- Кинетика. Ингибирование ферментов. Обмен веществ. Тема 3

Содержание
- 2. Клеточная организация ферментативной активности Ферменты располагаются в субклеточных структурах (органеллах) соответственно их функциям. Например: а) в
- 3. Скорость ферментативных реакций, как и всяких других реакций, зависит от температуры: при повышении температуры на каждые
- 4. При высоких температурах, более 55–60 оС активность фермента резко снижается из-за его тепловой денатурации, и, как
- 5. При высоких температурах, более 55–60 оС активность фермента резко снижается из-за его тепловой денатурации, и, как
- 6. Скорость ферментативной реакции имеет свой температурный оптимум, превышение которого приводит к понижению активности ферментов из-за тепловой
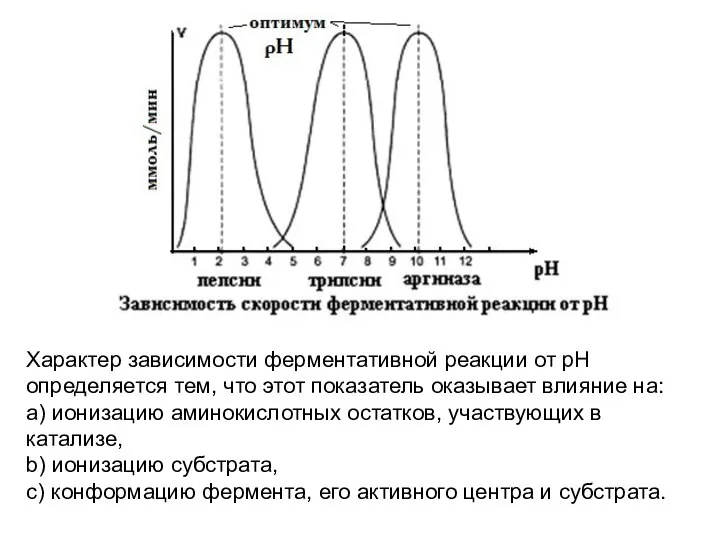
- 7. Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает влияние на: a) ионизацию
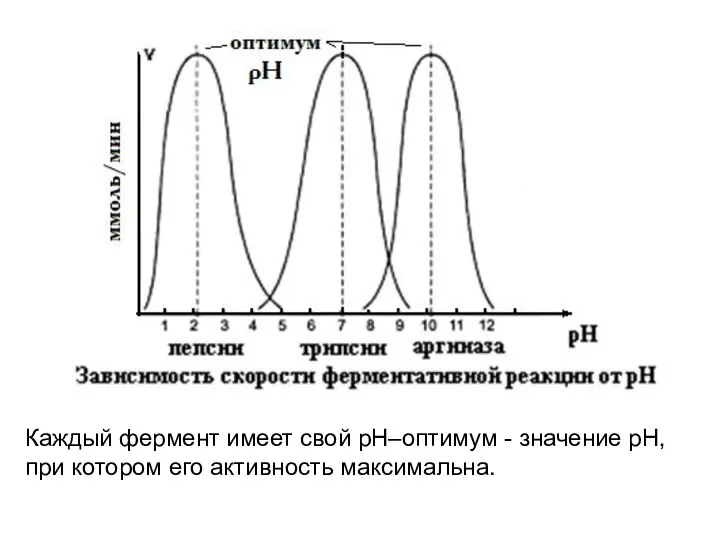
- 8. Каждый фермент имеет свой рН–оптимум - значение рН, при котором его активность максимальна.
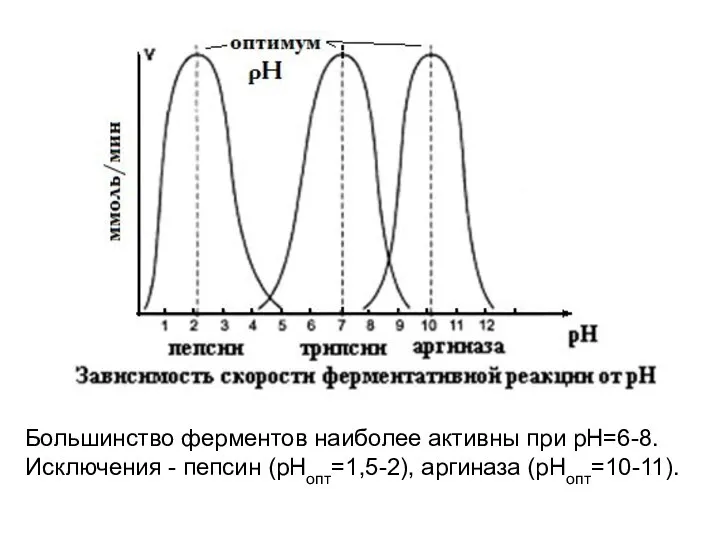
- 9. Большинство ферментов наиболее активны при рН=6-8. Исключения - пепсин (рНопт=1,5-2), аргиназа (рНопт=10-11).
- 10. Простейшая кинетическая схема взаимодействия фермента (Е) и субстрата (S). Во время реакции молекула фермента, E, и
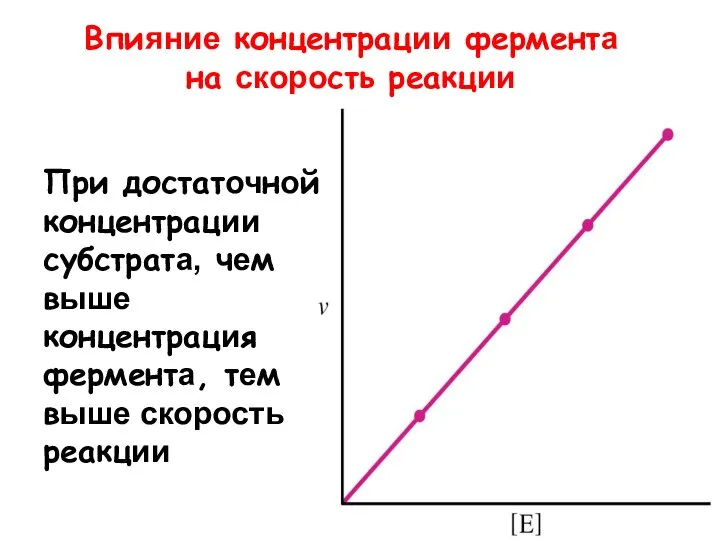
- 11. Впияние концентрации фермента на скорость реакции При достаточной концентрации субстрата, чем выше концентрация фермента, тем выше
- 12. Влияние концентрации субстрата на скорость реакции
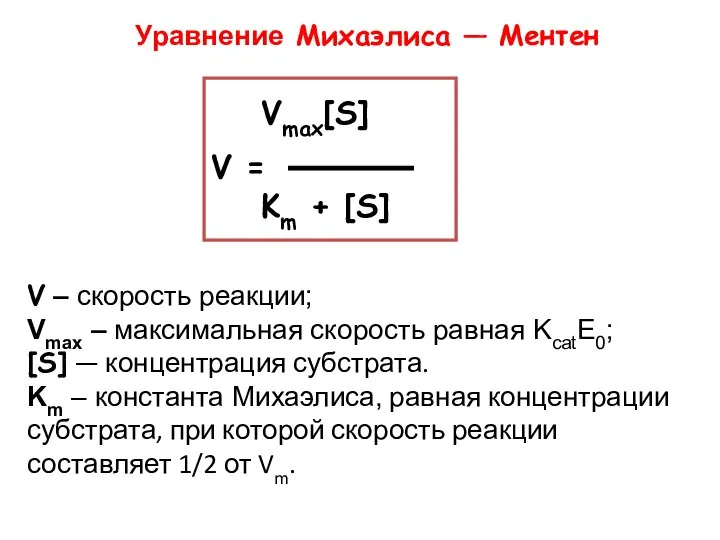
- 13. V – скорость реакции; Vmax – максимальная скорость равная KcatE0; [S] — концентрация субстрата. Km –
- 14. При фиксированной концентрации фермента начальная скорость реакции линейно пропорциональна концентрации субстрата, если последняя маленькая, но не
- 15. Уравнение Михаэлиса — Ментен V – скорость реакции; Vmax – максимальная скорость равная KcatE0; [S] —
- 16. Скорость катализа Уравнение Михаэлиса—Ментен — основное уравнение ферментативной кинетики, описывает зависимость скорости реакции, катализируемой ферментом, от
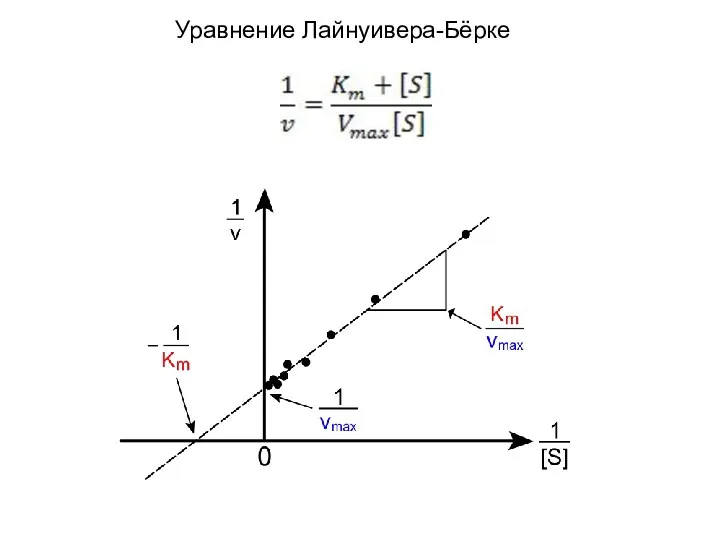
- 17. Уравнение Лайнуивера-Бёрке
- 18. Когда [S] много больше чем Km скорость реакции не зависит от [S] что соответствует нулевому порядку
- 19. Ингибирование ферментов Разные химические агенты (метаболиты, аналоги субстратов, токсины, лекарственные средства, металлы) могут ингибировать ферменты, понижая
- 20. Обратимые и необратимые ингибиторы Обратимые, с образованием EI комплекса, который быстро диссоциирует. Фермент угнетен только когда
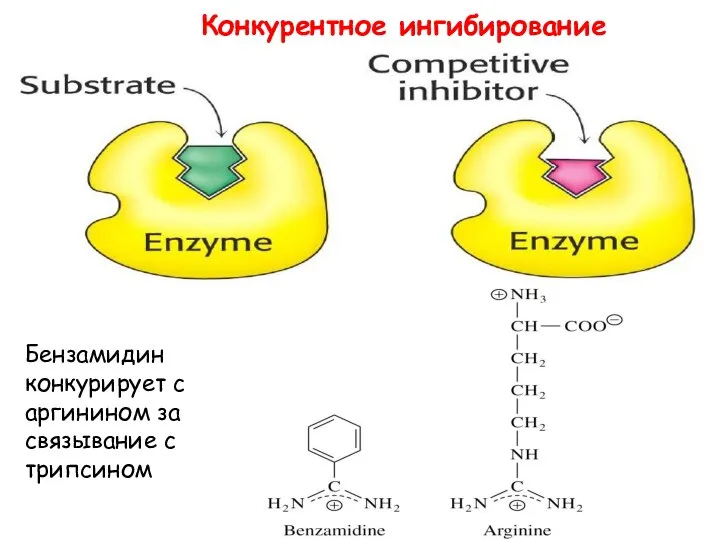
- 21. Конкурентное •Ингибитор имеет схожую с субстратом структуру, поэтому связывается с тем же активным центром •Фермент не
- 22. Конкурентное ингибирование Бензамидин конкурирует с аргинином за связывание с трипсином
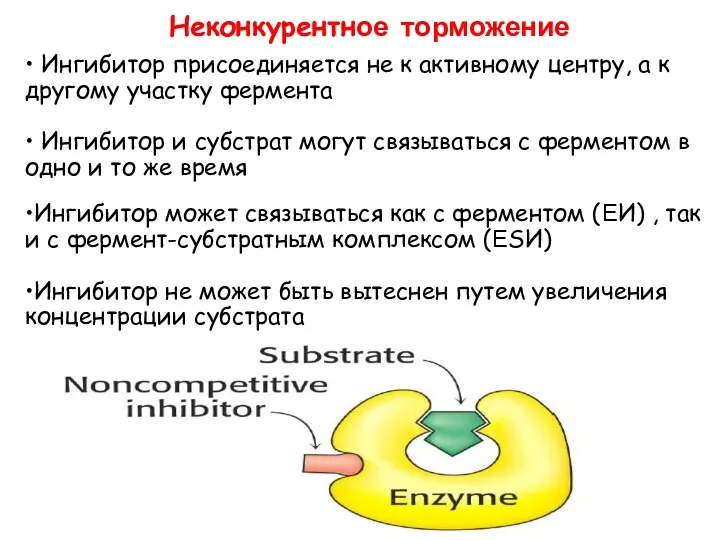
- 23. • Ингибитор присоединяется не к активному центру, а к другому участку фермента • Ингибитор и субстрат
- 24. Безконкурентное торможение Ингибитор присоединяется к комплексу фермент-субстрат (ЕS), но не к свободному ферменту Е.
- 25. Необратимое ингибирование Очень медленная дисоциация комплекса EI Связываются ковалентными связями с ферментом. Необратимые ингибиторы •ингибиторы специфические
- 26. Ингибиторы специфические к группам аминокислотных остатков - взаимодействуют со специфическими R группами аминокислот
- 27. Аналоги субстратов – структурно похожи на субстрат фермента - ковалентно модифицируют активный центр
- 28. •Ингибитор связывается как субстрат и сначала инициирует нормальный каталитический механизм •Потом образуются химически реактивные соединения, которые
- 29. Механизм ингибирования можно определить из графика зависимости скорости реакции от концентрации субстрата
