЕЛИАЛЬНЫЕ КЛЕТКИ ДИСТАЛЬНЫХ КАНАЛЬЦЕВ И СОБИРАТЕЛЬНЫХ ТРУБОЧЕК ПОЧЕК
Слайд 44НЕСАХАРНЫЙ ДИАБЕТ
Причиной является дефицит АДГ
1) генетические дефекты синтеза препро-АДГ в
НЕСАХАРНЫЙ ДИАБЕТ
Причиной является дефицит АДГ
1) генетические дефекты синтеза препро-АДГ в

Слайд 45АЛЬДОСТЕРОН
Синтез и секреция непосредственно стимулируются:
понижением [Na+] и повышением [K+] в плазме
АЛЬДОСТЕРОН
Синтез и секреция непосредственно стимулируются:
понижением [Na+] и повышением [K+] в плазме
![АЛЬДОСТЕРОН Синтез и секреция непосредственно стимулируются: понижением [Na+] и повышением [K+] в](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1046461/slide-44.jpg)
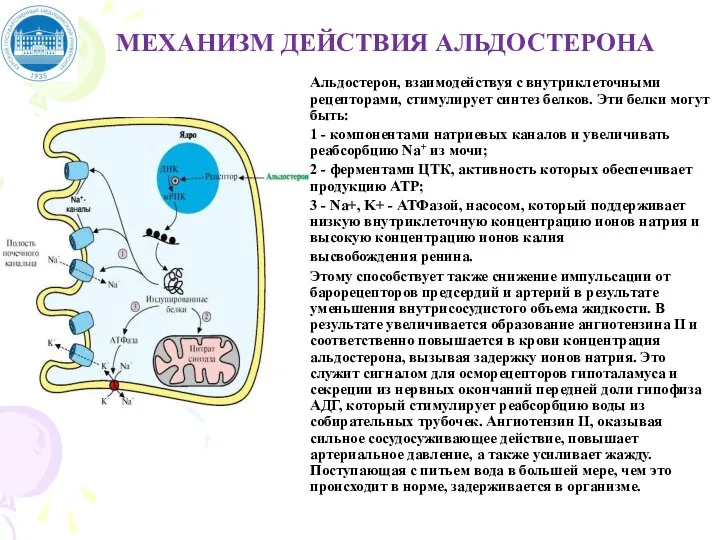
Слайд 46МЕХАНИЗМ ДЕЙСТВИЯ АЛЬДОСТЕРОНА
Альдостерон, взаимодействуя с внутриклеточными рецепторами, стимулирует синтез белков. Эти белки
МЕХАНИЗМ ДЕЙСТВИЯ АЛЬДОСТЕРОНА
Альдостерон, взаимодействуя с внутриклеточными рецепторами, стимулирует синтез белков. Эти белки
Слайд 47СУММАРНЫЙ БИОЛОГИЧЕСКИЙ ЭФФЕКТ АЛЬДОСТЕРОНА
Увеличение реабсорбции ионов Na+ в канальцах нефронов
