- Калориметрические методы анализа биомолекул

Содержание
- 2. Синхронное изменение спектров триптофановой флуоресценции и кругового дихроизма при плавлении белка Изменение подвижности белка в акриламидном
- 3. Разница между «постепенным» переходом и переходом по принципу «все или ничего» отражается не в виде кривой
- 4. Холодовая денатурация
- 5. Структура белка после денатурации
- 6. Расплавленная глобула и её свойства
- 8. Почему белок плавится как единое целое?
- 9. Скорость сворачивания белка при его синтезе достаточно велика. Подобная самоорганизация белковых структур относится, с физической точки
- 10. Парадокс Левинталя «С одной стороны, нативная пространственная структура по всем тестам ведет себя как самая стабильная
- 11. Структура переходного состояния белка CheY
- 12. Клеточная машинерия, способствующая правильному сворачиванию белка Шапероны Пролилизомераза Дисульфидизомераза
- 13. Шапероны Бывают двух типов: Фолдазы (GroEL/GroES, DnaK/DnaG) Холдазы (HsP 33) Примеры эукариотических шаперонов: GRP78/BiP, GRP94, GRP170,
- 14. Механизм работы шаперона из класса фолдаз (на примере GroEL/GroES)
- 16. Механизм работы шаперона из класса холдаз (на примере DnaK)
- 17. Дифференциальная сканирующая калориметрия (устройство прибора)
- 18. Дифференциальная сканирующая калориметрия (принцип метода) ΔH=∫ΔCp (T)dT ΔS=∫ΔCp(T)/TdT ΔCp= Mw x 0.06/ Cm x Vc x
- 19. Кривые ДСК биомолекул зависят от: рН Скорости нагрева Природы растворителя Ионной силы раствора Концентрации вещества
- 21. Критерий Вант-Гоффа «Эффективная теплота" перехода, вычисляемая из его ширины совпадает с "калориметрической теплотой" этого перехода, т.е.