- 30. Термин «аллостерический» происходит от греческих слов allo – другой, stereo – твердый (относящийся к трехмерной структуре,
- 31. Аллостерические ферменты
- 32. Аллостерические ферменты
- 33. Регуляция Активности Ферментов • Аллостерическая регуляция • Обратимая ковалентная модификация • Изоферменты • Протеолитическая активация Методы
- 34. Аллостерические модуляторы связываются нековалентно с аллостерическим центром. - регулируют активность фермента изменяя его конформацию. Аллостерические ферменты
- 35. Регуляция активности ферментов путем ковалентной модификации Ковалентное присоединение молекулы к аминокислотному остатку фермента может модифицировать активность
- 36. Фосфорилирование
- 37. Реакция дефосфорилирования Как правило, фосфорилированные ферменты более активные Ферменты, ответственные за фосфорилирование - протеинкиназы Ферменты, ответственные
- 38. множесственные формы фермента, которые отличаются аминокислотной последовательностью, но катализируют ту же реакцию. Изоферменты могут отличаться: кинетической
- 39. Анализ концентрации изоферментов в крови важен для диагностики различных болезней. Есть 5 изоферментов ЛДГ: H4 –
- 40. Протеолитическая активация • Много ферментов синтезируются как неактивные предшественники (зимогены) и активируются протеолитическим расщеплением Примеры специфического
- 42. Полиферментные комплексы - разные ферменты, которые катализируют последовательные реакции одного процесса и пространственно размещаются в одном
- 44. Введение в обмен веществ. Специфические и общие пути превращения углеводов, липидов и белков (окислительное декарбоксилирование ПВК,
- 45. Метаболизм – химические реакции, которые проходят в организме Метаболиты – маленькие промежуточные молекулы, которые образуются в
- 46. (a) Линейными (b) Циклическими (c) Спиральными (синтез жирных кислот) Последовательность реакций, которые имеют цель (например, расщепление
- 47. Катаболические реакции – деградация больших молекул с образованиемм меньших и энергии Анаболические реакции – синтез макромолекул
- 48. Регуляция метаболических путей Уровни регуляции метаболизма Нервная система Эндокринная система Взаимодействие между органами Клеточный (мембранный) уровень
- 49. Стадии метаболизма Катаболизм Стадия I (специфическая). Деградация макромолекул (белков, углеводов, липидов) к мономерам Стадия II (специфическая).
- 50. ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
- 51. Глюкоза Пируват Гликолиз Глицерол Амино- кислоты Ацетил CoA
- 52. Транспорт пирувата в митохондрию
- 53. Пируватдегидрогеназный комплекс - поли-ферментный комплекс, который состоит с 3 ферментов, 5 коферментов Превращение пирувата в ацетил
- 54. Ферменты: E1 = пируватдегидрогеназа E2 = дигидролипоил ацетилтрансфераза E3 = дигидролипоил дегидрогеназа Коферменты: ТПФ (тиамин пирофосфат),
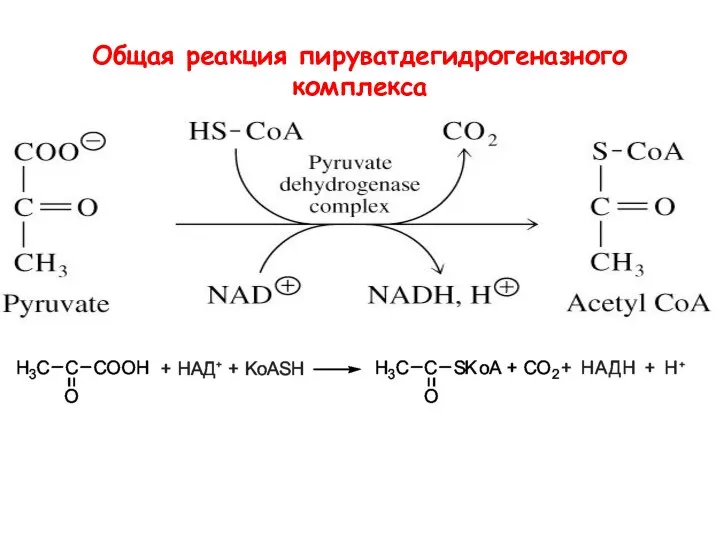
- 55. Общая реакция пируватдегидрогеназного комплекса
- 56. Названия: Цикл трикарбоновых кислот Цикл лимонной кислоты Цикл Кребса Ганс Адольф Кребс, выдающийся биохимик, родился в
- 57. У эукариотов все реакции цикла Кребса проходят в матриксе митохондрий
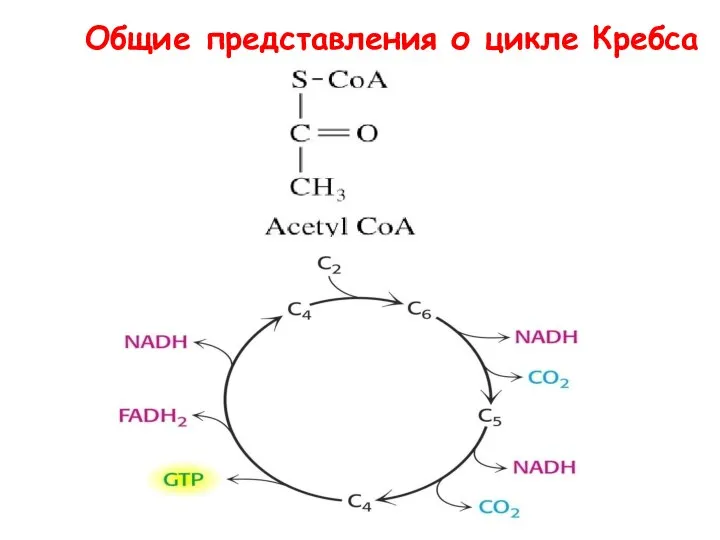
- 58. Общие представления о цикле Кребса
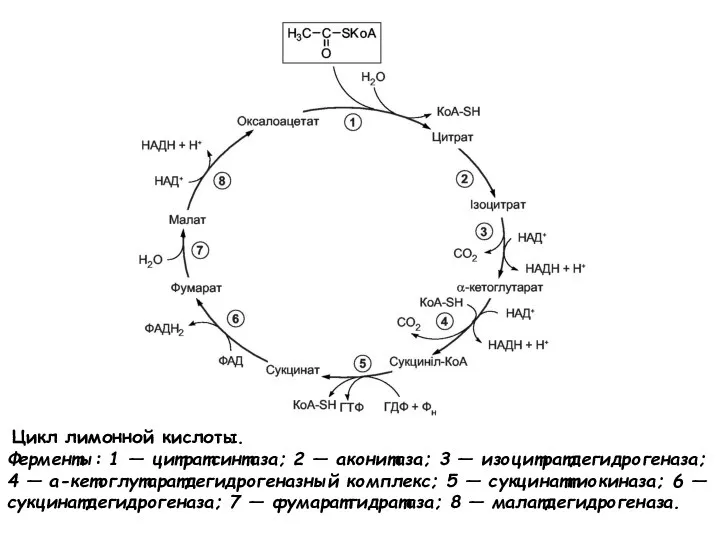
- 59. Цикл лимонной кислоты. Ферменты: 1 — цитратсинтаза; 2 — аконитаза; 3 — изоцитратдегидрогеназа; 4 — а-кетоглутаратдегидрогеназный
- 61. Скачать презентацию
Слайд 2Клеточная организация ферментативной активности
Ферменты располагаются в субклеточных структурах (органеллах) соответственно их функциям.
Клеточная организация ферментативной активности
Ферменты располагаются в субклеточных структурах (органеллах) соответственно их функциям.