Задержка NaCl в
СУММАРНЫЙ БИОЛОГИЧЕСКИЙ ЭФФЕКТ АЛЬДОСТЕРОНА
Увеличение реабсорбции ионов Na+ в канальцах нефронов
Задержка NaCl в

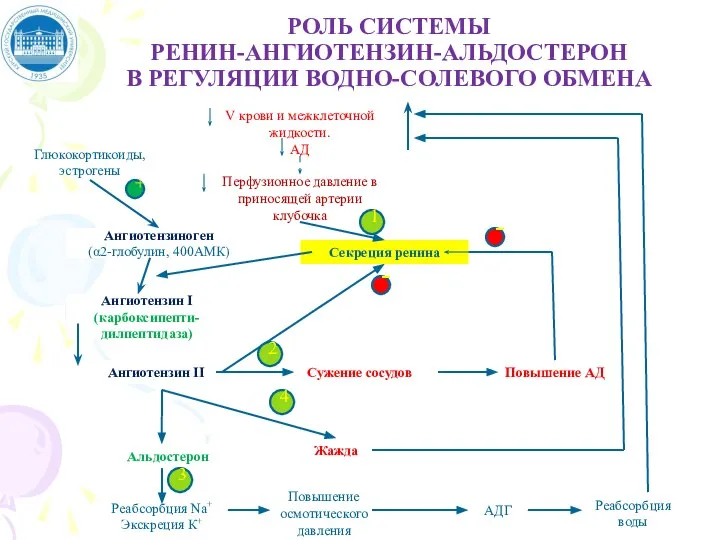
Слайд 48РОЛЬ СИСТЕМЫ
РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОН
В РЕГУЛЯЦИИ ВОДНО-СОЛЕВОГО ОБМЕНА
Повышение осмотического давления
РОЛЬ СИСТЕМЫ
РЕНИН-АНГИОТЕНЗИН-АЛЬДОСТЕРОН
В РЕГУЛЯЦИИ ВОДНО-СОЛЕВОГО ОБМЕНА
Повышение осмотического давления
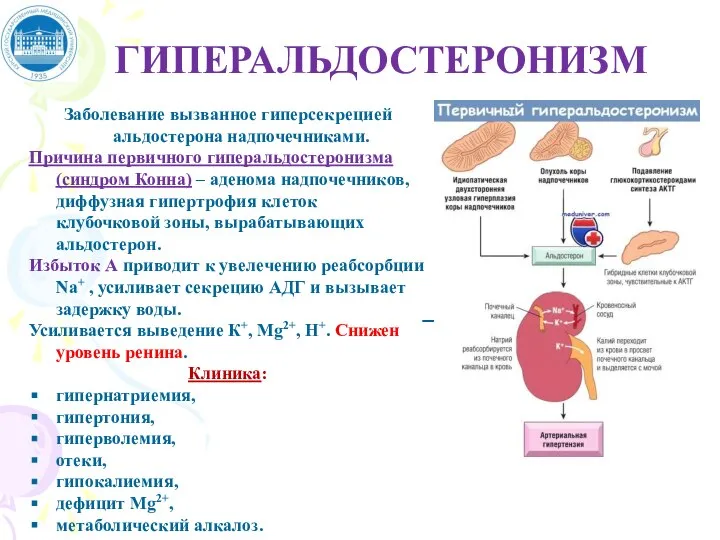
Слайд 49ГИПЕРАЛЬДОСТЕРОНИЗМ
Заболевание вызванное гиперсекрецией альдостерона надпочечниками.
Причина первичного гиперальдостеронизма (синдром Конна) – аденома надпочечников,
ГИПЕРАЛЬДОСТЕРОНИЗМ
Заболевание вызванное гиперсекрецией альдостерона надпочечниками.
Причина первичного гиперальдостеронизма (синдром Конна) – аденома надпочечников,
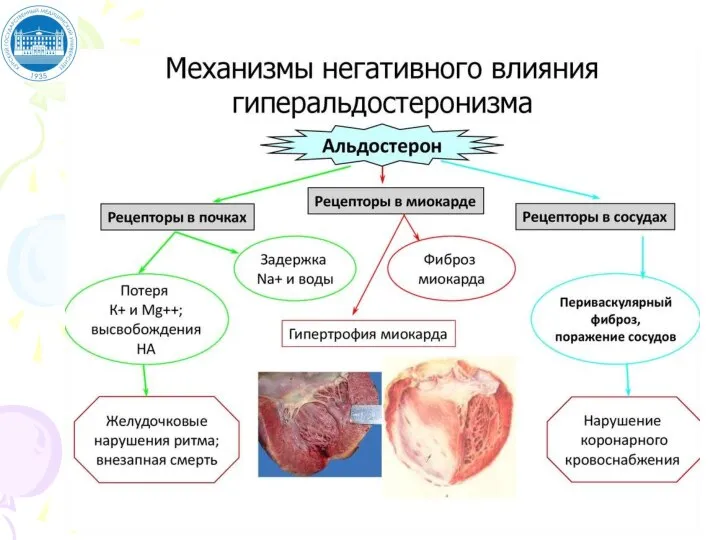
Слайд 50ВТОРИЧНЫЙ ГИПЕРАЛЬДОСТЕРОНИЗМ
Встречается чаще, чем первичный.
Причины: сердечная недостаточность, хронические заболевания почек, опухоли секретирующие
ВТОРИЧНЫЙ ГИПЕРАЛЬДОСТЕРОНИЗМ
Встречается чаще, чем первичный.
