- Защитные системы эукариот

Содержание
- 3. РНК-интерференция у эукариот Эндрю Файер и Крейг Мелло, открыли явление в 1998 году, получили Нобелевскую премию
- 4. Механизмы подавления экспрессии генов с помощью двуцепочечной РНК (РНК интерференция) цитоплазма ядро Dicer расщепление и деградация
- 5. Гидом для pAgo является однонитевая ДНК, мишенью – иногда РНК, иногда ДНК Pyrococcus furiosus Aquifex aeolicus
- 6. Домены в составе pAgo pAgo есть у 30% архей и 10% эубактерий,
- 7. РНК-интерференция возникла за счёт объединения эубактериальных и архейных компонентов Dicer
- 8. Жизненный цикл амебозойного протиста Dictyostelium
- 9. Не все бактерии одинаково полезны (и питательны)
- 10. Механизмы передачи сигнала у слизевика
- 11. S-клетки (sentinel cells) отвечают за фагоцитоз бактерий в плодовом теле
- 12. Наличие «барьера Вейсмана» создает эволюционные предпосылки для его нарушения
- 13. Амебы – обманщики в химерных колониях не хотят образовывать ножку, и избегают ее зачатка
- 14. Плодовые тела диктиостелиума сильно структурированы и видоспецифичны, межвидовая агрегация отсутствует
- 15. Задача гифы – обеспечить рост апикальной клетки
- 16. Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее делают более прочной, сохраняют ее единство при
- 17. При слиянии гиф двух или более контактирующих колоний происходит объединение в одной клетке нескольких генетически различных