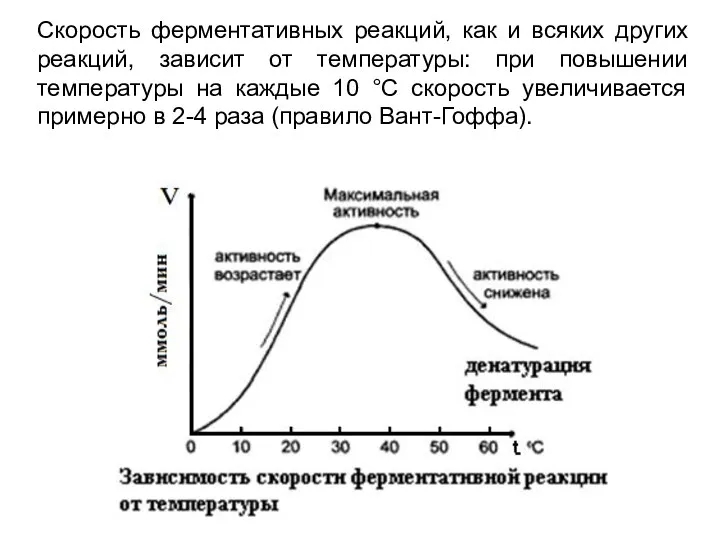
Слайд 3Скорость ферментативных реакций, как и всяких других реакций, зависит от температуры: при
Скорость ферментативных реакций, как и всяких других реакций, зависит от температуры: при
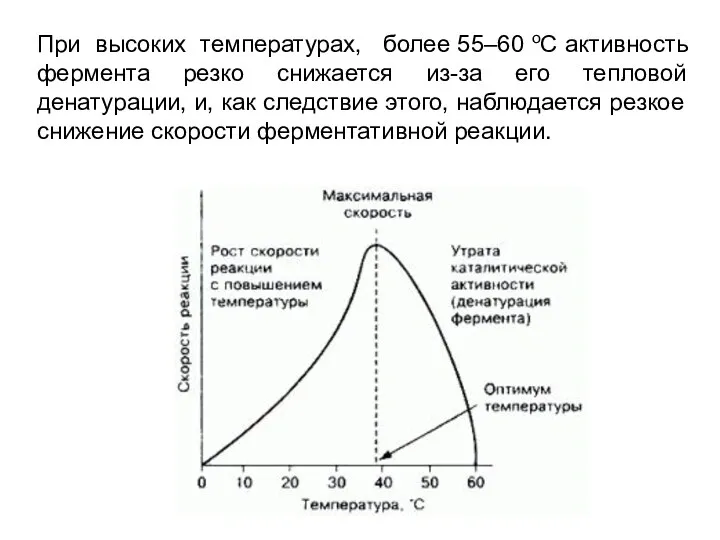
Слайд 4При высоких температурах, более 55–60 оС активность фермента резко снижается из-за его
При высоких температурах, более 55–60 оС активность фермента резко снижается из-за его
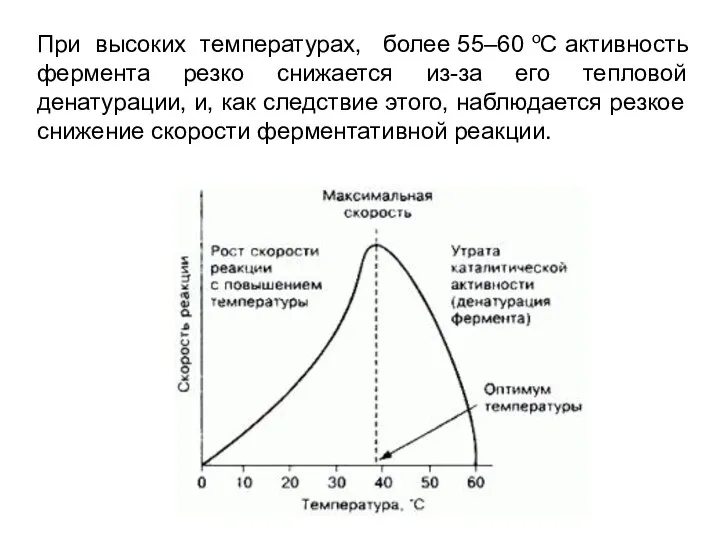
Слайд 5При высоких температурах, более 55–60 оС активность фермента резко снижается из-за его
При высоких температурах, более 55–60 оС активность фермента резко снижается из-за его
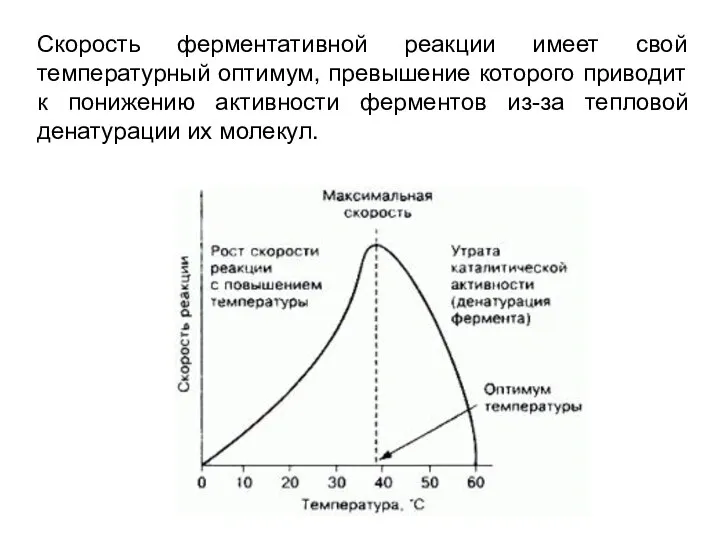
Слайд 6Скорость ферментативной реакции имеет свой температурный оптимум, превышение которого приводит к понижению
Скорость ферментативной реакции имеет свой температурный оптимум, превышение которого приводит к понижению
Слайд 7Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает
Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает
Слайд 8Каждый фермент имеет свой рН–оптимум - значение рН, при котором его активность
Каждый фермент имеет свой рН–оптимум - значение рН, при котором его активность
Слайд 9Большинство ферментов наиболее активны при рН=6-8. Исключения - пепсин (рНопт=1,5-2), аргиназа (рНопт=10-11).
Большинство ферментов наиболее активны при рН=6-8. Исключения - пепсин (рНопт=1,5-2), аргиназа (рНопт=10-11).
Слайд 10Простейшая кинетическая схема взаимодействия фермента (Е) и субстрата (S).
Во время реакции молекула
Простейшая кинетическая схема взаимодействия фермента (Е) и субстрата (S).
Во время реакции молекула