Причины: сердечная недостаточность, хронические заболевания почек, опухоли секретирующие

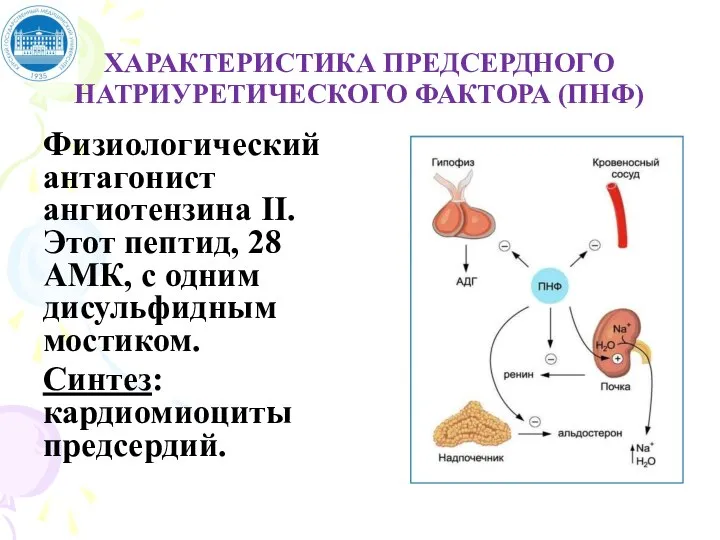
Слайд 52ХАРАКТЕРИСТИКА ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
Физиологический антагонист ангиотензина II. Этот пептид, 28 АМК,
ХАРАКТЕРИСТИКА ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
Физиологический антагонист ангиотензина II. Этот пептид, 28 АМК,
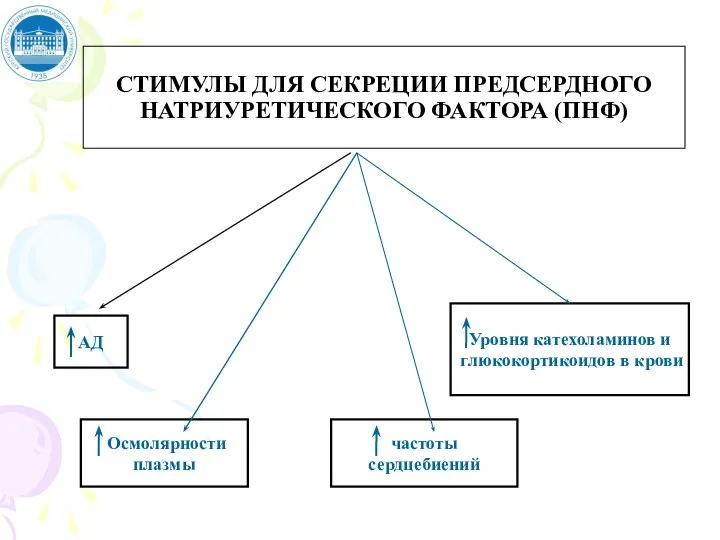
Слайд 53СТИМУЛЫ ДЛЯ СЕКРЕЦИИ ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
СТИМУЛЫ ДЛЯ СЕКРЕЦИИ ПРЕДСЕРДНОГО НАТРИУРЕТИЧЕСКОГО ФАКТОРА (ПНФ)
Слайд 54ОСНОВНЫЕ КЛЕТКИ-МИШЕНИ ПНФ:
1.почки,
2.периферические артерии.
расширение приносящих артериол.
усиление почечного кровотока.
увеличение скорости фильтрации и
ОСНОВНЫЕ КЛЕТКИ-МИШЕНИ ПНФ:
1.почки,
2.периферические артерии.
расширение приносящих артериол.
усиление почечного кровотока.