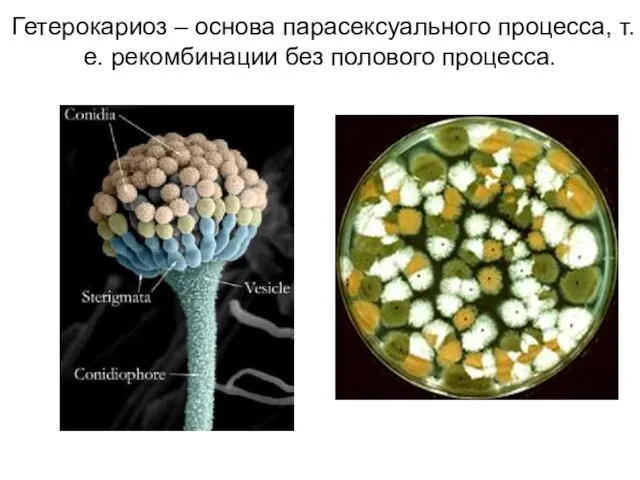
- 18. Гетерокариоз – основа парасексуального процесса, т.е. рекомбинации без полового процесса.
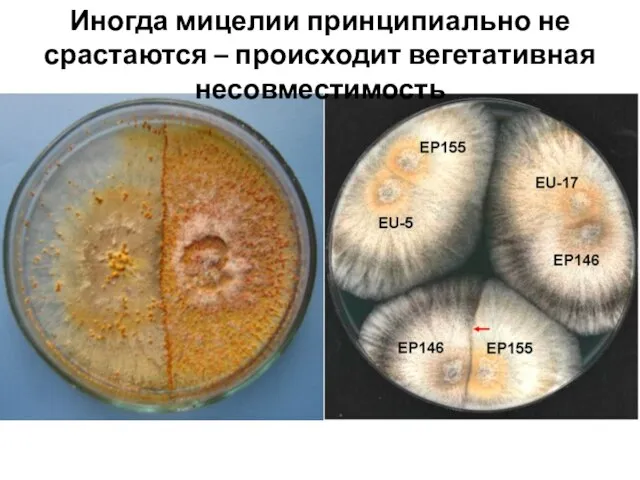
- 19. Иногда мицелии принципиально не срастаются – происходит вегетативная несовместимость
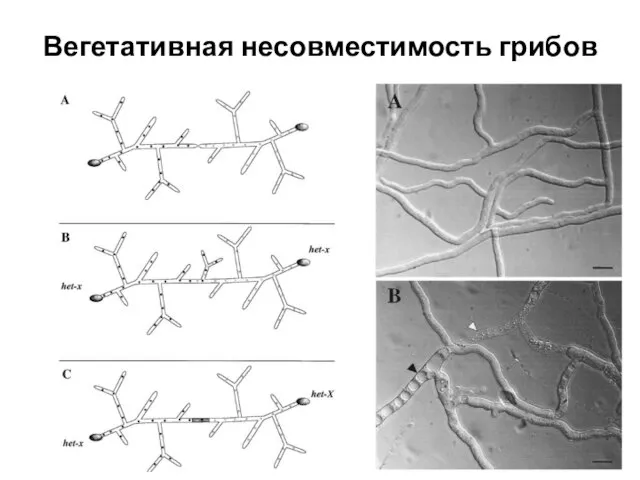
- 20. Вегетативная несовместимость грибов
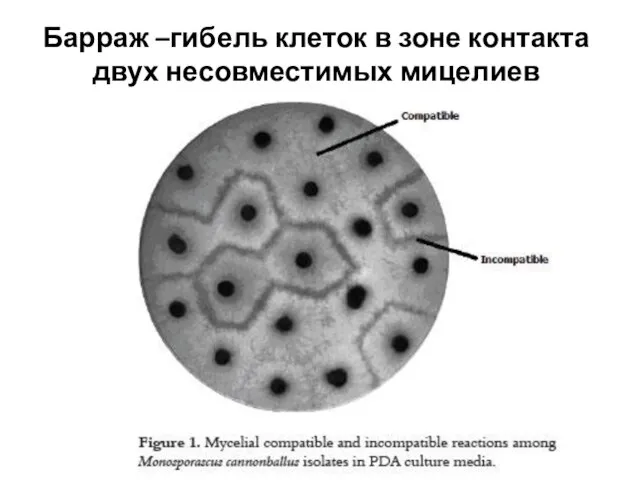
- 21. Барраж –гибель клеток в зоне контакта двух несовместимых мицелиев
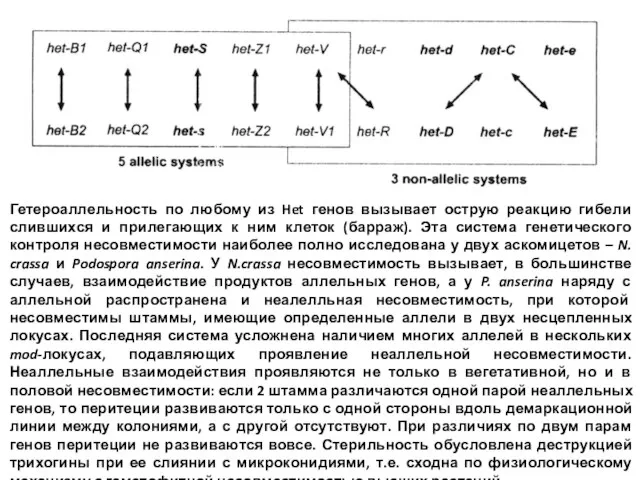
- 22. Гетероаллельность по любому из Het генов вызывает острую реакцию гибели слившихся и прилегающих к ним клеток
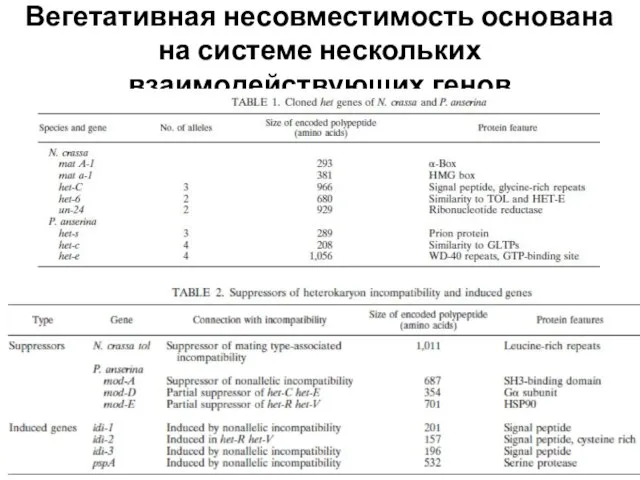
- 23. Вегетативная несовместимость основана на системе нескольких взаимодействующих генов
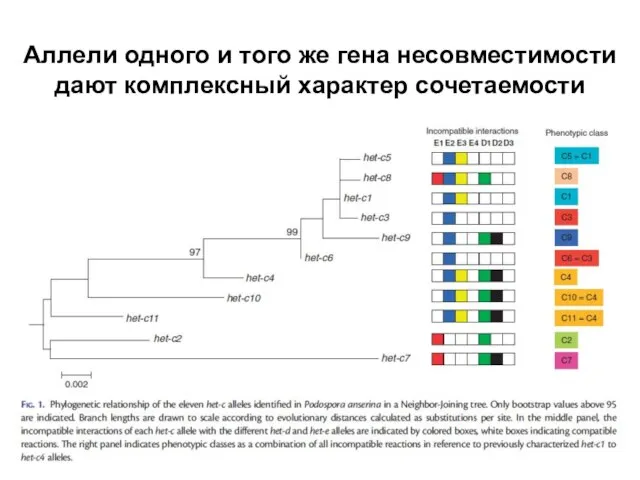
- 24. Аллели одного и того же гена несовместимости дают комплексный характер сочетаемости
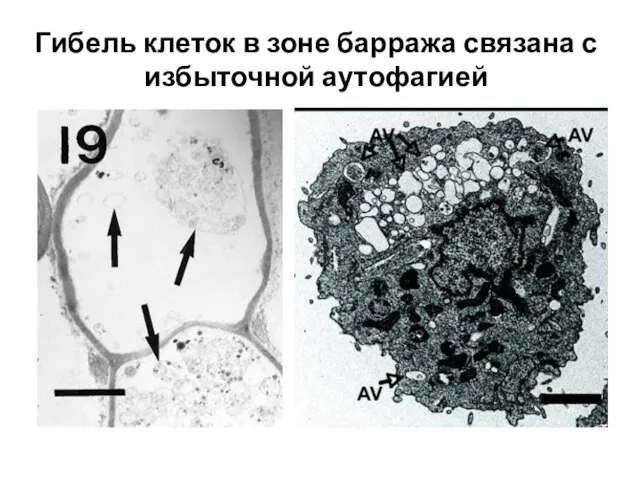
- 25. Гибель клеток в зоне барража связана с избыточной аутофагией
- 26. Преимущества от вегетативной несовместимости грибов Защита от эгоистичных паразитов, преодолевающих барьер Вейсмана Защита от вирусов, передаваемых
- 27. О Р Ф Уровни иммунитета растения
- 28. ИММУНИТЕТ НА УРОВНЕ ЦЕЛОГО ОРГАНИЗМА (первая линия обороны) Цель – не допустить патогенных организмов к чувствительным
- 29. Фитоиммунитет: распознавание элиситоров