- 22. ΔHv-g= ΔHcal
- 23. Калориметрия мультидоменных белков
- 24. Определение стабильности белков с помощью ДСК
- 25. Что еще можно узнать из спектров ДСК белков?
- 26. Изотермическая калориметрия титрования (устройство прибора)
- 27. Изотермическая калориметрия титрования (принцип метода) ΔG = -RTlnKa = ΔH-TΔS
- 28. Изотермическая калориметрия титрования. Приложения метода Идентификация таргетного белка из смеси биомолекул Из кривой связывания мы можем
- 29. Изотермическая калориметрия титрования. Приложения метода Идентификация таргетного белка из смеси биомолекул
- 30. Дифференциальный термический анализ
- 32. Скачать презентацию
Слайд 2Синхронное изменение спектров триптофановой флуоресценции и кругового дихроизма при плавлении белка
Изменение подвижности
Синхронное изменение спектров триптофановой флуоресценции и кругового дихроизма при плавлении белка
Изменение подвижности
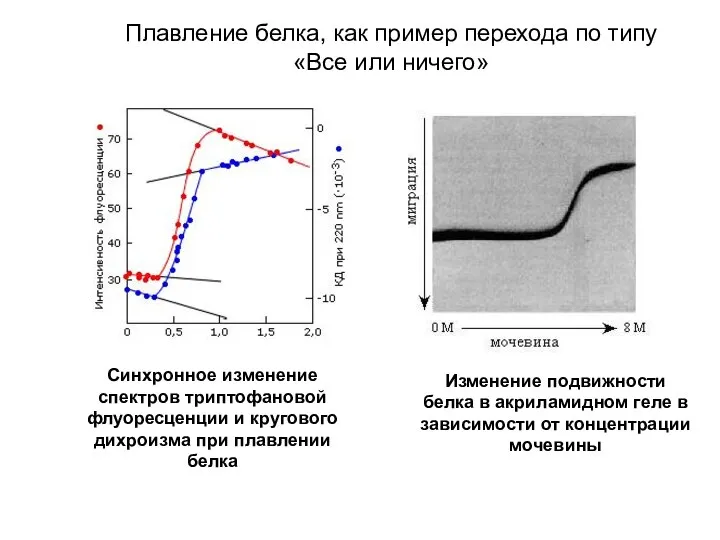
Слайд 3Разница между «постепенным» переходом и переходом по принципу «все или ничего» отражается
Разница между «постепенным» переходом и переходом по принципу «все или ничего» отражается
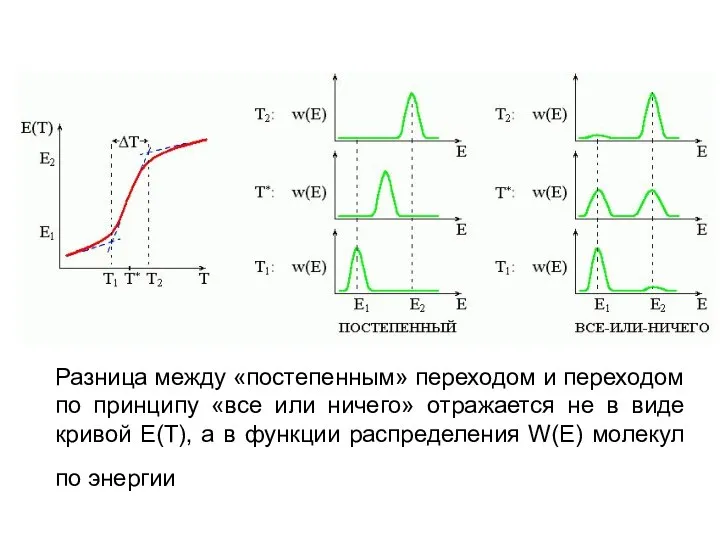
Слайд 4Холодовая денатурация
Холодовая денатурация
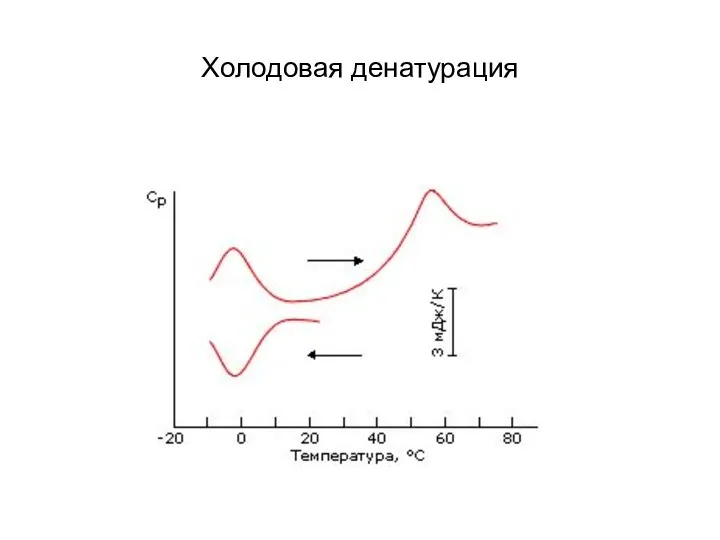
Слайд 5Структура белка после денатурации
Структура белка после денатурации
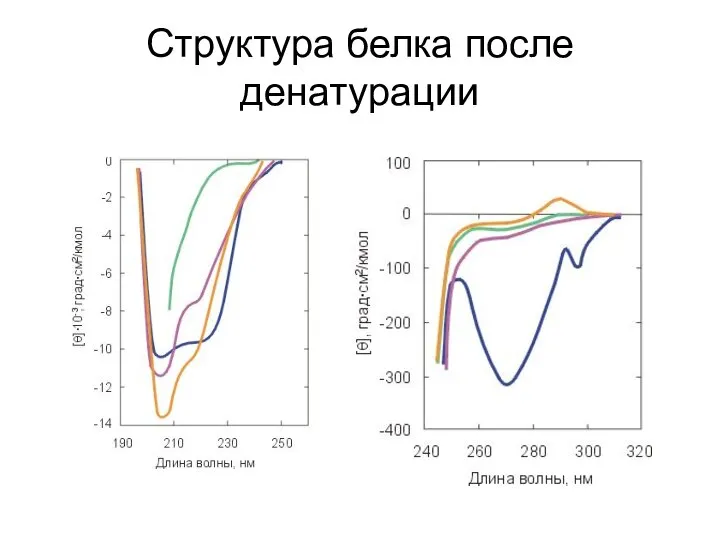
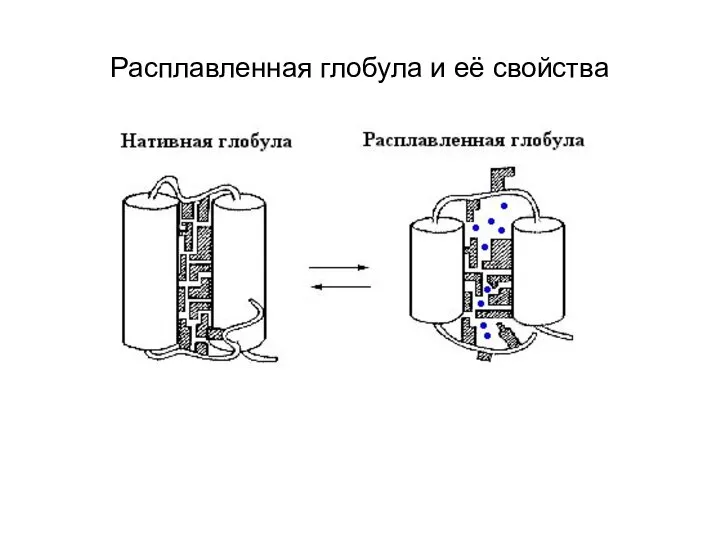
Слайд 6Расплавленная глобула и её свойства
Расплавленная глобула и её свойства

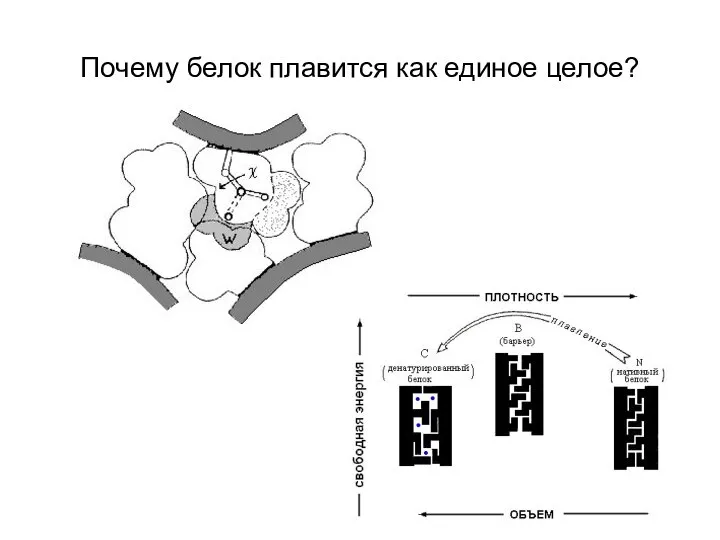
Слайд 8Почему белок плавится как единое целое?