увеличение скорости фильтрации и

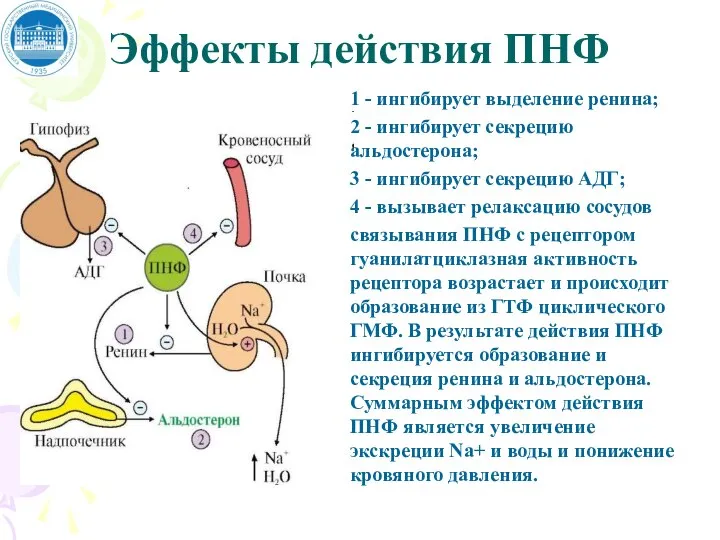
Слайд 55Эффекты действия ПНФ
1 - ингибирует выделение ренина;
2 - ингибирует секрецию альдостерона;
Эффекты действия ПНФ
1 - ингибирует выделение ренина;
2 - ингибирует секрецию альдостерона;
Слайд 56Гормональные нарущения
водно-солевого обмена
Гормональные нарущения
водно-солевого обмена

Слайд 58Гормональные нарушения обмена натрия и калия
Гормональные нарушения обмена натрия и калия



Слайд 61ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА КАЛЬЦИЯ И ФОСФАТОВ
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА КАЛЬЦИЯ И ФОСФАТОВ

Слайд 62РОЛЬ ИОНОВ КАЛЬЦИЯ В ОБМЕННЫХ ПРОЦЕССАХ:
структурный компонент костной ткани;
играет ключевую роль в
РОЛЬ ИОНОВ КАЛЬЦИЯ В ОБМЕННЫХ ПРОЦЕССАХ:
структурный компонент костной ткани;
играет ключевую роль в

