- Катаболизм аминокислот

Содержание
- 2. Аминокислоты – предшественники для синтеза, в первую оче- редь, белка, а также для синтеза других биоактивных
- 3. Пути превращения аминокислот в организме
- 4. Участие аминокислот в обмене Помимо участия в синтезе пептидов и белков, большинство аминокислот участвуют во многих
- 5. Метаболическая судьба углеродных «скелетов» аминокислот
- 6. Катаболизм аминокислот. Аминокислота теряет NH2-группу двумя путями: 1. Дезаминирование 2. Трансаминирование
- 7. Дезаминирование аминокислот Дезаминирование – реакция отщепления α-NH2-группы от аминокислоты. АМИНОКИСЛОТА α-КЕТОКИСЛОТА + NH4 (безазотистое соединение) (аммиак)
- 8. ЧЕТЫРЕ ТИПА ДЕЗАМИНИРОВАНИЯ: У человека – основной тип дезаминирования: ОКИСЛИТЕЛЬНЫЙ. Исключения: неокислительное дезаминирование (печень) а). Сер
- 9. ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ Реакция обратима: при повышении [NH4] идёт в обратном направле- нии – восстановительное аминирование α-КГ.
- 10. Кофермент глутамат-ДГ - НАД+ или НАДФ+ Никотинамид аденин динуклеотид (фосфат) Предшественник – водорастворимый витамин РР (никотиновая
- 11. НЕПРЯМОЕ ДЕЗАМИНИОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ) Большинство аминокислот дезаминируется в две стадии: I стадия: Трансаминирование аминотрансфераза (кофермент ПФ) аминокислота
- 12. ОКСИДАЗЫ L- и D-АМИНОКИСЛОТ Оксидазы присутствуют в печени и почках – дезаминируют неко- торые аминокислоты: Оксидаза
- 13. ФМН – флавинмононуклеотид и ФАД - флавинадениндинуклеотид ФМН – простетическая группа НАДН-ДГ дыхательной цепи митохондрий: 2e,
- 14. Восстановление ФМН и ФАД происходит по изоаллоксазиновому кольцу