Почему белок плавится как единое целое?
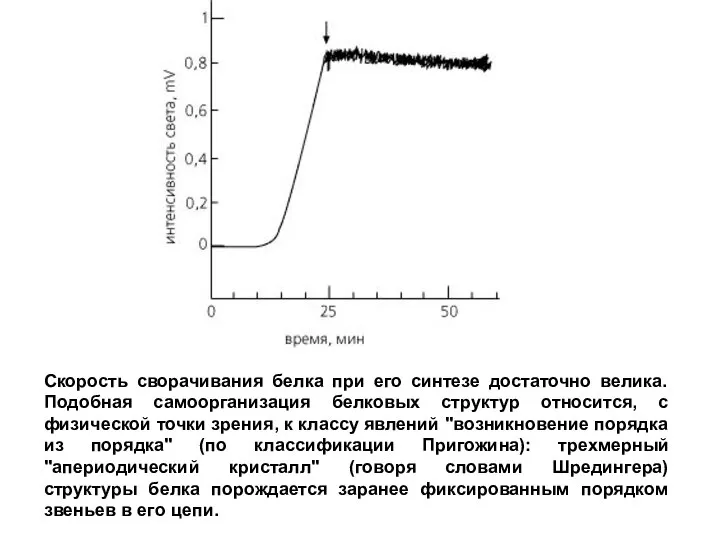
Слайд 9Скорость сворачивания белка при его синтезе достаточно велика. Подобная самоорганизация белковых структур
Скорость сворачивания белка при его синтезе достаточно велика. Подобная самоорганизация белковых структур
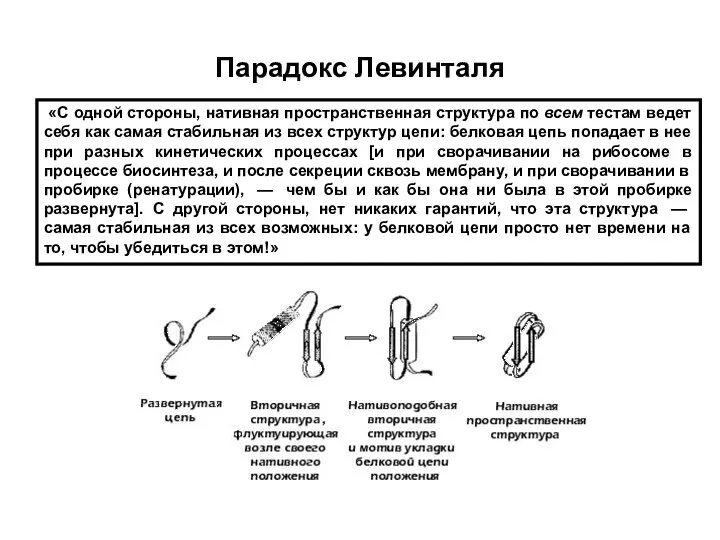
Слайд 10Парадокс Левинталя
«С одной стороны, нативная пространственная структура по всем тестам ведет себя как
Парадокс Левинталя
«С одной стороны, нативная пространственная структура по всем тестам ведет себя как
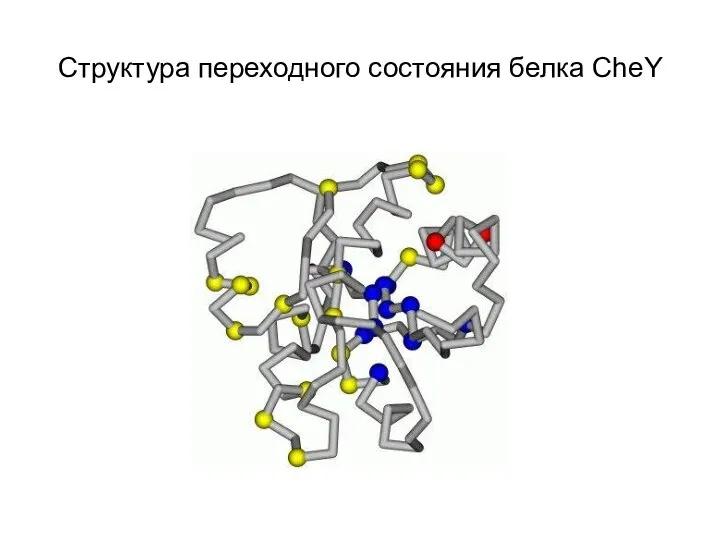
Слайд 11Структура переходного состояния белка CheY
Структура переходного состояния белка CheY
Слайд 12Клеточная машинерия, способствующая правильному сворачиванию белка
