- Индивидуальная работа. Генетические коллекции гороха

Содержание
- 2. Горох (лат. Písum) — род травянистых растений семейства Бобовые (Fabaceae). Научная классификация Домен: Эукариоты Царство: Растения
- 3. Ботаническое описание Виды рода Горох — однолетние травы со слабыми вьющимися стеблями. Листья перистые и заканчиваются
- 4. Горох – самый распространенный из всех бобовых. Все сорта делятся на две группы: сахарные и лущильные.
- 5. В настоящее время в мировых генбанках продовольственных зерновых бобовых культур сохраняется более 900 тыс. образцов или
- 6. Различают 3 типа коллекций культурных растений и их диких сородичей Базовые коллекции - стратегические, длительно хранящиеся
- 7. Ко-коллекции (core collections) созданы для улучшенной оценки пользовательных коллекций, которые включают только ограниченное число образцов. В
- 8. С точки зрения селекционера генетические ресурсы можно разбить на 3 типа: а) Ресурсы генбанков. Этот тип
- 9. Схема передачи селекционерам исходного материала гороха во Франции На схеме приведен пример организации помощи селекционерам по
- 10. Ежегодно пополняется генофонд Национального центра генетических ресурсов растений Украины, созданный в 1995 году при институте растениеводства
- 11. Производство гороха в мире постоянно растет. "Горохова революция", которая состоялась в ряде стран Европейского Союза, в
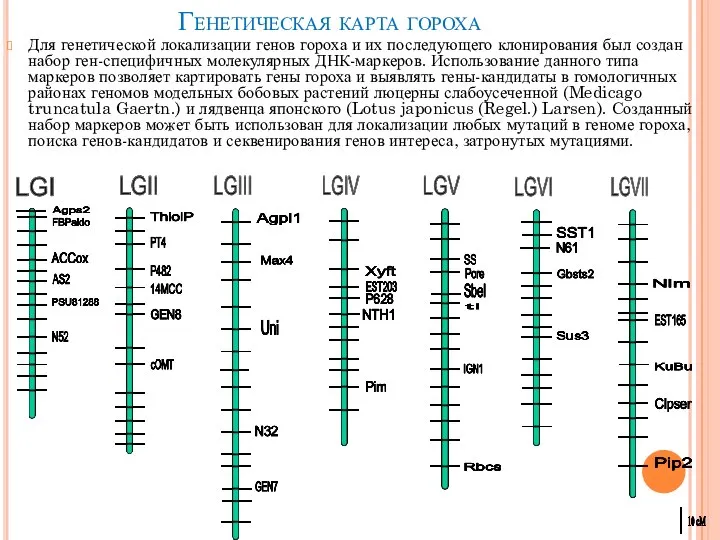
- 12. Генетическая карта гороха Для генетической локализации генов гороха и их последующего клонирования был создан набор ген-специфичных
- 13. Коллекции мутантов гороха
- 16. Роль гена Pssym40 в регуляции развития арбускулярной микоризы и его взаимодействие с другими регуляторными генами гороха
- 17. Проведены скрещивания новой серии симбиотических мутантов с исходной линией SGE и проанализированы по фенотипу потомство F1
- 18. В результате анализа была предположена относительная независимость выполнения генетических программ инфекции и развития тканей клубенька при
- 19. У гороха посевного описан мутант SGECdt (cdt), характеризующийся повышенным уровнем накопления кадмия и устойчивостью к данному
- 20. Мутация у гороха Pisum L. в локусе Тl Получен новый мутант гороха с многократно непарноперистым типом
- 30. Скачать презентацию
Слайд 2Горох (лат. Písum) — род травянистых растений семейства Бобовые (Fabaceae).
Научная классификация
Домен: Эукариоты
Царство: Растения
Отдел: Цветковые
Класс: Двудольные
Порядок: Бобовоцветные
Семейство: Бобовые
Род: Горох
Род включает семь видов:
Pisum abyssinicum A.Braun
Pisum ensifolium (Lapeyr.) E.H.L.Krause
Pisum fulvum Sibth. & Sm.
Pisum
Горох (лат. Písum) — род травянистых растений семейства Бобовые (Fabaceae).
Научная классификация
Домен: Эукариоты
Царство: Растения
Отдел: Цветковые
Класс: Двудольные
Порядок: Бобовоцветные
Семейство: Бобовые
Род: Горох
Род включает семь видов:
Pisum abyssinicum A.Braun
Pisum ensifolium (Lapeyr.) E.H.L.Krause
Pisum fulvum Sibth. & Sm.
Pisum