- 15. ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ Реакция переноса –NH2 от α-аминокислоты на α-кетокислоту. Итог: образование новой кетокислоты и новой аминокислоты.
- 16. МЕХАНИЗМ РЕАКЦИЙ ТРАНСАМИНИРОВАНЯ У всех аминотрансфераз простетической группой в активном центре яляется пиридоксальфосфат – производное пиридоксина
- 17. Пиридоксальфосфат образует с субстратом промежуточный продукт – шиффово основание (содержит альдиминовую связь): Альдегидная гр. ПФ связана
- 18. В ходе катализа в шиффовом основании изменяется положе- ние двойной связи. Образуется кетимин, который далее превра-
- 19. В результате работы аминотрансфераз азот аминогрупп многих А.К. переходит в состав аминогруппы Глу. Эти реакции проходят
- 20. ДЕКАРБОКСИЛИРОВАНИЕ АМИНОКИСЛОТ Реакция отщепления карбоксильной группы с образованем ами- нов: пиридоксальфосфат Реакции декарбоксилирования А.К., в отличие
- 22. Скачать презентацию

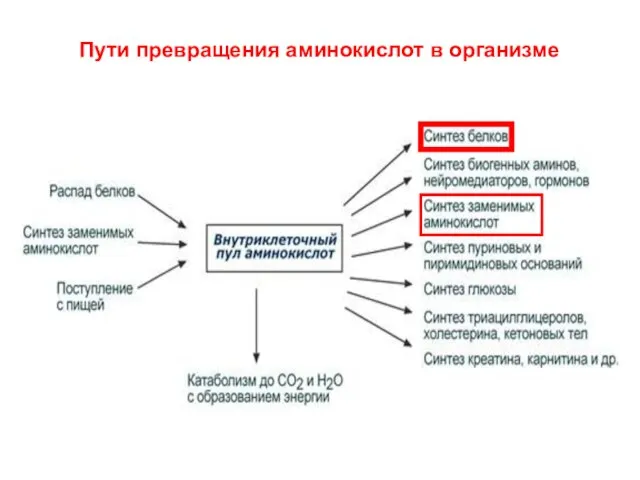
Слайд 3Пути превращения аминокислот в организме
Пути превращения аминокислот в организме
Слайд 4Участие аминокислот в обмене
Помимо участия в синтезе пептидов и белков, большинство
Участие аминокислот в обмене
Помимо участия в синтезе пептидов и белков, большинство

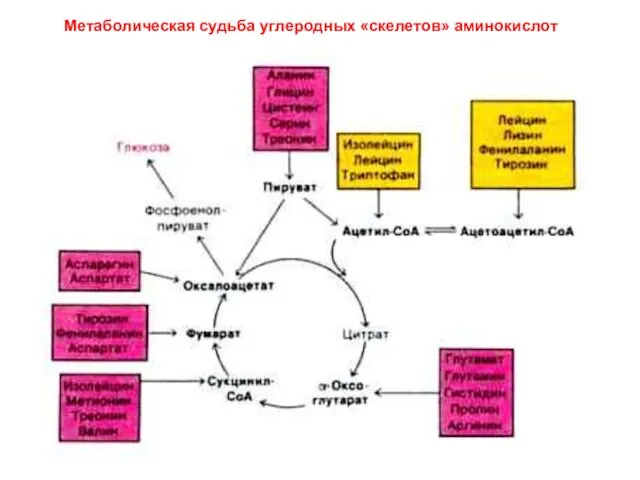
Слайд 5Метаболическая судьба углеродных «скелетов» аминокислот
Метаболическая судьба углеродных «скелетов» аминокислот
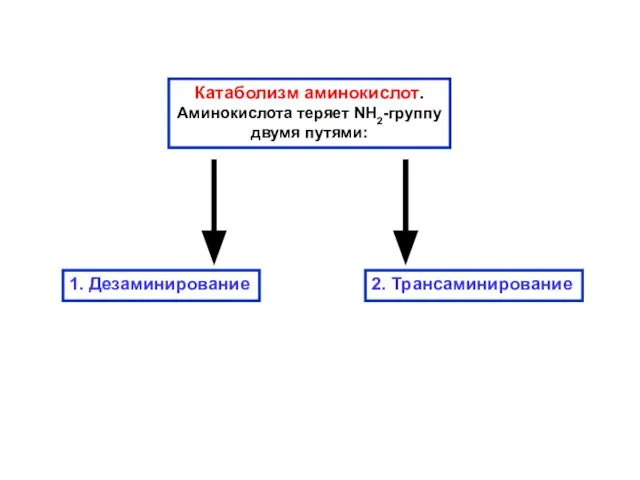
Слайд 6Катаболизм аминокислот.
Аминокислота теряет NH2-группу
двумя путями:
1. Дезаминирование
2. Трансаминирование
Катаболизм аминокислот.
Аминокислота теряет NH2-группу
двумя путями:
1. Дезаминирование
2. Трансаминирование
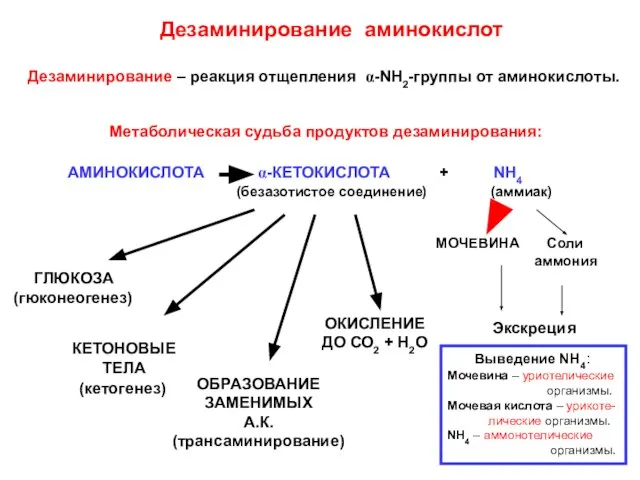
Слайд 7Дезаминирование аминокислот
Дезаминирование – реакция отщепления α-NH2-группы от аминокислоты.