Шапероны
Пролилизомераза
Дисульфидизомераза
Клеточная машинерия, способствующая правильному сворачиванию белка
Шапероны
Пролилизомераза
Дисульфидизомераза

Слайд 13Шапероны
Бывают двух типов:
Фолдазы (GroEL/GroES, DnaK/DnaG)
Холдазы (HsP 33)
Примеры эукариотических шаперонов:
GRP78/BiP, GRP94, GRP170, кальнексин,
Шапероны
Бывают двух типов:
Фолдазы (GroEL/GroES, DnaK/DnaG)
Холдазы (HsP 33)
Примеры эукариотических шаперонов:
GRP78/BiP, GRP94, GRP170, кальнексин,

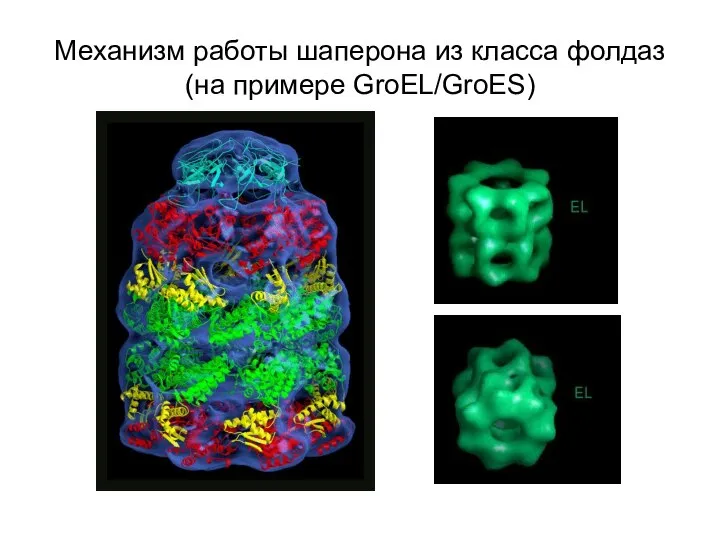
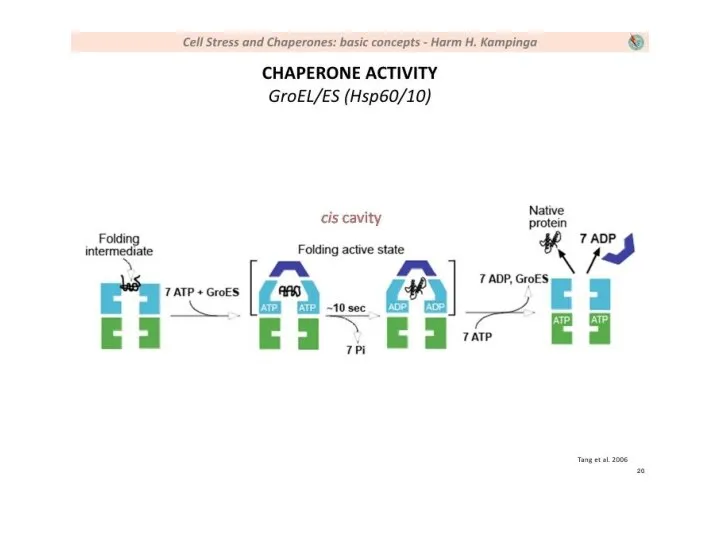
Слайд 14Механизм работы шаперона из класса фолдаз (на примере GroEL/GroES)
Механизм работы шаперона из класса фолдаз (на примере GroEL/GroES)
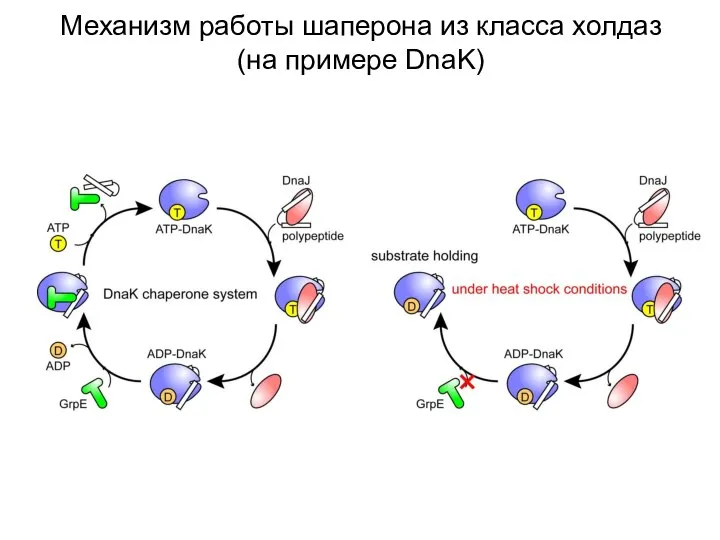