Слайд 3Ботаническое описание
Виды рода Горох — однолетние травы со слабыми вьющимися стеблями.
Листья перистые и заканчиваются ветвистыми усиками, с
Ботаническое описание
Виды рода Горох — однолетние травы со слабыми вьющимися стеблями.
Листья перистые и заканчиваются ветвистыми усиками, с

Слайд 4Горох – самый распространенный из всех бобовых. Все сорта делятся на две
Горох – самый распространенный из всех бобовых. Все сорта делятся на две

Слайд 5В настоящее время в мировых генбанках продовольственных зерновых бобовых культур сохраняется более 900
В настоящее время в мировых генбанках продовольственных зерновых бобовых культур сохраняется более 900

Слайд 6Различают 3 типа коллекций культурных растений и их диких сородичей
Базовые коллекции -
Различают 3 типа коллекций культурных растений и их диких сородичей
Базовые коллекции -

Слайд 7Ко-коллекции
(core collections) созданы для улучшенной оценки пользовательных коллекций, которые включают только
Ко-коллекции
(core collections) созданы для улучшенной оценки пользовательных коллекций, которые включают только

Слайд 8С точки зрения селекционера генетические ресурсы можно разбить на 3 типа:
а) Ресурсы генбанков. Этот
С точки зрения селекционера генетические ресурсы можно разбить на 3 типа:
а) Ресурсы генбанков. Этот

Слайд 9Схема передачи селекционерам исходного материала гороха во Франции
На схеме приведен пример организации
Схема передачи селекционерам исходного материала гороха во Франции
На схеме приведен пример организации
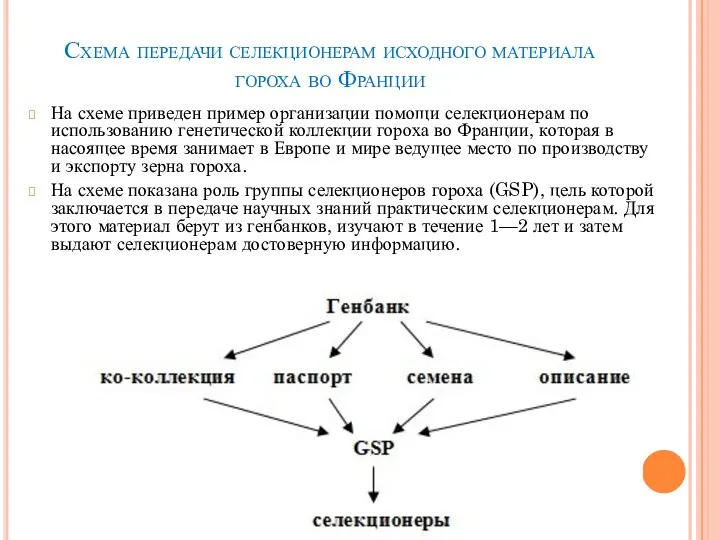
Слайд 10Ежегодно пополняется генофонд Национального центра генетических ресурсов растений Украины, созданный в 1995 году при институте
Ежегодно пополняется генофонд Национального центра генетических ресурсов растений Украины, созданный в 1995 году при институте

Слайд 11Производство гороха в мире постоянно растет. "Горохова революция", которая состоялась в ряде
Производство гороха в мире постоянно растет. "Горохова революция", которая состоялась в ряде

Слайд 12Генетическая карта гороха
Для генетической локализации генов гороха и их последующего клонирования был
Генетическая карта гороха
Для генетической локализации генов гороха и их последующего клонирования был
Слайд 13Коллекции мутантов гороха
Коллекции мутантов гороха
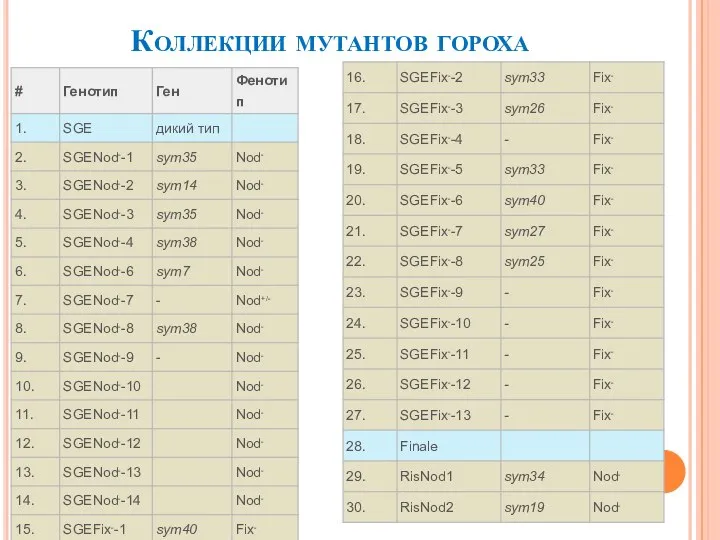