АМИНОКИСЛОТА α-КЕТОКИСЛОТА + NH4
Дезаминирование аминокислот
Дезаминирование – реакция отщепления α-NH2-группы от аминокислоты.
АМИНОКИСЛОТА α-КЕТОКИСЛОТА + NH4
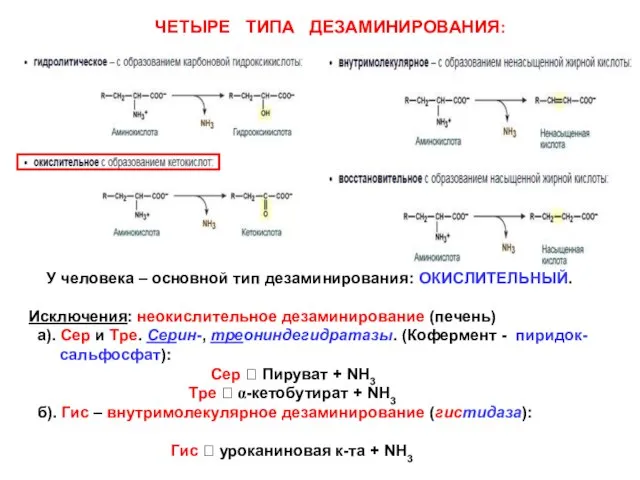
Слайд 8ЧЕТЫРЕ ТИПА ДЕЗАМИНИРОВАНИЯ:
У человека – основной тип дезаминирования: ОКИСЛИТЕЛЬНЫЙ.
Исключения: неокислительное дезаминирование
ЧЕТЫРЕ ТИПА ДЕЗАМИНИРОВАНИЯ:
У человека – основной тип дезаминирования: ОКИСЛИТЕЛЬНЫЙ.
Исключения: неокислительное дезаминирование
Слайд 9ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
Реакция обратима: при повышении [NH4] идёт в обратном направле-
нии –
ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ
Реакция обратима: при повышении [NH4] идёт в обратном направле-
нии –
![ОКИСЛИТЕЛЬНОЕ ДЕЗАМИНИРОВАНИЕ Реакция обратима: при повышении [NH4] идёт в обратном направле- нии](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/905409/slide-8.jpg)
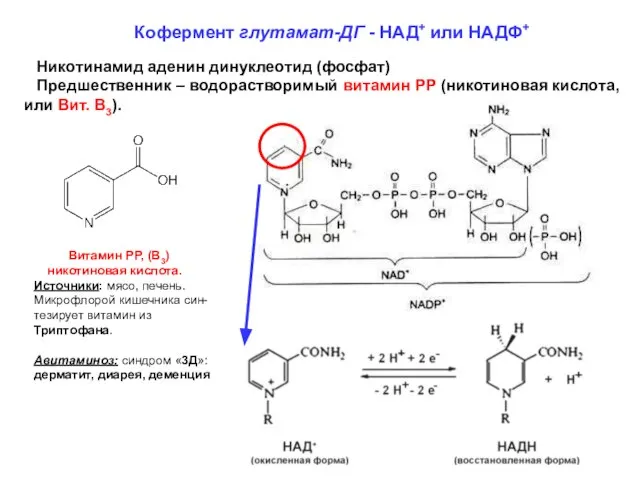
Слайд 10Кофермент глутамат-ДГ - НАД+ или НАДФ+
Никотинамид аденин динуклеотид (фосфат)
Предшественник –
Кофермент глутамат-ДГ - НАД+ или НАДФ+
Никотинамид аденин динуклеотид (фосфат)