Слайд 16Механизм работы шаперона из класса холдаз (на примере DnaK)
Механизм работы шаперона из класса холдаз (на примере DnaK)
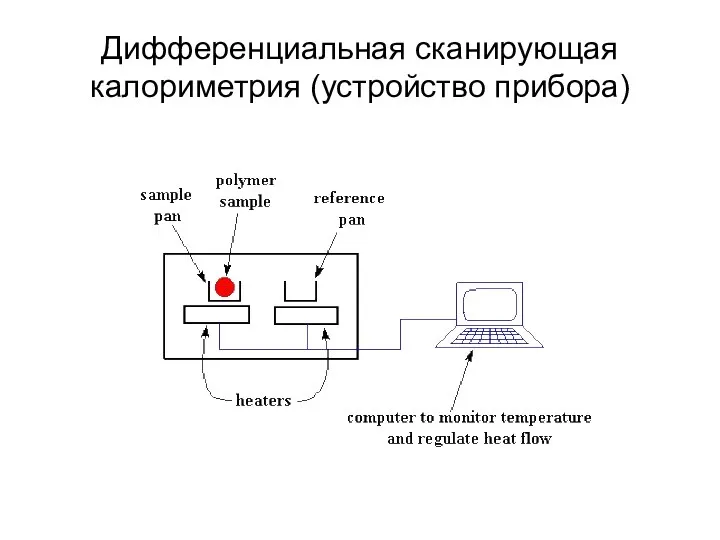
Слайд 17Дифференциальная сканирующая калориметрия (устройство прибора)
Дифференциальная сканирующая калориметрия (устройство прибора)
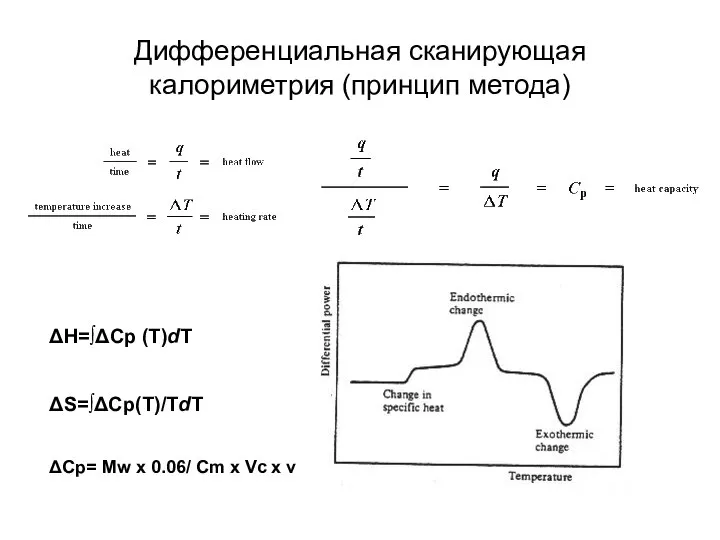
Слайд 18Дифференциальная сканирующая калориметрия (принцип метода)
ΔH=∫ΔCp (T)dT
ΔS=∫ΔCp(T)/TdT
ΔCp= Mw x 0.06/ Cm x Vc
Дифференциальная сканирующая калориметрия (принцип метода)
ΔH=∫ΔCp (T)dT
ΔS=∫ΔCp(T)/TdT
ΔCp= Mw x 0.06/ Cm x Vc
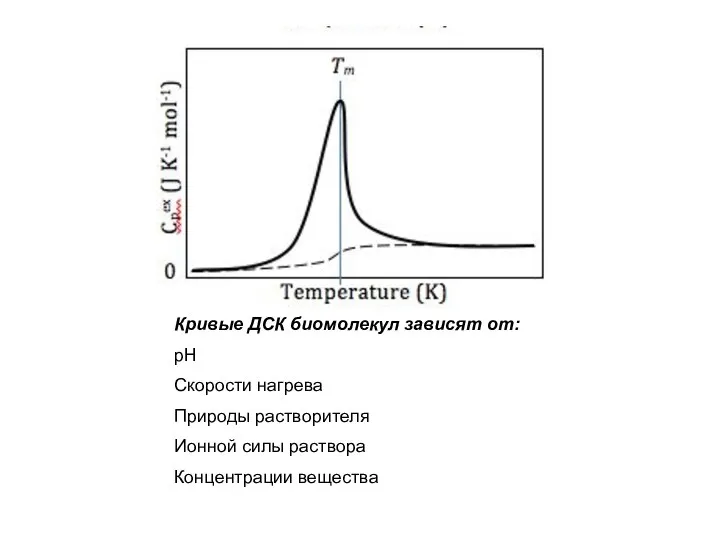
Слайд 19Кривые ДСК биомолекул зависят от:
рН
Скорости нагрева
Природы растворителя
Ионной силы раствора
Концентрации вещества
Кривые ДСК биомолекул зависят от:
рН
Скорости нагрева
Природы растворителя
Ионной силы раствора
Концентрации вещества
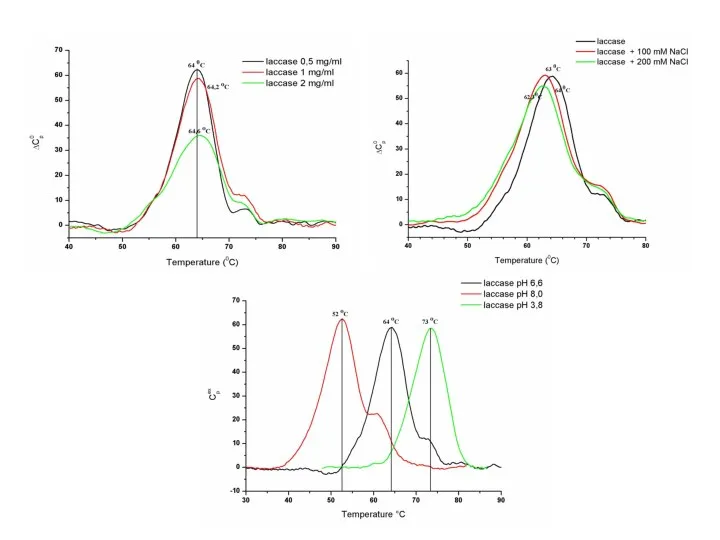
Слайд 21Критерий Вант-Гоффа
«Эффективная теплота" перехода, вычисляемая из его ширины совпадает с "калориметрической теплотой"
Критерий Вант-Гоффа
«Эффективная теплота" перехода, вычисляемая из его ширины совпадает с "калориметрической теплотой"

Слайд 22ΔHv-g= ΔHcal
ΔHv-g= ΔHcal
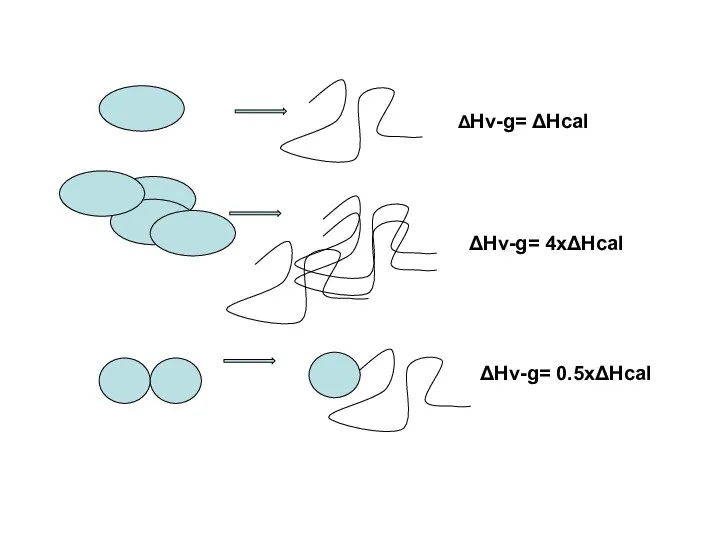
Слайд 23Калориметрия мультидоменных белков
Калориметрия мультидоменных белков
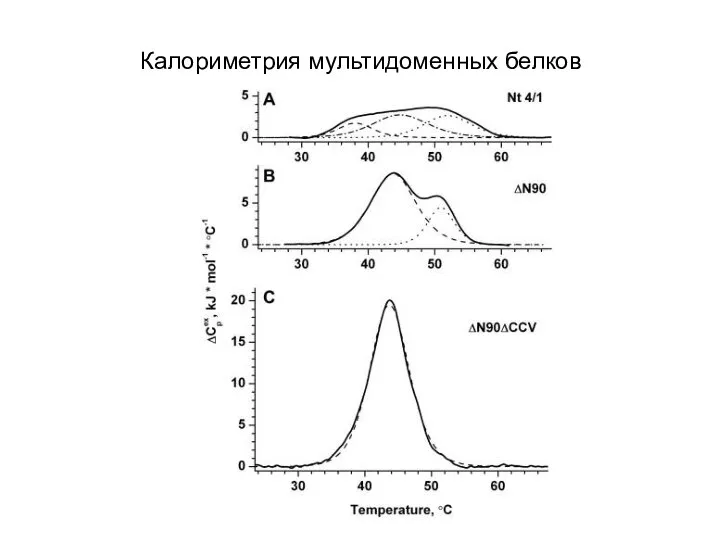
Слайд 24Определение стабильности белков с помощью ДСК
Определение стабильности белков с помощью ДСК
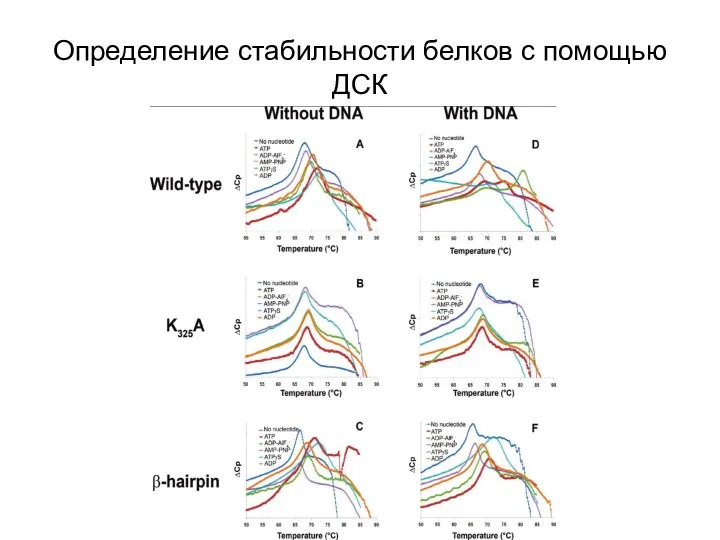
Слайд 25Что еще можно узнать из спектров ДСК белков?