Слайд 11Впияние концентрации фермента на скорость реакции
При достаточной концентрации субстрата, чем выше
Впияние концентрации фермента на скорость реакции
При достаточной концентрации субстрата, чем выше
Слайд 12Влияние концентрации субстрата на скорость реакции
Влияние концентрации субстрата на скорость реакции
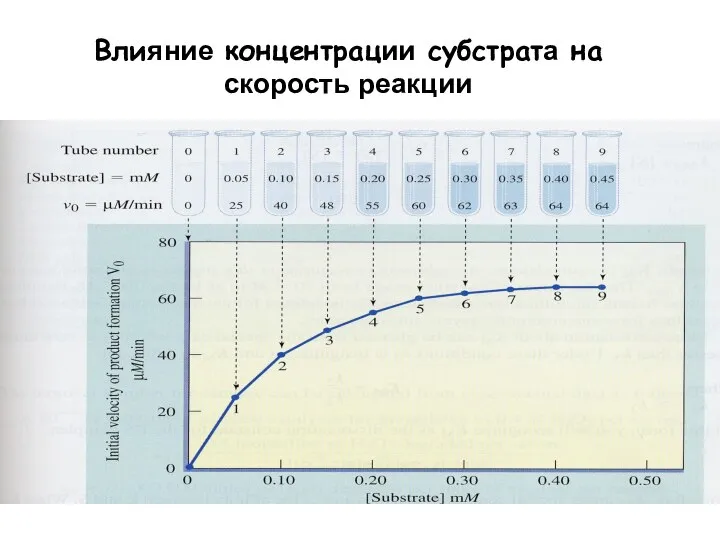
Слайд 13V – скорость реакции;
Vmax – максимальная скорость равная KcatE0;
[S] — концентрация
V – скорость реакции;
Vmax – максимальная скорость равная KcatE0;
[S] — концентрация
![V – скорость реакции; Vmax – максимальная скорость равная KcatE0; [S] —](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/959185/slide-12.jpg)
Слайд 14 При фиксированной концентрации фермента начальная скорость реакции линейно пропорциональна концентрации субстрата,
При фиксированной концентрации фермента начальная скорость реакции линейно пропорциональна концентрации субстрата,