- 30. Фитоиммунитет – распознавание PAMP (PTI) и эффекторных белков (ETI)
- 31. PAMP = неспецифические элиситоры
- 33. Важно детектировать не только грибы и бактерии, но и фитофагов
- 34. КАК РАСТЕНИЯ СПРАВЛЯЮТСЯ С ВИРУЛЕНТНЫМИ ПАРАЗИТАМИ (ТРЕТЬЯ ЛИНИЯ ОБОРОНЫ) У растений возникла система ген-на ген, в
- 35. Белки с лейцин-богатыми повторами –основа и ETI, и PTI
- 36. R-белки – рецепторы с лейцин-богатыми повторами
- 37. «Игра в пятнашки» между патогеном и растением
- 38. Фитоантиципины – выделяются конститутивно (то есть постоянно) Фитоалексины –выделяются в ответ на появление фитофага или патогена
- 39. Эффекторные белки фитоиммунитета Дефензины – низкомолекулярные (45-54 аминокислот) белки, богатые цистеином. Продуцируются млекопитающими, грибами, насекомыми и
- 40. Реакция сверхчувствительности растений
- 41. Реакция сверхчувствительности и клеточная гибель
- 42. Механизмы реакции сверхчувствительности
- 43. РНК-интерференция против вирусов и вироидов
- 44. микроРНК как переключатель типов иммунного ответа у растений
- 46. Скачать презентацию
Слайд 3РНК-интерференция у эукариот
Эндрю Файер и Крейг Мелло, открыли явление в 1998 году,
РНК-интерференция у эукариот
Эндрю Файер и Крейг Мелло, открыли явление в 1998 году,

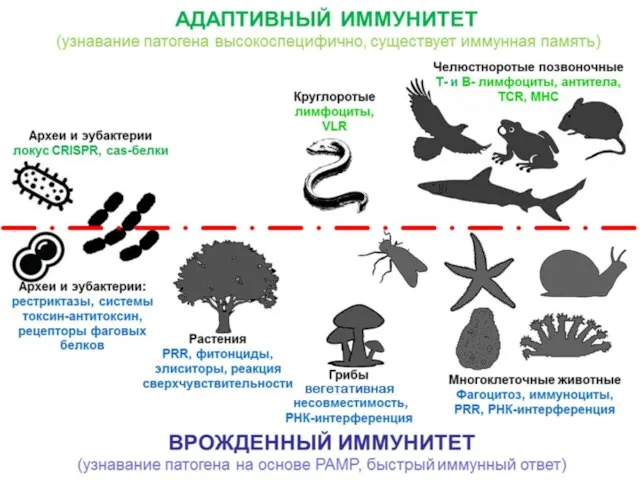
Слайд 4Механизмы подавления экспрессии генов с помощью двуцепочечной РНК (РНК интерференция)