Что еще можно узнать из спектров ДСК белков?
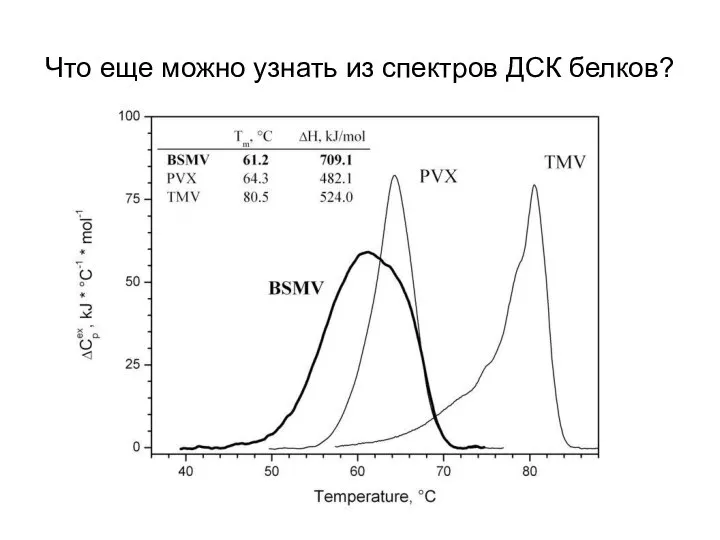
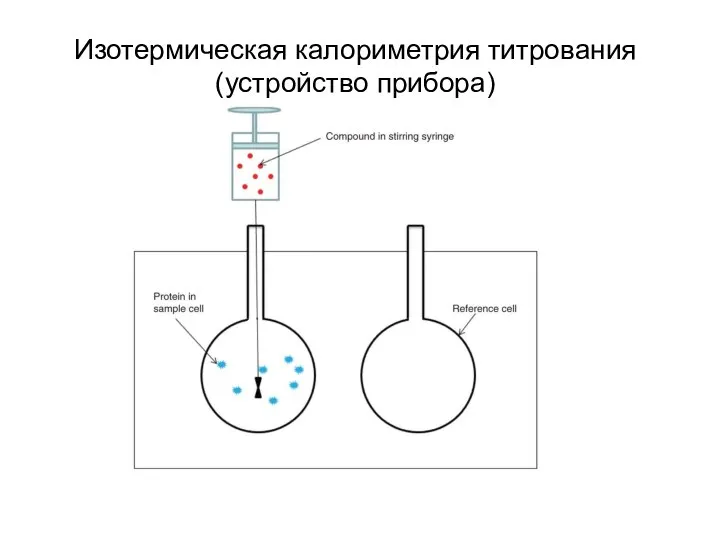
Слайд 26Изотермическая калориметрия титрования (устройство прибора)
Изотермическая калориметрия титрования (устройство прибора)
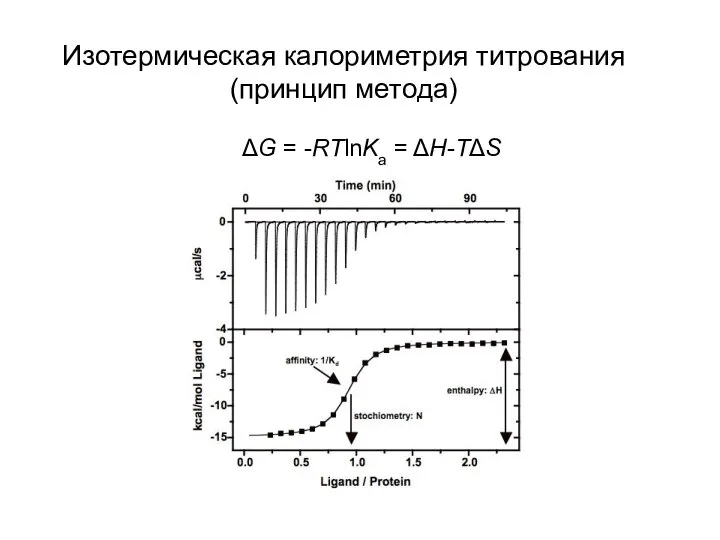
Слайд 27Изотермическая калориметрия титрования (принцип метода)
ΔG = -RTlnKa = ΔH-TΔS
Изотермическая калориметрия титрования (принцип метода)
ΔG = -RTlnKa = ΔH-TΔS
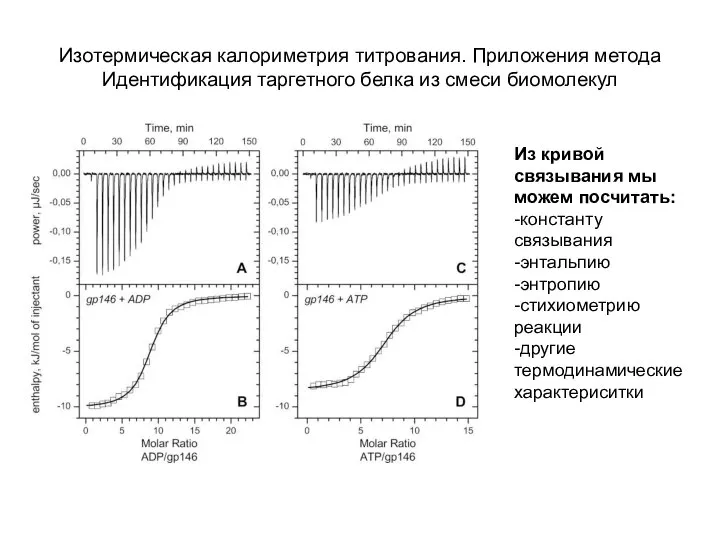
Слайд 28Изотермическая калориметрия титрования. Приложения метода
Идентификация таргетного белка из смеси биомолекул
Из кривой связывания
Изотермическая калориметрия титрования. Приложения метода
Идентификация таргетного белка из смеси биомолекул
Из кривой связывания
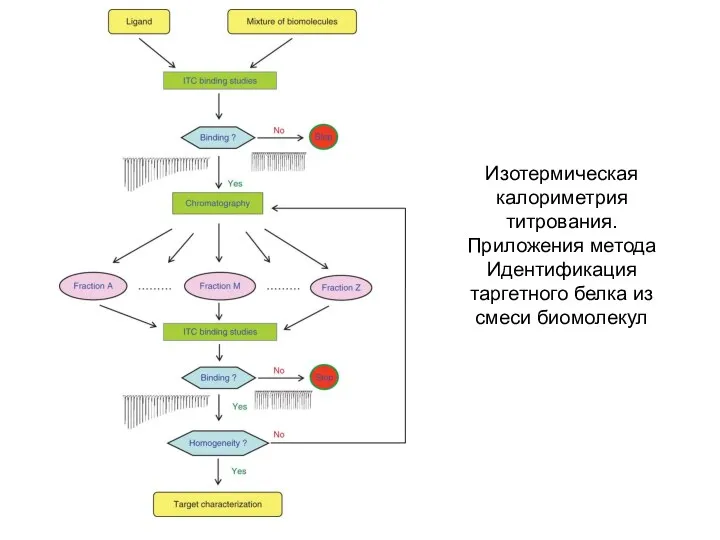
Слайд 29Изотермическая калориметрия титрования. Приложения метода
Идентификация таргетного белка из смеси биомолекул
Изотермическая калориметрия титрования. Приложения метода
Идентификация таргетного белка из смеси биомолекул
Слайд 30Дифференциальный термический анализ
Дифференциальный термический анализ
 Анатомия сердца
Анатомия сердца Растения Австралии
Растения Австралии Камыш - королевская власть, царственность
Камыш - королевская власть, царственность Классификация перцептивных процессов. Интермодальные ощущения и синестезии
Классификация перцептивных процессов. Интермодальные ощущения и синестезии Penguins
Penguins Великолепная изоляция