Слайд 15 Уравнение Михаэлиса — Ментен
V – скорость реакции;
Vmax – максимальная скорость
Уравнение Михаэлиса — Ментен
V – скорость реакции;
Vmax – максимальная скорость
Слайд 16Скорость катализа
Уравнение Михаэлиса—Ментен — основное уравнение ферментативной кинетики, описывает зависимость скорости реакции,
Скорость катализа
Уравнение Михаэлиса—Ментен — основное уравнение ферментативной кинетики, описывает зависимость скорости реакции,

Слайд 17Уравнение Лайнуивера-Бёрке
Уравнение Лайнуивера-Бёрке
Слайд 18Когда [S] много больше чем Km скорость реакции не зависит от [S]
Когда [S] много больше чем Km скорость реакции не зависит от [S]
![Когда [S] много больше чем Km скорость реакции не зависит от [S]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/959185/slide-17.jpg)
Слайд 19Ингибирование ферментов
Разные химические агенты (метаболиты, аналоги субстратов, токсины, лекарственные средства, металлы) могут
Ингибирование ферментов
Разные химические агенты (метаболиты, аналоги субстратов, токсины, лекарственные средства, металлы) могут

Слайд 20Обратимые и необратимые ингибиторы
Обратимые, с образованием EI комплекса, который быстро диссоциирует.
Обратимые и необратимые ингибиторы
Обратимые, с образованием EI комплекса, который быстро диссоциирует.

Слайд 21Конкурентное
•Ингибитор имеет схожую с субстратом структуру, поэтому связывается с тем же активным
Конкурентное
•Ингибитор имеет схожую с субстратом структуру, поэтому связывается с тем же активным

Слайд 22Конкурентное ингибирование
Бензамидин конкурирует с аргинином за связывание с трипсином
Конкурентное ингибирование
Бензамидин конкурирует с аргинином за связывание с трипсином
Слайд 23• Ингибитор присоединяется не к активному центру, а к другому участку фермента
•
• Ингибитор присоединяется не к активному центру, а к другому участку фермента
•
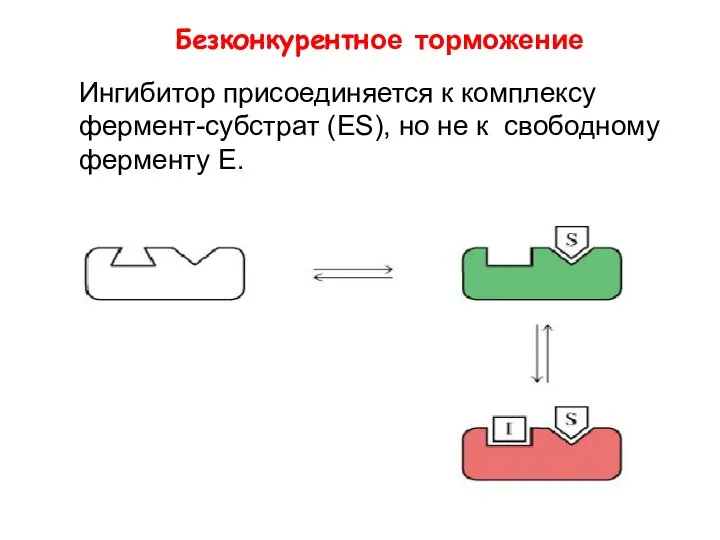
Слайд 24Безконкурентное торможение
Ингибитор присоединяется к комплексу фермент-субстрат (ЕS), но не к свободному ферменту
Безконкурентное торможение
Ингибитор присоединяется к комплексу фермент-субстрат (ЕS), но не к свободному ферменту
Слайд 25Необратимое ингибирование
Очень медленная дисоциация комплекса EI
Связываются ковалентными связями с ферментом.
Необратимые ингибиторы
•ингибиторы
Необратимое ингибирование
Очень медленная дисоциация комплекса EI
Связываются ковалентными связями с ферментом.
Необратимые ингибиторы
•ингибиторы

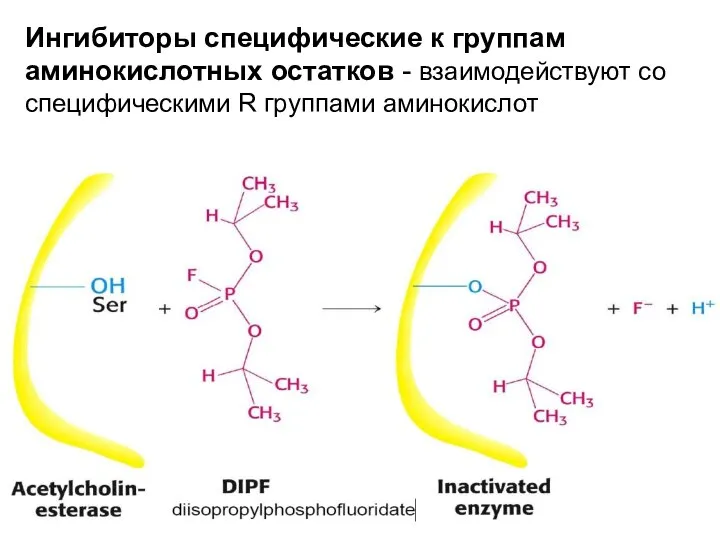
Слайд 26Ингибиторы специфические к группам аминокислотных остатков - взаимодействуют со специфическими R группами
Ингибиторы специфические к группам аминокислотных остатков - взаимодействуют со специфическими R группами
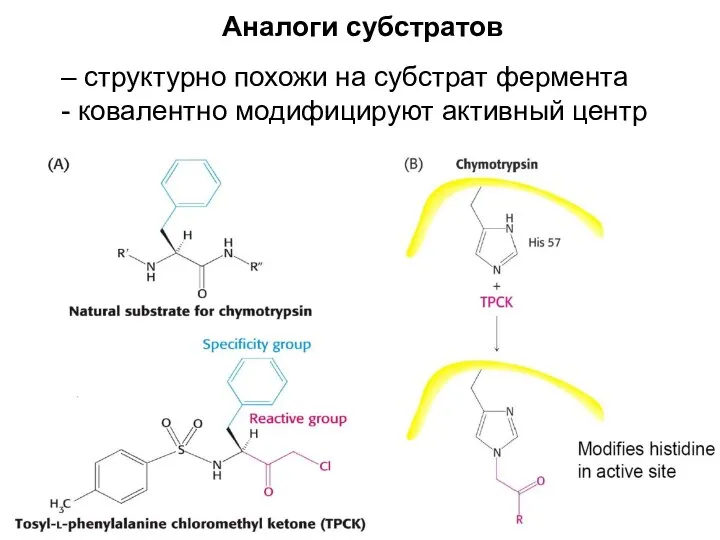
Слайд 27Аналоги субстратов
– структурно похожи на субстрат фермента
- ковалентно модифицируют активный центр
Аналоги субстратов
– структурно похожи на субстрат фермента
- ковалентно модифицируют активный центр
Слайд 28•Ингибитор связывается как субстрат и сначала инициирует нормальный каталитический механизм
•Потом образуются
•Ингибитор связывается как субстрат и сначала инициирует нормальный каталитический механизм
•Потом образуются