цитоплазма
ядро
Dicer
расщепление и
деградация
Механизмы подавления экспрессии генов с помощью двуцепочечной РНК (РНК интерференция)
цитоплазма
ядро
Dicer
расщепление и
деградация
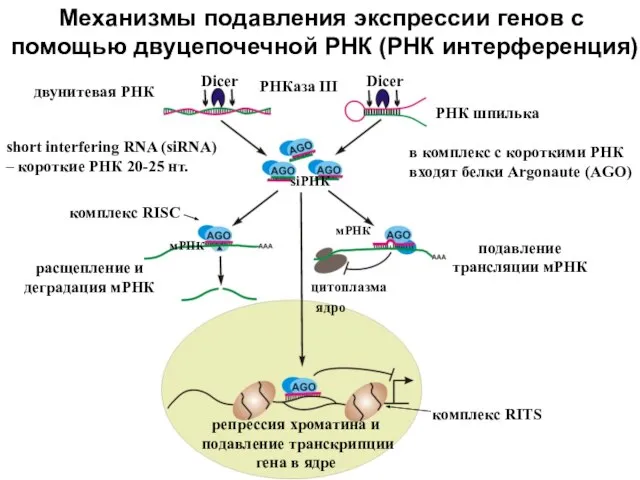
Слайд 5Гидом для pAgo является однонитевая ДНК, мишенью – иногда РНК, иногда ДНК
Pyrococcus
Гидом для pAgo является однонитевая ДНК, мишенью – иногда РНК, иногда ДНК
Pyrococcus
Слайд 6Домены в составе pAgo
pAgo есть у 30% архей и 10% эубактерий,
Домены в составе pAgo
pAgo есть у 30% архей и 10% эубактерий,

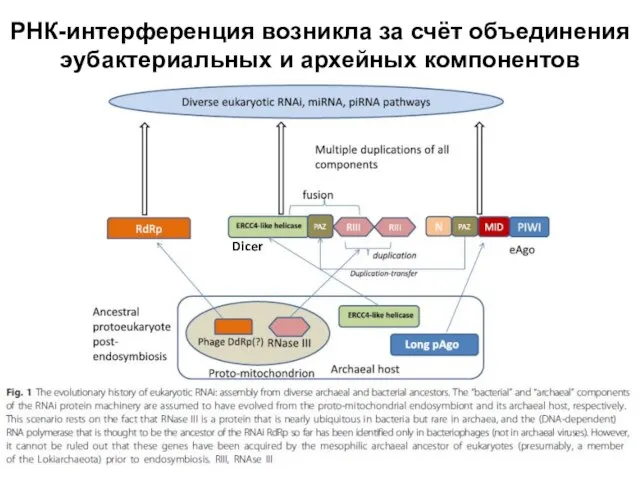
Слайд 7РНК-интерференция возникла за счёт объединения эубактериальных и архейных компонентов
Dicer
РНК-интерференция возникла за счёт объединения эубактериальных и архейных компонентов
Dicer
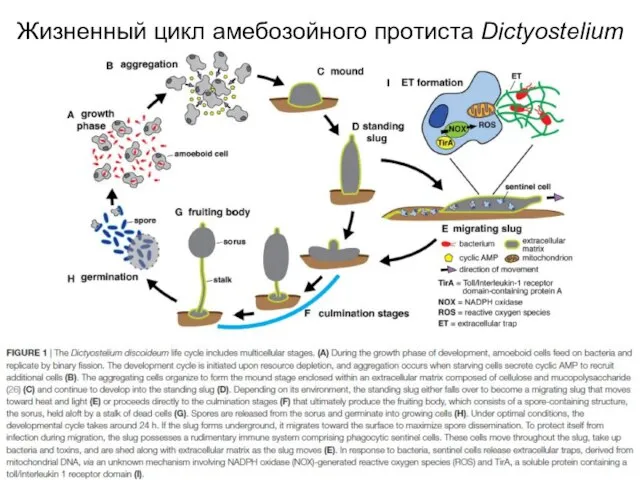
Слайд 8Жизненный цикл амебозойного протиста Dictyostelium
Жизненный цикл амебозойного протиста Dictyostelium
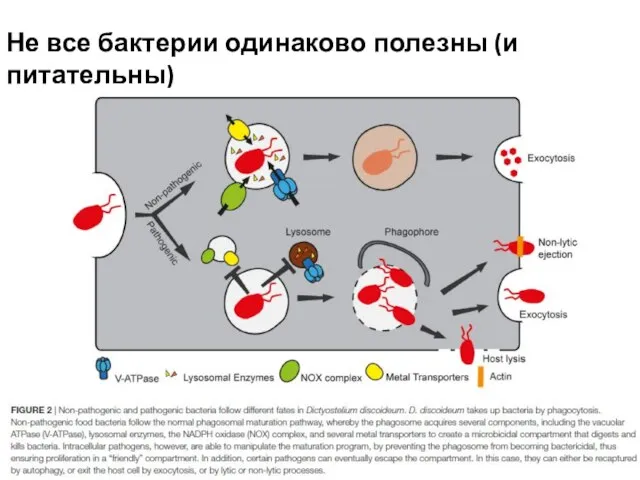
Слайд 9Не все бактерии одинаково полезны (и питательны)
Не все бактерии одинаково полезны (и питательны)
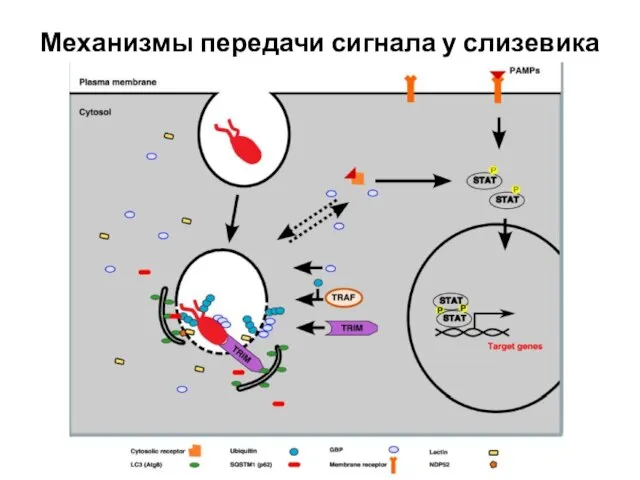
Слайд 10Механизмы передачи сигнала у слизевика
Механизмы передачи сигнала у слизевика
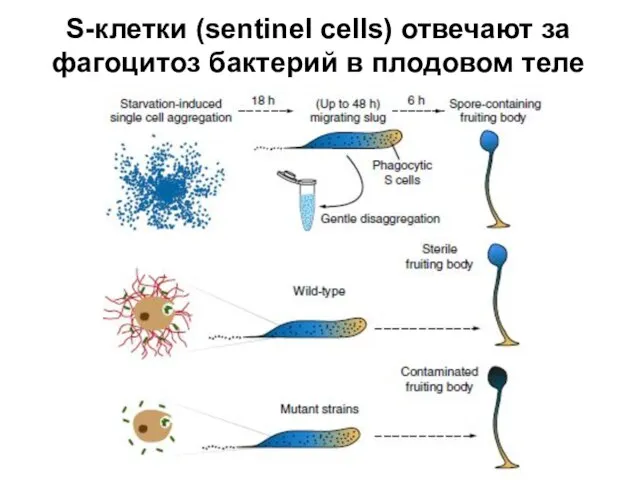
Слайд 11S-клетки (sentinel cells) отвечают за фагоцитоз бактерий в плодовом теле
S-клетки (sentinel cells) отвечают за фагоцитоз бактерий в плодовом теле