Предшественник –
Слайд 11НЕПРЯМОЕ ДЕЗАМИНИОВАНИЕ
(ТРАНСДЕЗАМИНИРОВАНИЕ)
Большинство аминокислот дезаминируется в две стадии:
I стадия: Трансаминирование
аминотрансфераза
НЕПРЯМОЕ ДЕЗАМИНИОВАНИЕ
(ТРАНСДЕЗАМИНИРОВАНИЕ)
Большинство аминокислот дезаминируется в две стадии:
I стадия: Трансаминирование
аминотрансфераза

Слайд 12ОКСИДАЗЫ L- и D-АМИНОКИСЛОТ
Оксидазы присутствуют в печени и почках – дезаминируют
ОКСИДАЗЫ L- и D-АМИНОКИСЛОТ
Оксидазы присутствуют в печени и почках – дезаминируют

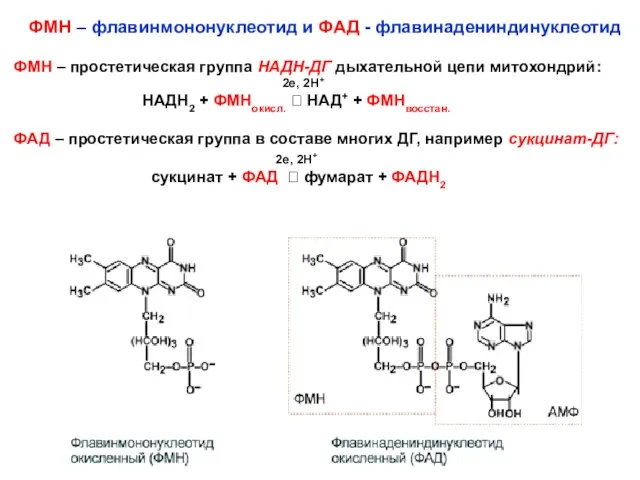
Слайд 13ФМН – флавинмононуклеотид и ФАД - флавинадениндинуклеотид
ФМН – простетическая группа НАДН-ДГ дыхательной
ФМН – флавинмононуклеотид и ФАД - флавинадениндинуклеотид
ФМН – простетическая группа НАДН-ДГ дыхательной
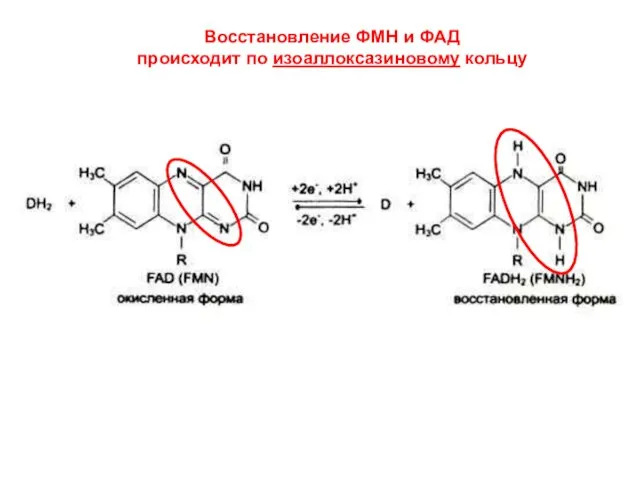
Слайд 14Восстановление ФМН и ФАД
происходит по изоаллоксазиновому кольцу
Восстановление ФМН и ФАД
происходит по изоаллоксазиновому кольцу
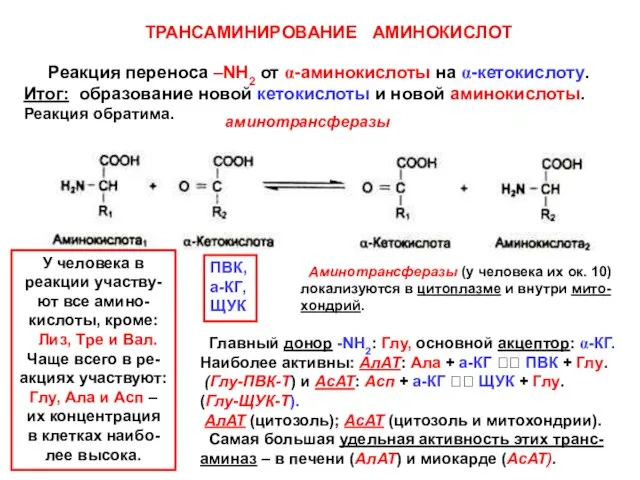
Слайд 15ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ
Реакция переноса –NH2 от α-аминокислоты на α-кетокислоту.