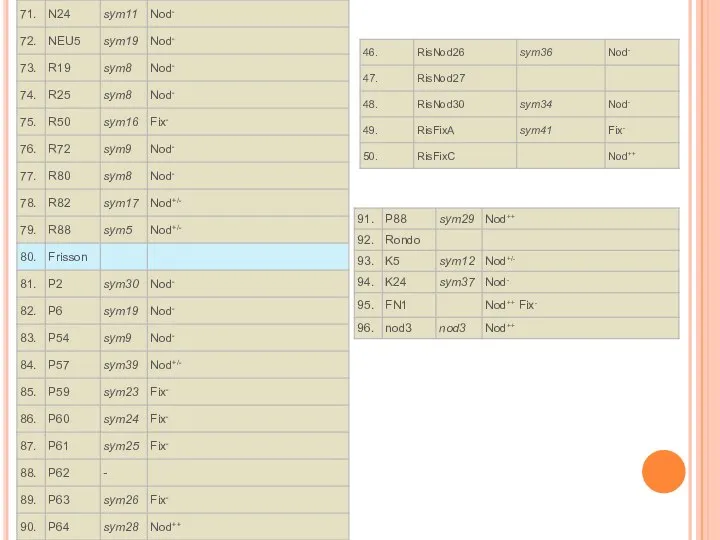
Слайд 16Роль гена Pssym40 в регуляции развития арбускулярной микоризы и его взаимодействие с
Роль гена Pssym40 в регуляции развития арбускулярной микоризы и его взаимодействие с

Слайд 17Проведены скрещивания новой серии симбиотических мутантов с исходной линией SGE и проанализированы
Проведены скрещивания новой серии симбиотических мутантов с исходной линией SGE и проанализированы

Слайд 18В результате анализа была предположена относительная независимость выполнения генетических программ инфекции и
В результате анализа была предположена относительная независимость выполнения генетических программ инфекции и

Слайд 19У гороха посевного описан мутант SGECdt (cdt), характеризующийся повышенным уровнем накопления кадмия
У гороха посевного описан мутант SGECdt (cdt), характеризующийся повышенным уровнем накопления кадмия

Слайд 20Мутация у гороха Pisum L. в локусе Тl
Получен новый мутант гороха с
Мутация у гороха Pisum L. в локусе Тl
Получен новый мутант гороха с









 Пернатые друзья
Пернатые друзья Типы питания живых организмов. 6 класс
Типы питания живых организмов. 6 класс Вегетативная нервная система
Вегетативная нервная система Размножение и развитие организмов
Размножение и развитие организмов Сцепленное наследование генов. ЕГЭ по биологии, 28 задача
Сцепленное наследование генов. ЕГЭ по биологии, 28 задача Центры происхождения культурных растений и домашних животных
Центры происхождения культурных растений и домашних животных Размножение. Уровни размножения
Размножение. Уровни размножения Как зимой помочь птицам
Как зимой помочь птицам Происхождение человека, антропогенез
Происхождение человека, антропогенез Скелет
Скелет состав и строение белков
состав и строение белков Птицы. Специализация
Птицы. Специализация Презентация на тему СИМБИОЗ
Презентация на тему СИМБИОЗ  Строение тела человека
Строение тела человека Отбор и его значение в селекции и семеноводстве
Отбор и его значение в селекции и семеноводстве Самый-самый
Самый-самый Гидропоника в мире
Гидропоника в мире Презентация на тему Размножение земноводных
Презентация на тему Размножение земноводных  Возрастная физиология
Возрастная физиология Только самое главное и важное для подготовке к ЕГЭ по биологии
Только самое главное и важное для подготовке к ЕГЭ по биологии Самые необычные животные планеты: ангорский кролик
Самые необычные животные планеты: ангорский кролик Опорно-двигательная система. Скелет человека
Опорно-двигательная система. Скелет человека Картирование хромосом
Картирование хромосом Заповідники. Ботанічні сади
Заповідники. Ботанічні сади Мариукультура. Культивирование тихоокеанских устриц
Мариукультура. Культивирование тихоокеанских устриц Разделы зоологии
Разделы зоологии Физиология спорта. Особенности физиологии младших школьников
Физиология спорта. Особенности физиологии младших школьников пр. р. 6 базид
пр. р. 6 базид