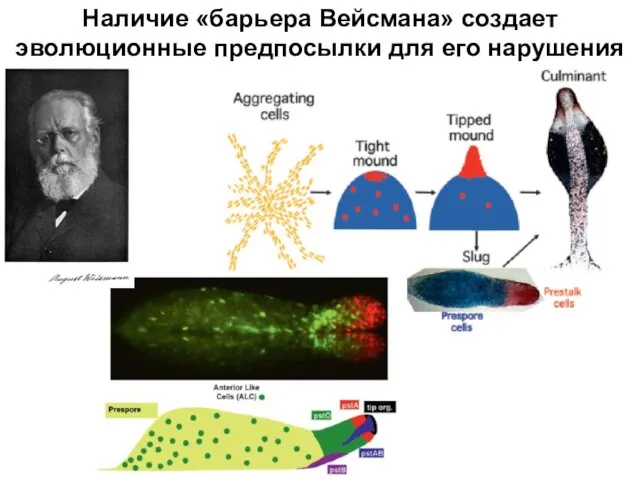
Слайд 12Наличие «барьера Вейсмана» создает эволюционные предпосылки для его нарушения
Наличие «барьера Вейсмана» создает эволюционные предпосылки для его нарушения
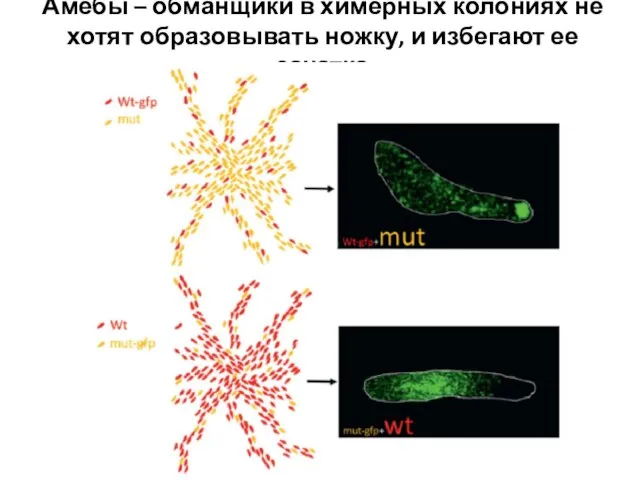
Слайд 13Амебы – обманщики в химерных колониях не хотят образовывать ножку, и избегают
Амебы – обманщики в химерных колониях не хотят образовывать ножку, и избегают
Слайд 14Плодовые тела диктиостелиума сильно структурированы и видоспецифичны, межвидовая агрегация отсутствует
Плодовые тела диктиостелиума сильно структурированы и видоспецифичны, межвидовая агрегация отсутствует

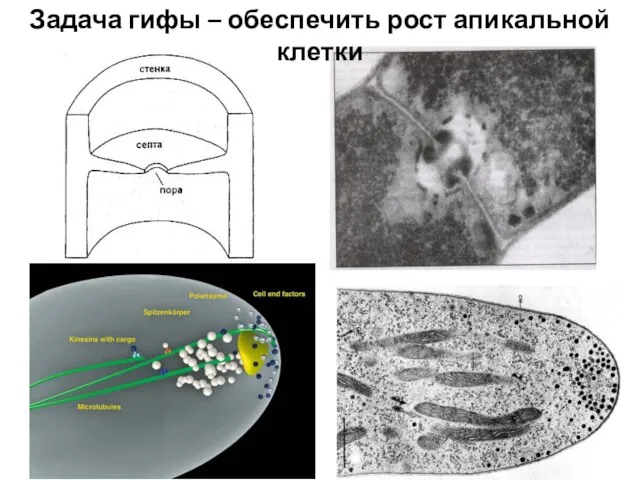
Слайд 15Задача гифы – обеспечить рост апикальной клетки
Задача гифы – обеспечить рост апикальной клетки
Слайд 16Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее делают более прочной,
Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее делают более прочной,

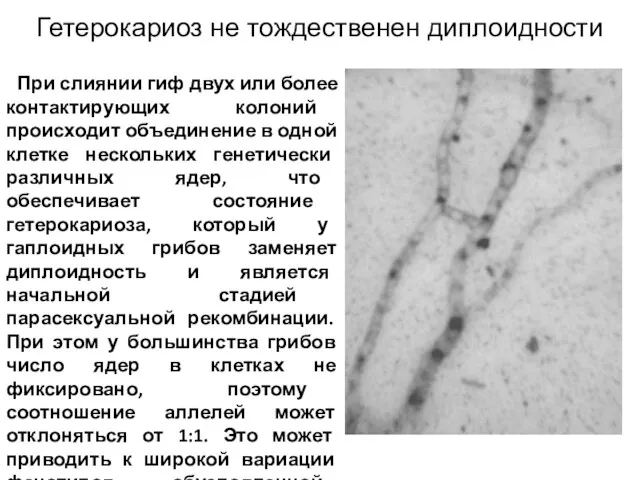
Слайд 17 При слиянии гиф двух или более контактирующих колоний происходит объединение в
При слиянии гиф двух или более контактирующих колоний происходит объединение в
Слайд 18Гетерокариоз – основа парасексуального процесса, т.е. рекомбинации без полового процесса.
Гетерокариоз – основа парасексуального процесса, т.е. рекомбинации без полового процесса.