Итог: образование новой
ТРАНСАМИНИРОВАНИЕ АМИНОКИСЛОТ
Реакция переноса –NH2 от α-аминокислоты на α-кетокислоту.
Итог: образование новой
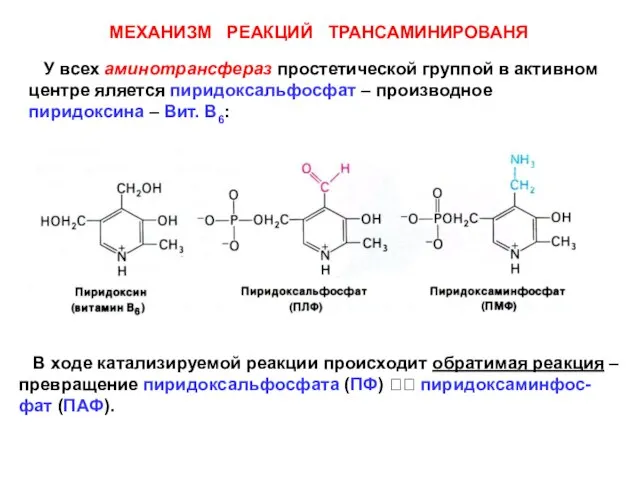
Слайд 16МЕХАНИЗМ РЕАКЦИЙ ТРАНСАМИНИРОВАНЯ
У всех аминотрансфераз простетической группой в активном центре яляется
МЕХАНИЗМ РЕАКЦИЙ ТРАНСАМИНИРОВАНЯ
У всех аминотрансфераз простетической группой в активном центре яляется
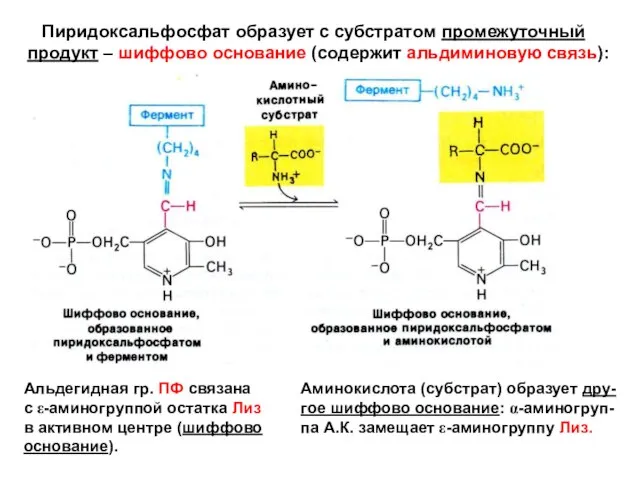
Слайд 17 Пиридоксальфосфат образует с субстратом промежуточный
продукт – шиффово основание (содержит альдиминовую
Пиридоксальфосфат образует с субстратом промежуточный
продукт – шиффово основание (содержит альдиминовую
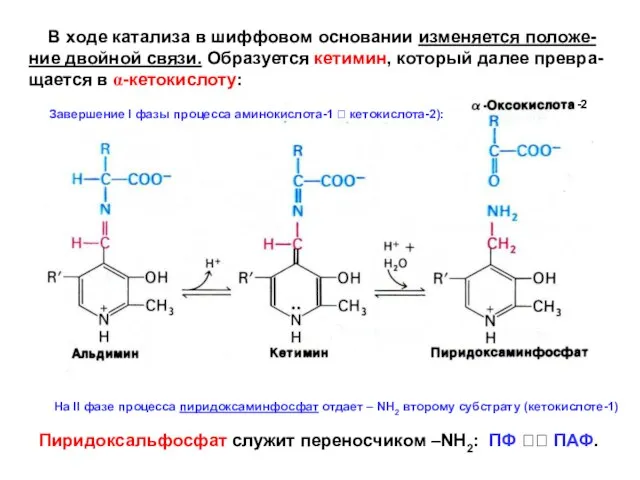
Слайд 18 В ходе катализа в шиффовом основании изменяется положе-
ние двойной связи. Образуется
В ходе катализа в шиффовом основании изменяется положе-
ние двойной связи. Образуется
Слайд 19 В результате работы аминотрансфераз азот аминогрупп многих
А.К. переходит в состав
В результате работы аминотрансфераз азот аминогрупп многих
А.К. переходит в состав

Слайд 20ДЕКАРБОКСИЛИРОВАНИЕ АМИНОКИСЛОТ
Реакция отщепления карбоксильной группы с образованем ами-
нов:
пиридоксальфосфат
Реакции декарбоксилирования А.К.,
ДЕКАРБОКСИЛИРОВАНИЕ АМИНОКИСЛОТ
Реакция отщепления карбоксильной группы с образованем ами-
нов:
пиридоксальфосфат
Реакции декарбоксилирования А.К.,

 Брюхоногие моллюски
Брюхоногие моллюски Построение вариационной кривой
Построение вариационной кривой Презентация на тему Особенности скелета человека
Презентация на тему Особенности скелета человека  PPT Energy-1
PPT Energy-1 Хвойные растения
Хвойные растения According to the soil specialization. (And some botanical science)