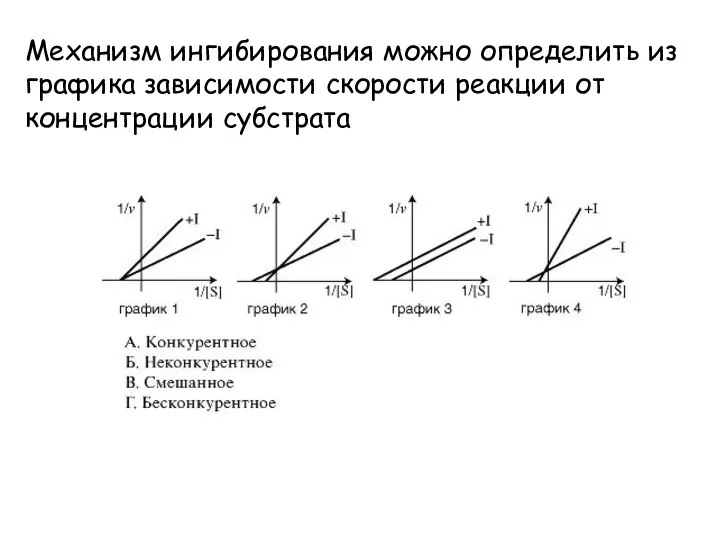
Слайд 29Механизм ингибирования можно определить из графика зависимости скорости реакции от концентрации субстрата
Механизм ингибирования можно определить из графика зависимости скорости реакции от концентрации субстрата
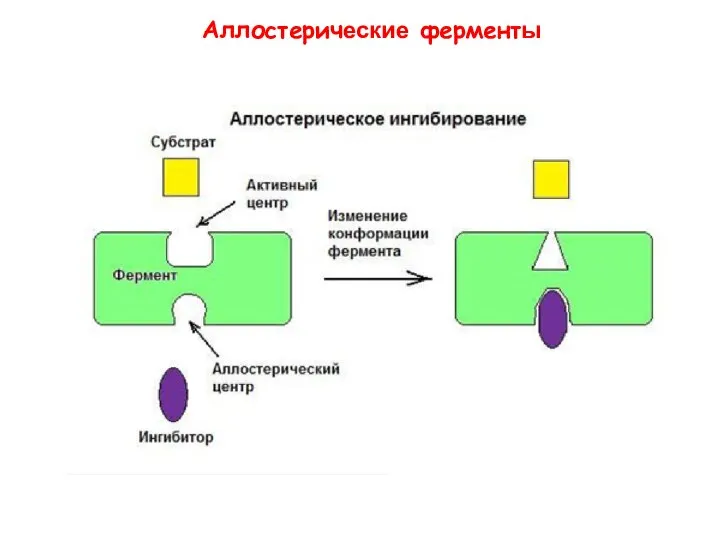
Слайд 30Термин «аллостерический» происходит от греческих слов allo – другой, stereo – твердый
Термин «аллостерический» происходит от греческих слов allo – другой, stereo – твердый

Слайд 31Аллостерические ферменты
Аллостерические ферменты
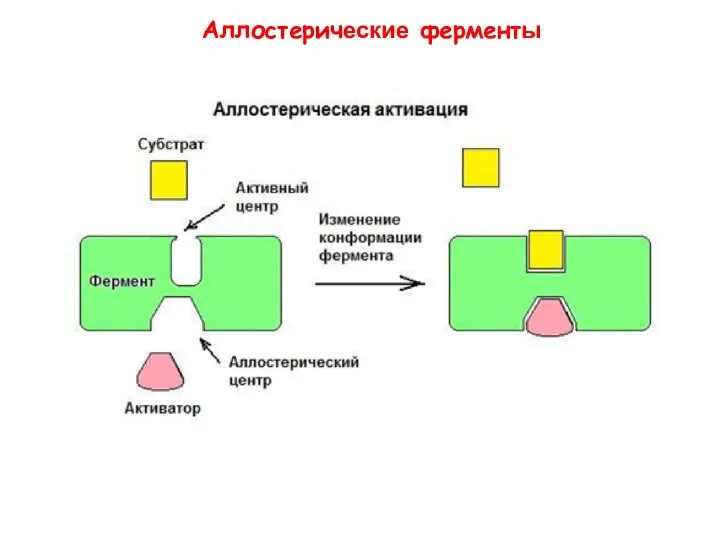
Слайд 32Аллостерические ферменты
Аллостерические ферменты
Слайд 33Регуляция Активности Ферментов
• Аллостерическая регуляция
• Обратимая ковалентная модификация
• Изоферменты
• Протеолитическая активация
Методы регуляции
Регуляция Активности Ферментов
• Аллостерическая регуляция
• Обратимая ковалентная модификация
• Изоферменты
• Протеолитическая активация
Методы регуляции

Слайд 34Аллостерические модуляторы
связываются нековалентно с аллостерическим центром.
- регулируют активность фермента изменяя его конформацию.
Аллостерические
Аллостерические модуляторы
связываются нековалентно с аллостерическим центром.
- регулируют активность фермента изменяя его конформацию.
Аллостерические

Слайд 35Регуляция активности ферментов путем ковалентной модификации
Ковалентное присоединение молекулы к аминокислотному остатку фермента
Регуляция активности ферментов путем ковалентной модификации
Ковалентное присоединение молекулы к аминокислотному остатку фермента