Слайд 19Иногда мицелии принципиально не срастаются – происходит вегетативная несовместимость
Иногда мицелии принципиально не срастаются – происходит вегетативная несовместимость
Слайд 20Вегетативная несовместимость грибов
Вегетативная несовместимость грибов
Слайд 21Барраж –гибель клеток в зоне контакта двух несовместимых мицелиев
Барраж –гибель клеток в зоне контакта двух несовместимых мицелиев
Слайд 22Гетероаллельность по любому из Het генов вызывает острую реакцию гибели слившихся и
Гетероаллельность по любому из Het генов вызывает острую реакцию гибели слившихся и
Слайд 23Вегетативная несовместимость основана на системе нескольких взаимодействующих генов
Вегетативная несовместимость основана на системе нескольких взаимодействующих генов
Слайд 24Аллели одного и того же гена несовместимости дают комплексный характер сочетаемости
Аллели одного и того же гена несовместимости дают комплексный характер сочетаемости
Слайд 25Гибель клеток в зоне барража связана с избыточной аутофагией
Гибель клеток в зоне барража связана с избыточной аутофагией
Слайд 26Преимущества от вегетативной несовместимости грибов
Защита от эгоистичных паразитов, преодолевающих барьер Вейсмана
Защита от
Преимущества от вегетативной несовместимости грибов
Защита от эгоистичных паразитов, преодолевающих барьер Вейсмана
Защита от

Слайд 27О
Р
Ф
Уровни иммунитета растения
О
Р
Ф
Уровни иммунитета растения

Слайд 28ИММУНИТЕТ НА УРОВНЕ ЦЕЛОГО ОРГАНИЗМА
(первая линия обороны)
Цель – не допустить патогенных организмов
ИММУНИТЕТ НА УРОВНЕ ЦЕЛОГО ОРГАНИЗМА
(первая линия обороны)
Цель – не допустить патогенных организмов

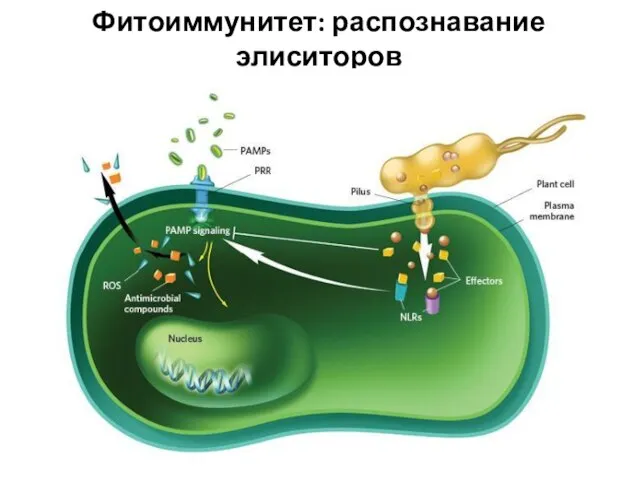
Слайд 29Фитоиммунитет: распознавание элиситоров
Фитоиммунитет: распознавание элиситоров
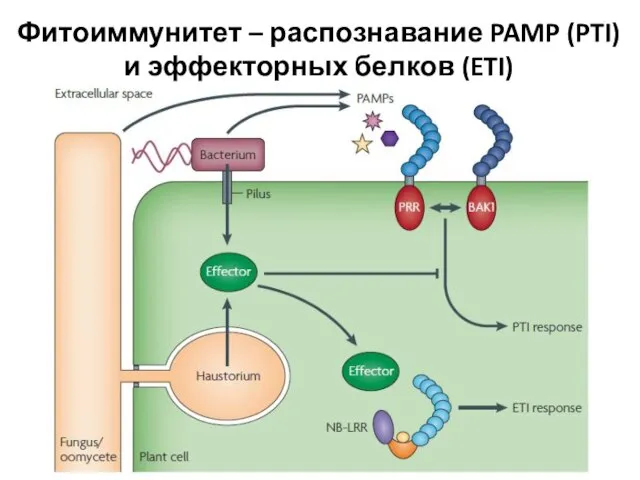
Слайд 30Фитоиммунитет – распознавание PAMP (PTI) и эффекторных белков (ETI)
Фитоиммунитет – распознавание PAMP (PTI) и эффекторных белков (ETI)
Слайд 31PAMP = неспецифические элиситоры
PAMP = неспецифические элиситоры