Слайд 63В организме взрослого человека содержится до 1,5 кг Ca2+.
Пределы колебаний Ca2+ в
В организме взрослого человека содержится до 1,5 кг Ca2+.
Пределы колебаний Ca2+ в

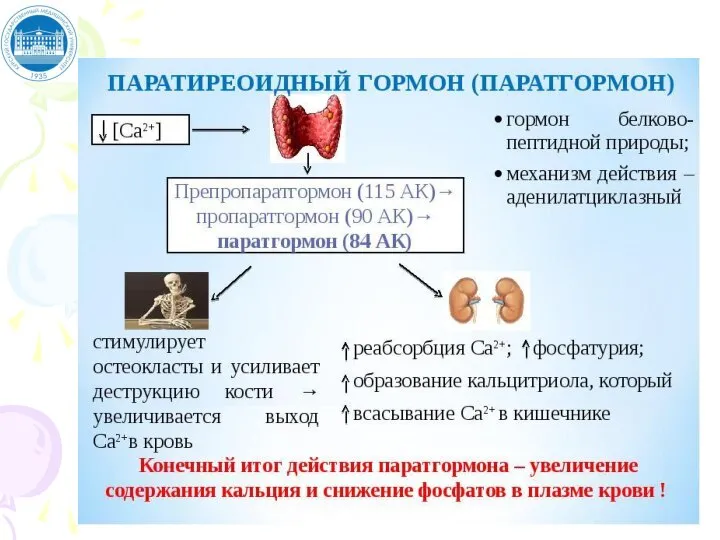
Слайд 66ХАРАКТЕРИСТИКА ПАРАТГОРМОНА
ПТГ – одноцепочечный полипептид, 84 АМК. Действие ПТГ:
1. Повышение [Ca2+];
2. Снижение
ХАРАКТЕРИСТИКА ПАРАТГОРМОНА
ПТГ – одноцепочечный полипептид, 84 АМК. Действие ПТГ:
1. Повышение [Ca2+];
2. Снижение

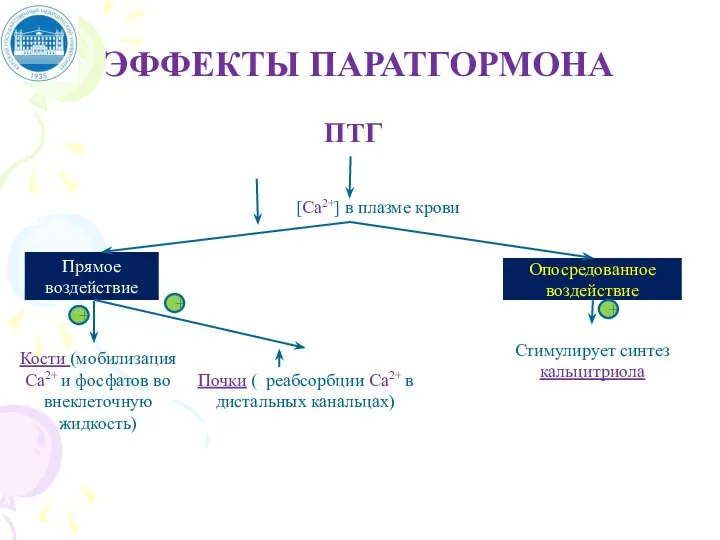
Слайд 67ЭФФЕКТЫ ПАРАТГОРМОНА
ЭФФЕКТЫ ПАРАТГОРМОНА
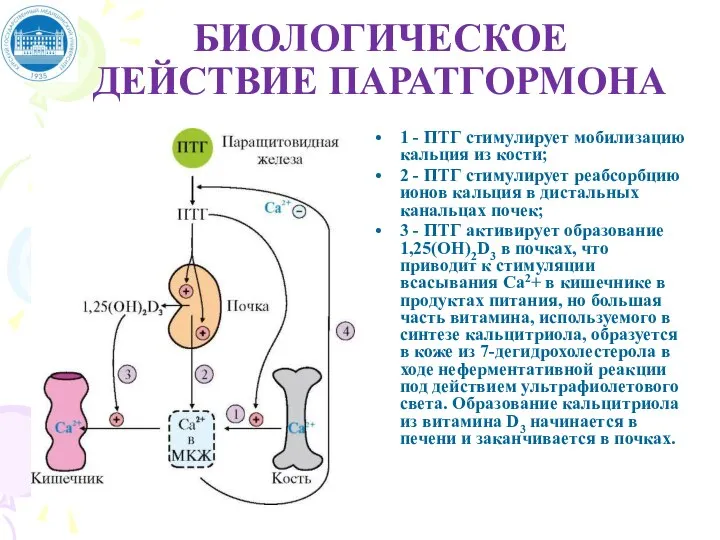
Слайд 68БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ПАРАТГОРМОНА
1 - ПТГ стимулирует мобилизацию кальция из кости;
2 -
БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ПАРАТГОРМОНА
1 - ПТГ стимулирует мобилизацию кальция из кости;
2 -
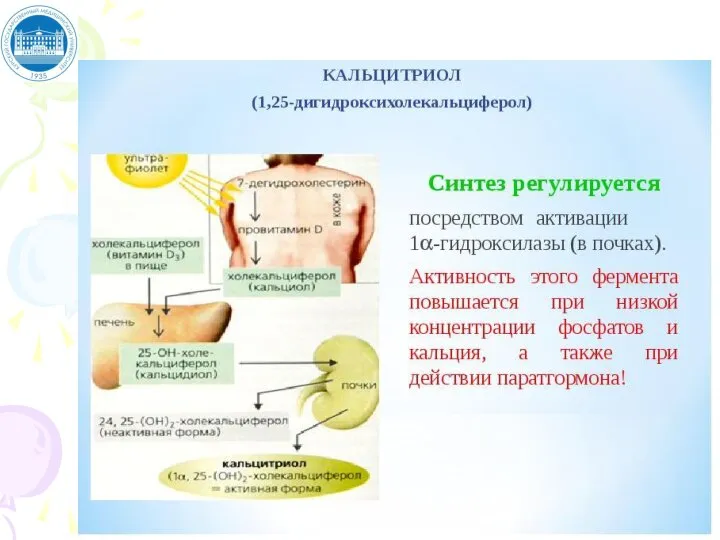
