- Key points to discuss about Genetic load

Содержание
- 2. Key Concepts: Genetic load is the reduction in mean fitness of a population caused by some
- 3. What is genetic load ?
- 4. Genetic load is the difference between the fitness of an average genotype in a population and
- 5. Genetic Load Sources The mutation load is the decrease in fitness or viability (or other trait
- 6. Direct Evidence of change in Mutational load in Humans. The extent to which selection has shaped
- 7. Genetic Load the Haldanes Dilemma.
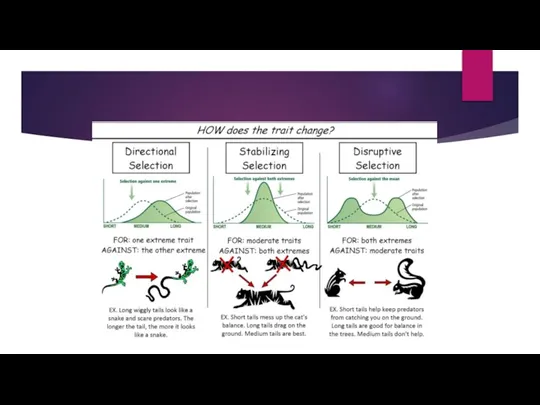
- 8. Directional Selection. In population genetics, directional selection, or positive selection is a mode of natural selection
- 9. Some Key points About Directional Selection. Directional selection occurs when individuals homozygous for one allele have
- 10. When a dominant favored allele is rare most individuals carrying it are heterozygous, and the large
- 11. Effects of Sexual Selection on the Heritability of trait. Strong directional selection usually exhausts additive genetic
- 12. There are a number of possible explanations for why selection does not eliminate all of the
- 14. Plasmids role as an genetic Load Plasmids represent an added genetic load to their host cell
- 15. Substitutional load substitutional load In genetics, the cost in genetic deaths to the population of replacing
- 16. Segregational or recombination load high levels of heterozygosity could be maintained by overdominant selection at multiple
- 17. Causes : Deleterious mutation Deleterious mutation load is the main contributing factor to genetic load overall.[5]
- 18. New beneficial mutations create fitter genotypes than those previously present in the population. When load is
- 19. Inbreeding Inbreeding increases homozygosity. In the short run, an increase in inbreeding increases the probability with
- 21. Скачать презентацию

Слайд 3What is genetic load ?
What is genetic load ?

Слайд 4Genetic load is the difference between the fitness of an average genotype in a population and the fitness of
Genetic load is the difference between the fitness of an average genotype in a population and the fitness of

Слайд 5Genetic Load Sources
The mutation load is the decrease in fitness or viability
Genetic Load Sources
The mutation load is the decrease in fitness or viability

Слайд 6Direct Evidence of change in Mutational load in Humans.
The extent to which
Direct Evidence of change in Mutational load in Humans.
The extent to which

Слайд 7Genetic Load the Haldanes Dilemma.
Genetic Load the Haldanes Dilemma.

Слайд 8Directional Selection.
In population genetics, directional selection, or positive selection is a mode of natural selection in which
Directional Selection.
In population genetics, directional selection, or positive selection is a mode of natural selection in which

Слайд 9Some Key points About Directional Selection.
Directional selection occurs when individuals homozygous for one
Some Key points About Directional Selection.
Directional selection occurs when individuals homozygous for one

Слайд 10When a dominant favored allele is rare most individuals carrying it are
When a dominant favored allele is rare most individuals carrying it are

Слайд 11Effects of Sexual Selection on the Heritability of trait.
Strong directional selection usually exhausts additive
Effects of Sexual Selection on the Heritability of trait.
Strong directional selection usually exhausts additive

Слайд 12There are a number of possible explanations for why selection does not
There are a number of possible explanations for why selection does not

Слайд 14Plasmids role as an genetic Load
Plasmids represent an added genetic load
Plasmids role as an genetic Load
Plasmids represent an added genetic load

Слайд 15Substitutional load
substitutional load In genetics, the cost in genetic deaths to the population
Substitutional load
substitutional load In genetics, the cost in genetic deaths to the population

Слайд 16Segregational or recombination load
high levels of heterozygosity could be maintained by overdominant selection at
Segregational or recombination load
high levels of heterozygosity could be maintained by overdominant selection at