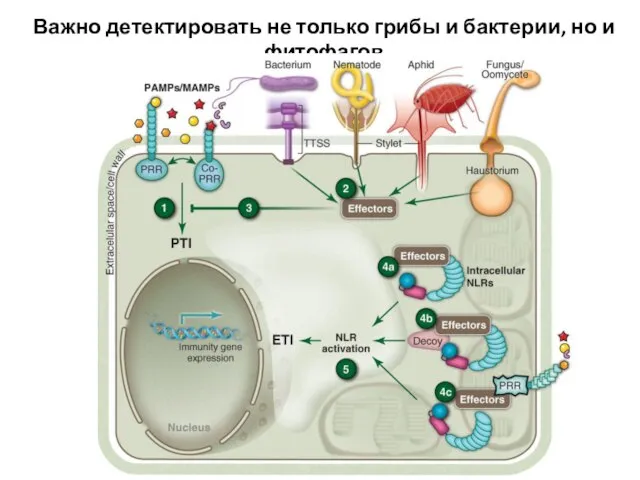
Слайд 33Важно детектировать не только грибы и бактерии, но и фитофагов
Важно детектировать не только грибы и бактерии, но и фитофагов
Слайд 34КАК РАСТЕНИЯ СПРАВЛЯЮТСЯ С ВИРУЛЕНТНЫМИ ПАРАЗИТАМИ (ТРЕТЬЯ ЛИНИЯ ОБОРОНЫ)
У растений возникла система
КАК РАСТЕНИЯ СПРАВЛЯЮТСЯ С ВИРУЛЕНТНЫМИ ПАРАЗИТАМИ (ТРЕТЬЯ ЛИНИЯ ОБОРОНЫ)
У растений возникла система

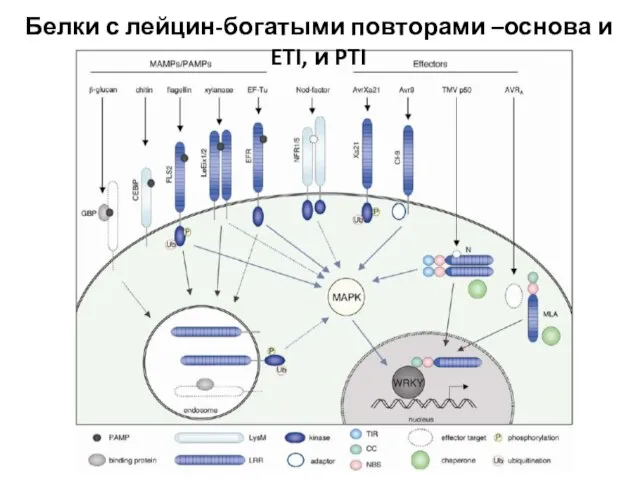
Слайд 35Белки с лейцин-богатыми повторами –основа и ETI, и PTI
Белки с лейцин-богатыми повторами –основа и ETI, и PTI
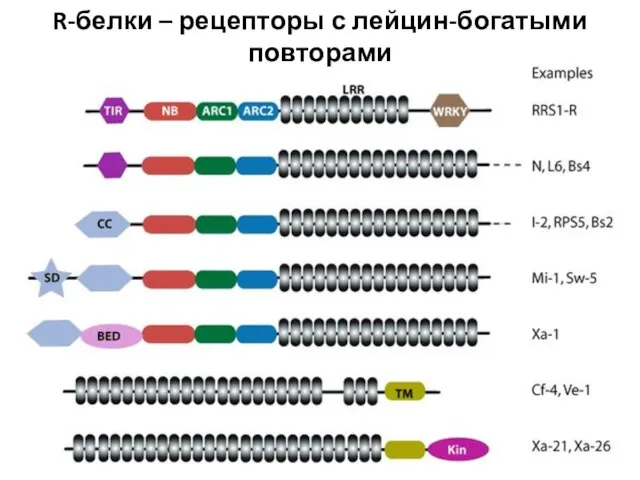
Слайд 36R-белки – рецепторы с лейцин-богатыми повторами
R-белки – рецепторы с лейцин-богатыми повторами
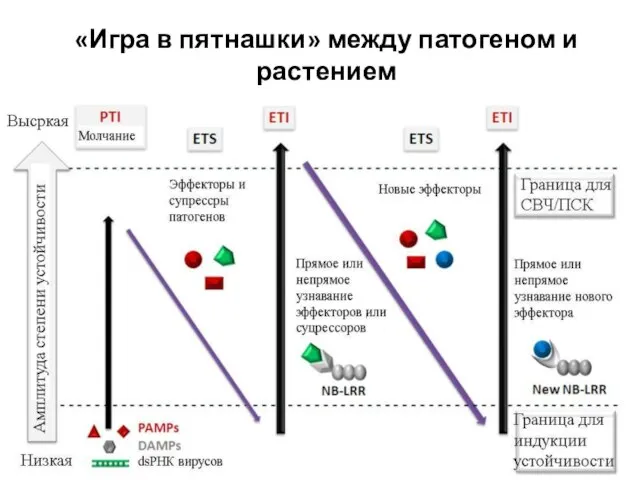
Слайд 37«Игра в пятнашки» между патогеном и растением
«Игра в пятнашки» между патогеном и растением
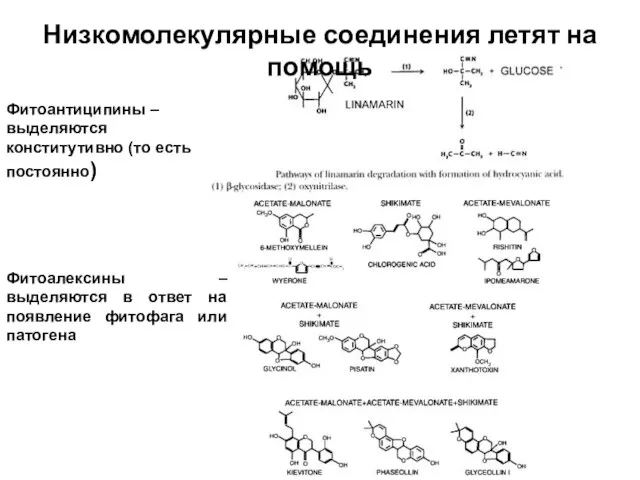
Слайд 38Фитоантиципины – выделяются конститутивно (то есть постоянно)
Фитоалексины –выделяются в ответ на появление
Фитоантиципины – выделяются конститутивно (то есть постоянно)
Фитоалексины –выделяются в ответ на появление
Слайд 39Эффекторные белки фитоиммунитета
Дефензины – низкомолекулярные (45-54 аминокислот) белки, богатые цистеином. Продуцируются млекопитающими,
Эффекторные белки фитоиммунитета
Дефензины – низкомолекулярные (45-54 аминокислот) белки, богатые цистеином. Продуцируются млекопитающими,

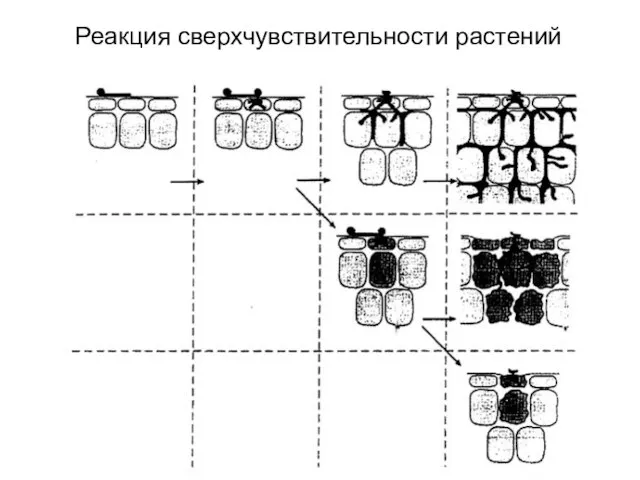
Слайд 40Реакция сверхчувствительности растений
Реакция сверхчувствительности растений
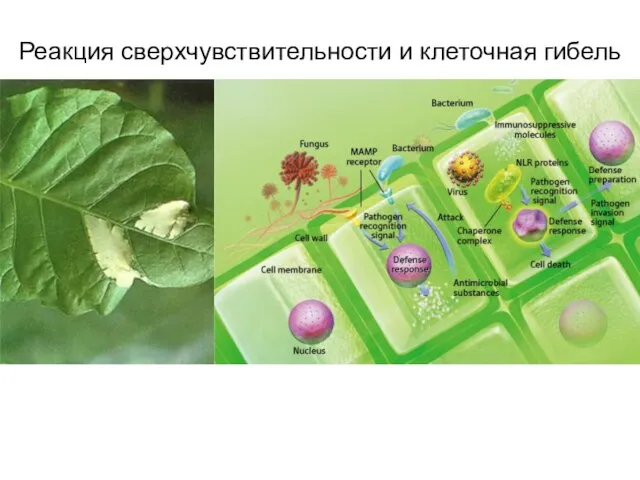
Слайд 41Реакция сверхчувствительности и клеточная гибель
Реакция сверхчувствительности и клеточная гибель
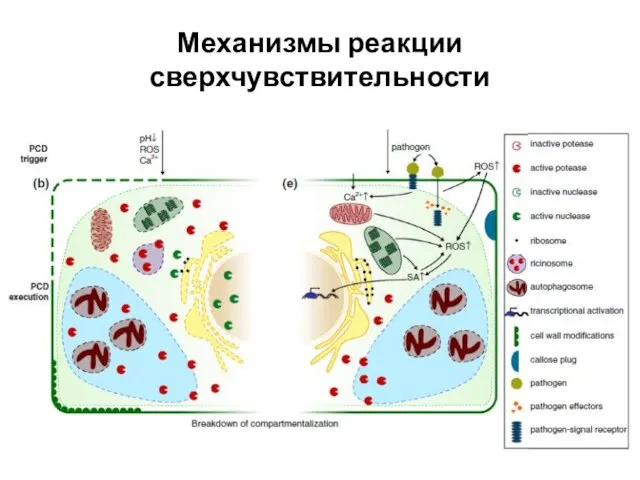
Слайд 42Механизмы реакции сверхчувствительности
Механизмы реакции сверхчувствительности
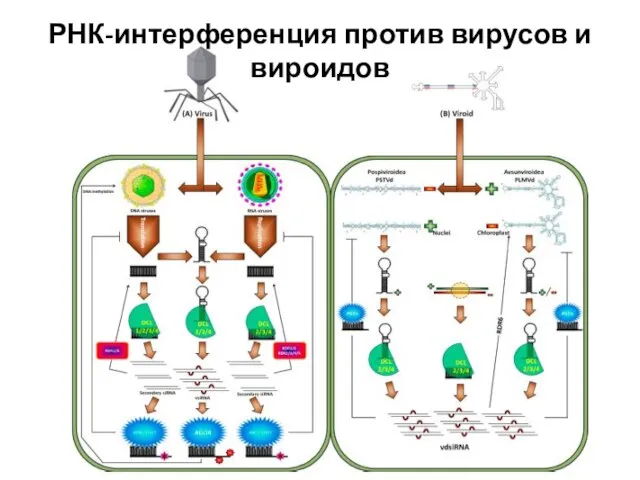
Слайд 43РНК-интерференция против вирусов и вироидов
РНК-интерференция против вирусов и вироидов
Слайд 44микроРНК как переключатель типов иммунного ответа у растений