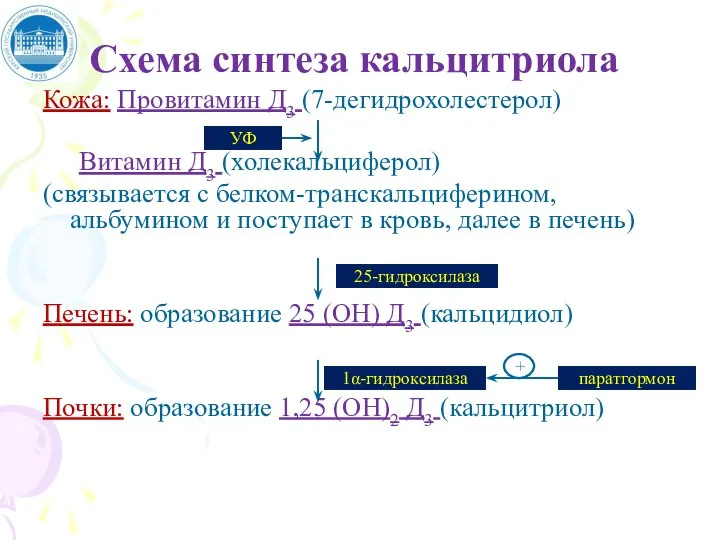
Слайд 70Схема синтеза кальцитриола
Кожа: Провитамин Д3 (7-дегидрохолестерол)
Витамин Д3 (холекальциферол)
(связывается с белком-транскальциферином, альбумином
Схема синтеза кальцитриола
Кожа: Провитамин Д3 (7-дегидрохолестерол)
Витамин Д3 (холекальциферол)
(связывается с белком-транскальциферином, альбумином
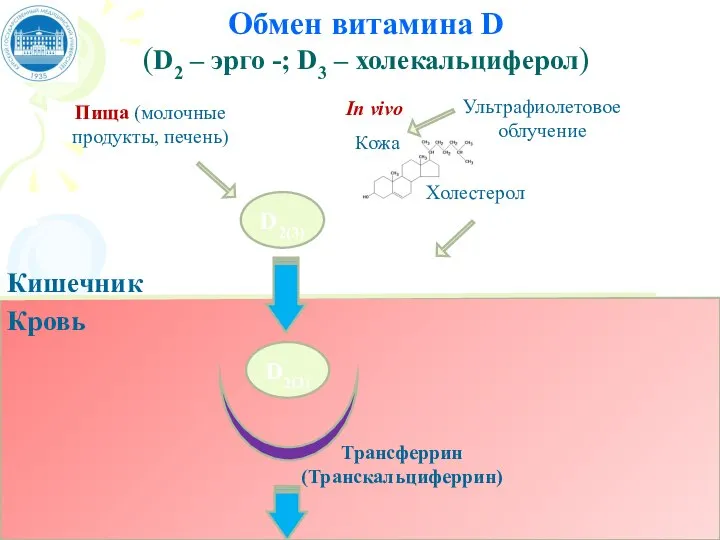
Слайд 71Обмен витамина D
(D2 – эрго -; D3 – холекальциферол)
Пища (молочные продукты,
Обмен витамина D
(D2 – эрго -; D3 – холекальциферол)
Пища (молочные продукты,
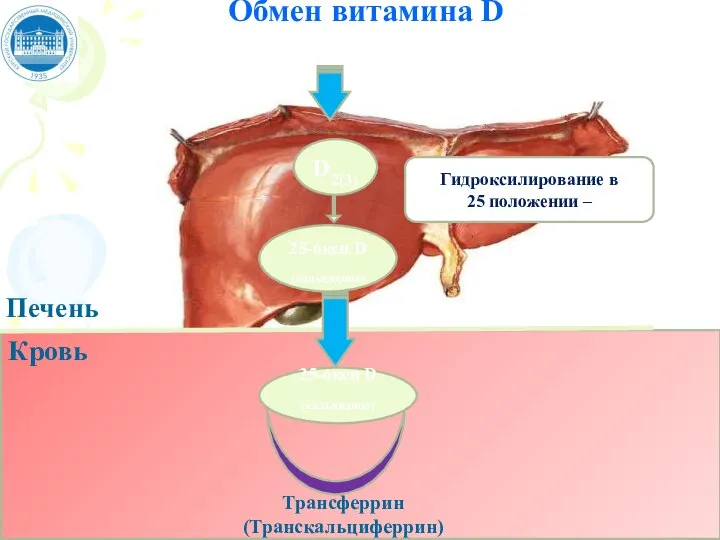
Слайд 72Обмен витамина D
Трансферрин (Транскальциферрин)
Печень
D2(3)
Кровь
25-окси D (кальцидиол)
25-окси D
(кальцидиол)
Гидроксилирование в
25 положении –
Обмен витамина D
Трансферрин (Транскальциферрин)
Печень
D2(3)
Кровь
25-окси D (кальцидиол)