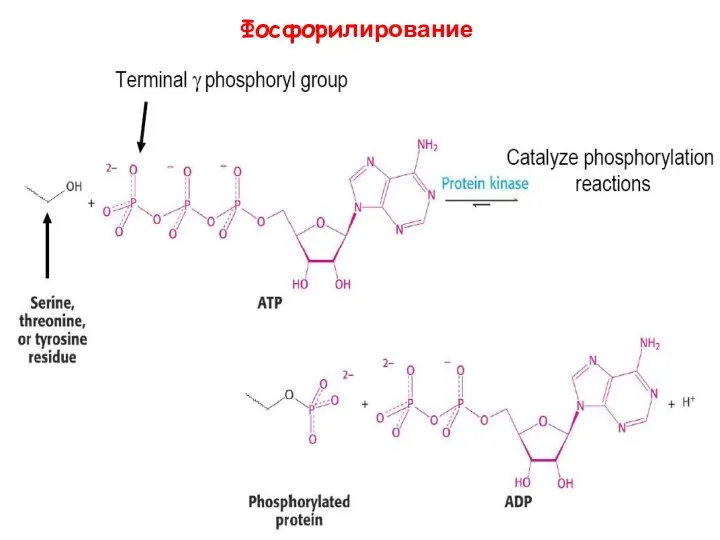
Слайд 36Фосфорилирование
Фосфорилирование
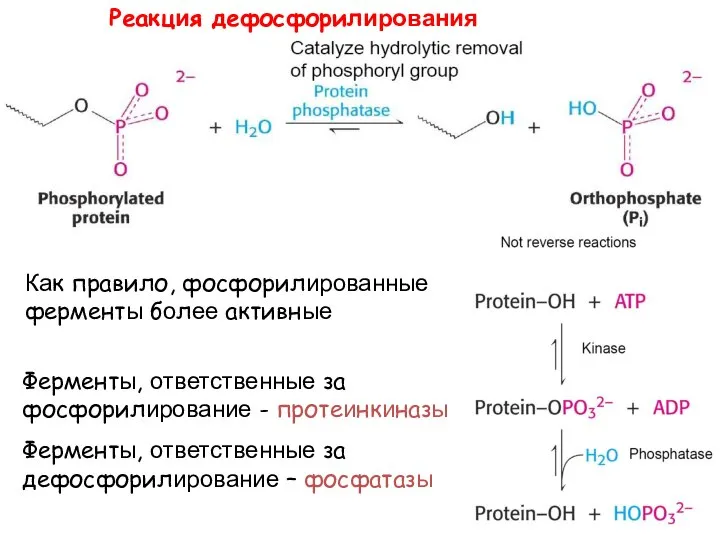
Слайд 37Реакция дефосфорилирования
Как правило, фосфорилированные ферменты более активные
Ферменты, ответственные за фосфорилирование -
Реакция дефосфорилирования
Как правило, фосфорилированные ферменты более активные
Ферменты, ответственные за фосфорилирование -
Слайд 38множесственные формы фермента, которые отличаются аминокислотной последовательностью, но катализируют ту же реакцию.
Изоферменты
множесственные формы фермента, которые отличаются аминокислотной последовательностью, но катализируют ту же реакцию.
Изоферменты

Слайд 39Анализ концентрации изоферментов в крови важен для диагностики различных болезней.
Есть 5 изоферментов
Анализ концентрации изоферментов в крови важен для диагностики различных болезней.
Есть 5 изоферментов

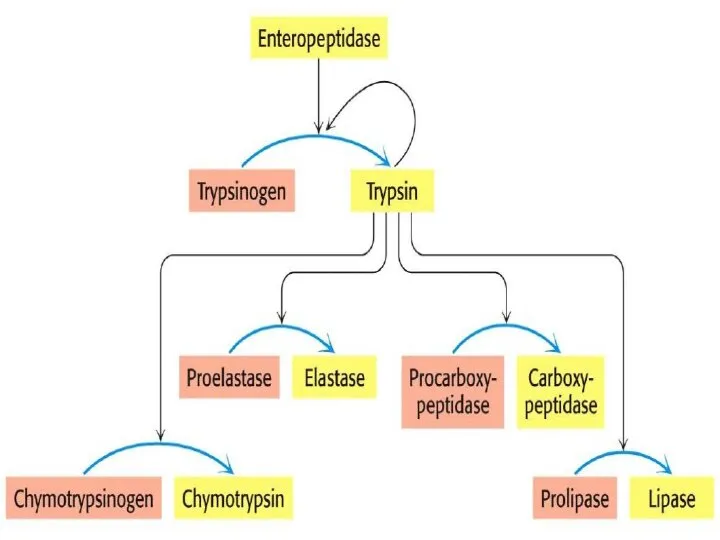
Слайд 40Протеолитическая активация
• Много ферментов синтезируются как неактивные предшественники (зимогены) и активируются протеолитическим
Протеолитическая активация
• Много ферментов синтезируются как неактивные предшественники (зимогены) и активируются протеолитическим

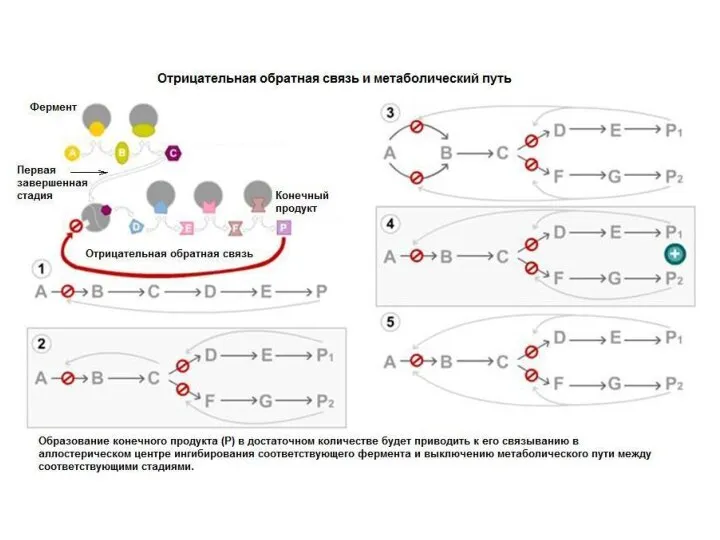
Слайд 42Полиферментные комплексы - разные ферменты, которые катализируют последовательные реакции одного процесса и
Полиферментные комплексы - разные ферменты, которые катализируют последовательные реакции одного процесса и

Слайд 44Введение в обмен веществ. Специфические и общие пути превращения углеводов, липидов и
Введение в обмен веществ. Специфические и общие пути превращения углеводов, липидов и