микроРНК как переключатель типов иммунного ответа у растений

 Титанобоа - вид вымерших змей
Титанобоа - вид вымерших змей Семейство Колокольчиковые (Campanulaceae)
Семейство Колокольчиковые (Campanulaceae) Строение голосеменных
Строение голосеменных Обмен веществ и энергии в клетках
Обмен веществ и энергии в клетках Перелетные птицы
Перелетные птицы Орган зрения
Орган зрения Работы И.В.Мичурина
Работы И.В.Мичурина А.Блок Ворона
А.Блок Ворона Фотоальбом. Ботаника
Фотоальбом. Ботаника Городской цветник Аленький цветочек
Городской цветник Аленький цветочек Патология ультраструктур клетки
Патология ультраструктур клетки Дары природы. Иван-чай – наш домашний лекарь
Дары природы. Иван-чай – наш домашний лекарь Микробиология
Микробиология Чарльз Дарвин об эволюции органического мира
Чарльз Дарвин об эволюции органического мира Система восприятия человеком состояния внешней среды
Система восприятия человеком состояния внешней среды Такса: описание и характеристика породы
Такса: описание и характеристика породы Тренировочный тест. Птицы
Тренировочный тест. Птицы Презентация на тему Птицы леса
Презентация на тему Птицы леса  Классификация мутаций. Генные, хромосомные и геномные мутации
Классификация мутаций. Генные, хромосомные и геномные мутации Презентация на тему Великие естествоиспытатели
Презентация на тему Великие естествоиспытатели  Животные. Разнообразие
Животные. Разнообразие Генетическая информация
Генетическая информация Женская половая система
Женская половая система Презентация на тему Особенности высшей нервной деятельности человека
Презентация на тему Особенности высшей нервной деятельности человека  Размножение и развитие организмов
Размножение и развитие организмов Гидропоника
Гидропоника Воздух в жизни растений 6 класс
Воздух в жизни растений 6 класс Сосна Корейская (кедр). Сирень. Жимолость Тельмана. Кирказон крупнолистный (аристолохия)
Сосна Корейская (кедр). Сирень. Жимолость Тельмана. Кирказон крупнолистный (аристолохия)