25-окси D
(кальцидиол)
Гидроксилирование в
25 положении –
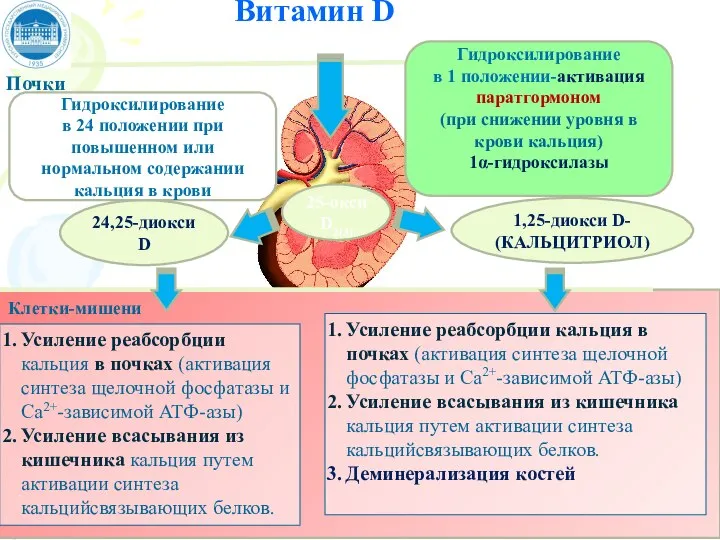
Слайд 73Витамин D
Почки
Клетки-мишени
25-окси D2(3)
Гидроксилирование
в 1 положении-активация паратгормоном
(при снижении уровня в
Витамин D
Почки
Клетки-мишени
25-окси D2(3)
Гидроксилирование
в 1 положении-активация паратгормоном
(при снижении уровня в
Слайд 74ЭФФЕКТЫ КАЛЬЦИТРИОЛА
[1,25 (ОН)2 Д3]
Клетки кишечника:
Индуцируют синтез Ca2+ - переносящих белков, которые
ЭФФЕКТЫ КАЛЬЦИТРИОЛА
[1,25 (ОН)2 Д3]
Клетки кишечника:
Индуцируют синтез Ca2+ - переносящих белков, которые
![ЭФФЕКТЫ КАЛЬЦИТРИОЛА [1,25 (ОН)2 Д3] Клетки кишечника: Индуцируют синтез Ca2+ - переносящих](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1046461/slide-73.jpg)
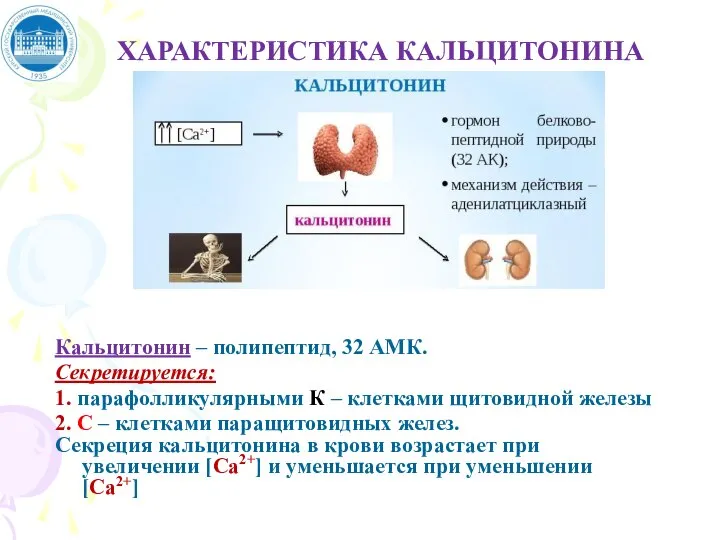
Слайд 75ХАРАКТЕРИСТИКА КАЛЬЦИТОНИНА
Кальцитонин – полипептид, 32 АМК.