According to the soil specialization. (And some botanical science) Интересная зоология: разработка комплекса приёмов работы с детьми по обобщению школьного курса зоологии в 7 классе
Интересная зоология: разработка комплекса приёмов работы с детьми по обобщению школьного курса зоологии в 7 классе Подземный житель — крот
Подземный житель — крот Приспособленность растений к жизни в дубраве
Приспособленность растений к жизни в дубраве Презентация на тему Мой родной край виртуальное путешествие
Презентация на тему Мой родной край виртуальное путешествие  Как растет чай
Как растет чай Класс Пресмыкающиеся. Отряд Чешуйчатые
Класс Пресмыкающиеся. Отряд Чешуйчатые Презентация на тему ПРЕДМЕТ И ЗАДАЧИ БИОТЕХНОЛОГИИ
Презентация на тему ПРЕДМЕТ И ЗАДАЧИ БИОТЕХНОЛОГИИ  Құрғақ және қуаң дала фаунасы
Құрғақ және қуаң дала фаунасы Системы объектов
Системы объектов Презентация на тему ИСПАРЕНИЕ ВОДЫ РАСТЕНИЯМИ
Презентация на тему ИСПАРЕНИЕ ВОДЫ РАСТЕНИЯМИ  Свет и зрение
Свет и зрение Естественные защитные силы организма
Естественные защитные силы организма Опыление и оплодотворение у растений
Опыление и оплодотворение у растений Плоские черви. Урок 3
Плоские черви. Урок 3 Вегетативная нервная система
Вегетативная нервная система Пчеловодство
Пчеловодство Законы генетики. Врожденная близорукость
Законы генетики. Врожденная близорукость Законы наследственности. Типы гибридизации
Законы наследственности. Типы гибридизации Покрытосеменные растения
Покрытосеменные растения Сходство зародышей и эмбриональная дивергенция. Биогенетический закон. Развитие организмов и окружающая среда
Сходство зародышей и эмбриональная дивергенция. Биогенетический закон. Развитие организмов и окружающая среда Возр. особенности скелета (2)
Возр. особенности скелета (2) Лекарственные растения и их роль в жизни человека
Лекарственные растения и их роль в жизни человека