Великолепная изоляция Презентация на тему Видоизменения корней
Презентация на тему Видоизменения корней  Надежная защита организма
Надежная защита организма Удивительные растения
Удивительные растения Стебель - осевая часть побега. Лекция 2
Стебель - осевая часть побега. Лекция 2 Организм человека
Организм человека Метаболизм. Энергообмен
Метаболизм. Энергообмен genetika
genetika Можно ли жить и не дышать
Можно ли жить и не дышать Ядро клетки. Хромосомы
Ядро клетки. Хромосомы Лиственница
Лиственница Zheludochno-kishechny_trakt_Prezentatsia_2
Zheludochno-kishechny_trakt_Prezentatsia_2 Презентация на тему Готовимся к ЕГЭ по биологии. – практикум по подготовке к успешному выполнению задания С
Презентация на тему Готовимся к ЕГЭ по биологии. – практикум по подготовке к успешному выполнению задания С  Бактерии. Разновидности бактерий
Бактерии. Разновидности бактерий Физиология питания
Физиология питания Мультиклеточный глиальный ответ
Мультиклеточный глиальный ответ Природные сообщества
Природные сообщества Актуальные и перспективные медицинские технологии
Актуальные и перспективные медицинские технологии Биохимия мышечной ткани
Биохимия мышечной ткани Презентация по биологии_ _Движение живых организмов_
Презентация по биологии_ _Движение живых организмов_ Brain and mind
Brain and mind 5.Нервная система. ЦНС
5.Нервная система. ЦНС Сердечно-сосудистая система человека. (Лекция 2)
Сердечно-сосудистая система человека. (Лекция 2)