Секретируется:
1. парафолликулярными К – клетками щитовидной железы
ХАРАКТЕРИСТИКА КАЛЬЦИТОНИНА
Кальцитонин – полипептид, 32 АМК.
Секретируется:
1. парафолликулярными К – клетками щитовидной железы
Слайд 76Ингибирует высвобождение Са2+ из костной ткани;
Стимулирует экскрецию Са2+ и фосфатов с мочой;
Снижение
Ингибирует высвобождение Са2+ из костной ткани;
Стимулирует экскрецию Са2+ и фосфатов с мочой;
Снижение


Слайд 78Гормональные нарушения обмена кальция
Гормональные нарушения обмена кальция


 В3 витамині суда, алкогольде, сірке қышқылында ерітілген майлы зат болып табылады
В3 витамині суда, алкогольде, сірке қышқылында ерітілген майлы зат болып табылады PSHE and Citizenship. Lesson 5
PSHE and Citizenship. Lesson 5 Значение грибов в круговороте веществ в природе
Значение грибов в круговороте веществ в природе Томский лес
Томский лес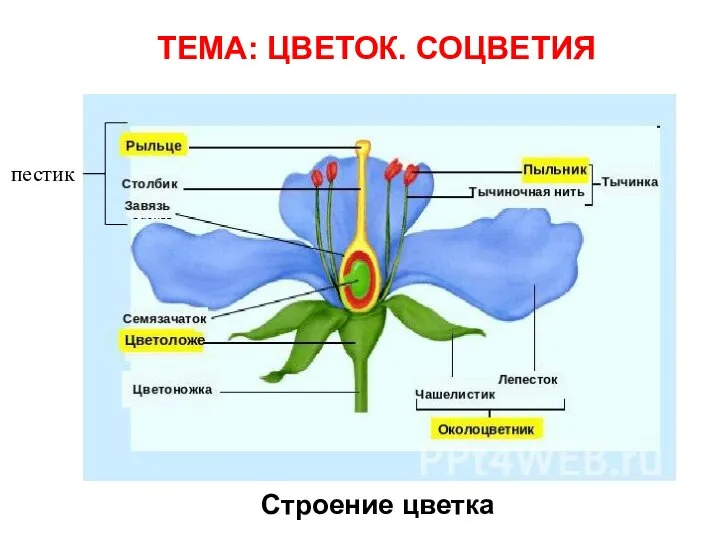 Цветок. Соцветия
Цветок. Соцветия Царство растений
Царство растений Современные представления о возникновении жизни на Земле
Современные представления о возникновении жизни на Земле Введение в остеологию кости туловища
Введение в остеологию кости туловища Палеозойская эра -570-230 млн л.н
Палеозойская эра -570-230 млн л.н Жизненные циклы растений
Жизненные циклы растений Строение мочевыделительной системы животных
Строение мочевыделительной системы животных Мир животных в природе
Мир животных в природе Живородящие птицы
Живородящие птицы Зоогеографическое районирование Мирового океана
Зоогеографическое районирование Мирового океана Иммунитет и иммунная система
Иммунитет и иммунная система Лишайники Мурманской области
Лишайники Мурманской области Пути и направления эволюции. 11 класс. 2
Пути и направления эволюции. 11 класс. 2 Внутренняя среда организма. 8 класс
Внутренняя среда организма. 8 класс Функции и виды тканей
Функции и виды тканей Проведение вирусологических методов исследования
Проведение вирусологических методов исследования Цепи питания
Цепи питания Деформирующий остеоартроз
Деформирующий остеоартроз 文:[美]史蒂夫・危金斯
文:[美]史蒂夫・危金斯 Энергообмен и его регуляция
Энергообмен и его регуляция Фенотипическая или модификационная изменчивость. Тест
Фенотипическая или модификационная изменчивость. Тест Координация и регуляция
Координация и регуляция Пептидний гормон секретин
Пептидний гормон секретин Презентация на тему Сосальщики и ленточные черви
Презентация на тему Сосальщики и ленточные черви