Слайд 17Causes :
Deleterious mutation
Deleterious mutation load is the main contributing factor to genetic load
Causes :
Deleterious mutation
Deleterious mutation load is the main contributing factor to genetic load

Слайд 18
New beneficial mutations create fitter genotypes than those previously present in
New beneficial mutations create fitter genotypes than those previously present in

Слайд 19Inbreeding
Inbreeding increases homozygosity. In the short run, an increase in inbreeding increases the
Inbreeding
Inbreeding increases homozygosity. In the short run, an increase in inbreeding increases the

 Организм как единое целое
Организм как единое целое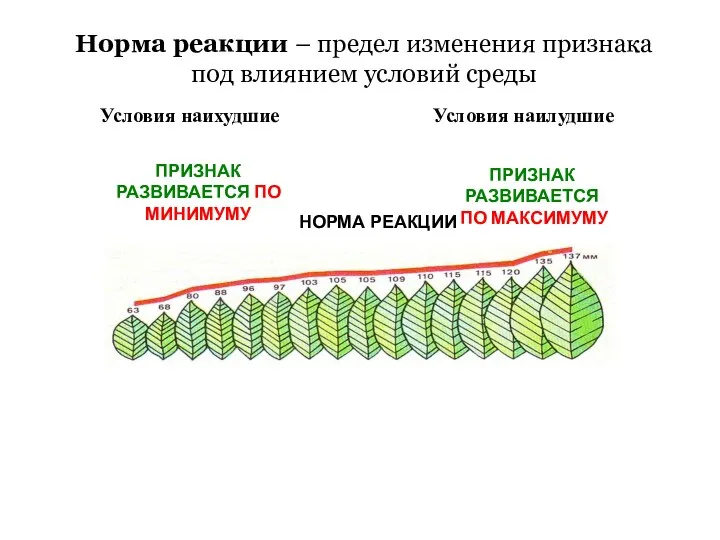 Норма реакции – предел изменения признака под влиянием условий среды. Часть 4-2
Норма реакции – предел изменения признака под влиянием условий среды. Часть 4-2 Rośliny dwuletnie na balkon i taras
Rośliny dwuletnie na balkon i taras Наследственность и среда в межиндивидуальной вариативности ЭЭГ. Метод исследования головного мозга
Наследственность и среда в межиндивидуальной вариативности ЭЭГ. Метод исследования головного мозга Мозг - центральный отдел нервной системы
Мозг - центральный отдел нервной системы Эволюционные учения
Эволюционные учения Wolverine К
Wolverine К genetika
genetika Презентация на тему Строение клетки (10 класс)
Презентация на тему Строение клетки (10 класс)  Специализация кошки
Специализация кошки Дыхательная система человека
Дыхательная система человека Сможет ли двугорбый верблюд жить в Сахаре?
Сможет ли двугорбый верблюд жить в Сахаре? Гипотезы происхождения жизни
Гипотезы происхождения жизни Генетика человека
Генетика человека Популяция и ее генетическая структура
Популяция и ее генетическая структура Биополимеры
Биополимеры Животные жарких стран и Севера
Животные жарких стран и Севера Введение в физиологию. Физиология возбудимых тканей. Лекция 1
Введение в физиологию. Физиология возбудимых тканей. Лекция 1 Отчет по практике. Ярус травянистых растений пришкольного участка Муниципального бюджетного общеобразовательного учреждения
Отчет по практике. Ярус травянистых растений пришкольного участка Муниципального бюджетного общеобразовательного учреждения Изучение состава почвенных водорослей
Изучение состава почвенных водорослей Доберман. Происхождение породы
Доберман. Происхождение породы Презентация на тему Скелет и мышцы птицы
Презентация на тему Скелет и мышцы птицы  Строение и деятельность внутренних органов земноводных
Строение и деятельность внутренних органов земноводных Как мы выращивали грибы вешенки. 2 класс
Как мы выращивали грибы вешенки. 2 класс Викторина про собак
Викторина про собак Анализаторы. Строение и функции зрительного анализатора
Анализаторы. Строение и функции зрительного анализатора Космостық биотехнология және биоқауіпсіздік талаптары
Космостық биотехнология және биоқауіпсіздік талаптары Основные группы микроорганизмов
Основные группы микроорганизмов