Слайд 45Метаболизм – химические реакции, которые проходят в организме
Метаболиты – маленькие промежуточные молекулы,
Метаболизм – химические реакции, которые проходят в организме
Метаболиты – маленькие промежуточные молекулы,
Слайд 46(a) Линейными (b) Циклическими
(c) Спиральными (синтез жирных кислот)
Последовательность реакций, которые имеют цель
(a) Линейными (b) Циклическими
(c) Спиральными (синтез жирных кислот)
Последовательность реакций, которые имеют цель
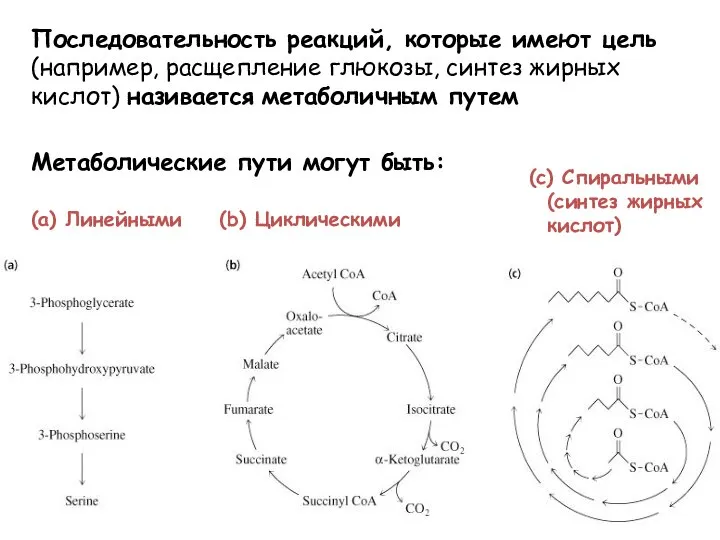
Слайд 47Катаболические реакции – деградация больших молекул с образованиемм меньших и энергии
Анаболические
Катаболические реакции – деградация больших молекул с образованиемм меньших и энергии
Анаболические

Слайд 48Регуляция метаболических путей
Уровни регуляции метаболизма
Нервная система
Эндокринная система
Взаимодействие между органами
Клеточный (мембранный) уровень
Молекулярный уровень
Регуляция метаболических путей
Уровни регуляции метаболизма
Нервная система
Эндокринная система
Взаимодействие между органами
Клеточный (мембранный) уровень
Молекулярный уровень

Слайд 49Стадии метаболизма
Катаболизм
Стадия I (специфическая). Деградация макромолекул (белков, углеводов, липидов) к мономерам
Стадия II
Стадии метаболизма
Катаболизм
Стадия I (специфическая). Деградация макромолекул (белков, углеводов, липидов) к мономерам
Стадия II

Слайд 50ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
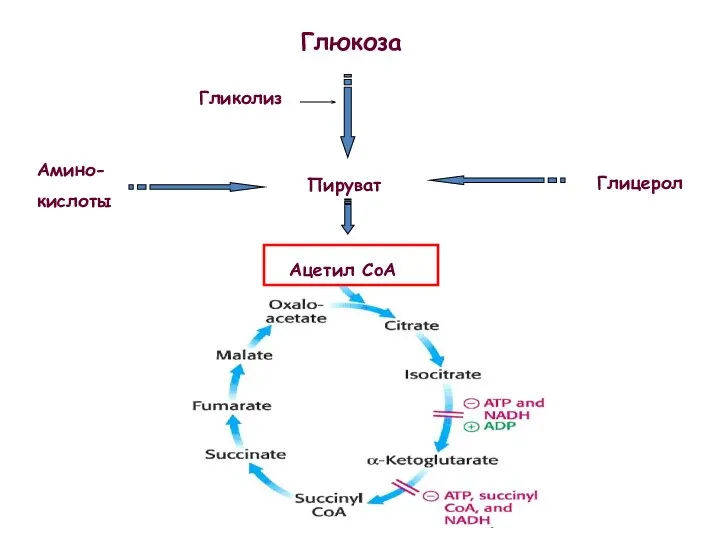
Слайд 51Глюкоза
Пируват
Гликолиз
Глицерол
Амино-
кислоты
Ацетил CoA
Глюкоза
Пируват
Гликолиз
Глицерол
Амино-
кислоты
Ацетил CoA
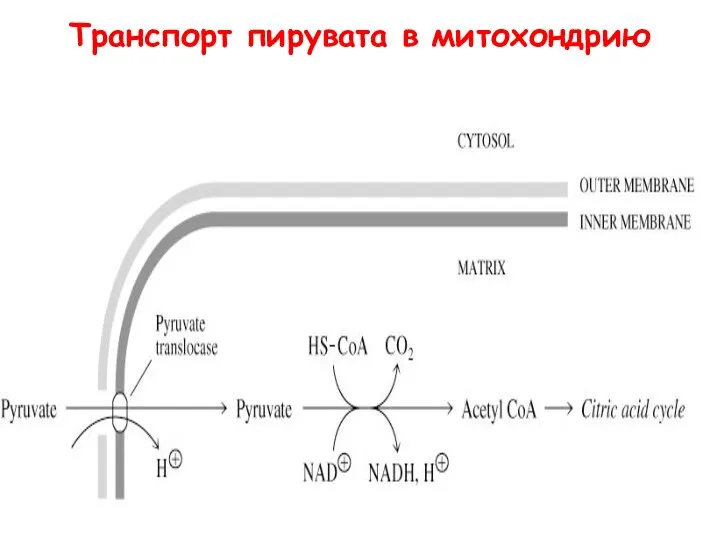
Слайд 52Транспорт пирувата в митохондрию
Транспорт пирувата в митохондрию
Слайд 53Пируватдегидрогеназный комплекс - поли-ферментный комплекс, который состоит с 3 ферментов, 5 коферментов
Пируватдегидрогеназный комплекс - поли-ферментный комплекс, который состоит с 3 ферментов, 5 коферментов
Слайд 54Ферменты:
E1 = пируватдегидрогеназа
E2 = дигидролипоил ацетилтрансфераза
E3 = дигидролипоил дегидрогеназа
Коферменты: ТПФ (тиамин пирофосфат),
Ферменты:
E1 = пируватдегидрогеназа
E2 = дигидролипоил ацетилтрансфераза
E3 = дигидролипоил дегидрогеназа
Коферменты: ТПФ (тиамин пирофосфат),

Слайд 55Общая реакция пируватдегидрогеназного комплекса
Общая реакция пируватдегидрогеназного комплекса
Слайд 56Названия:
Цикл трикарбоновых кислот
Цикл лимонной кислоты
Цикл Кребса
Ганс Адольф Кребс,
выдающийся биохимик, родился
Названия:
Цикл трикарбоновых кислот
Цикл лимонной кислоты
Цикл Кребса
Ганс Адольф Кребс,
выдающийся биохимик, родился

Слайд 57У эукариотов все реакции цикла Кребса проходят в матриксе митохондрий
У эукариотов все реакции цикла Кребса проходят в матриксе митохондрий
Слайд 58Общие представления о цикле Кребса
Общие представления о цикле Кребса
Слайд 59 Цикл лимонной кислоты.
Ферменты: 1 — цитратсинтаза; 2 — аконитаза; 3 —
Цикл лимонной кислоты.
Ферменты: 1 — цитратсинтаза; 2 — аконитаза; 3 —
 Введение в нейроанатомию. Развитие спинного и головного мозга. Классификация нервной системы: центральная и периферическая части
Введение в нейроанатомию. Развитие спинного и головного мозга. Классификация нервной системы: центральная и периферическая части Презентация на тему ОСОБЕННОСТИ РАСТИТЕЛЬНОЙ КЛЕТКИ
Презентация на тему ОСОБЕННОСТИ РАСТИТЕЛЬНОЙ КЛЕТКИ  Рабочая тетрадь. Биология 5 класс
Рабочая тетрадь. Биология 5 класс Биологическое ассорти
Биологическое ассорти Alpine salamander
Alpine salamander Роль белков
Роль белков Этиология речевых нарушений
Этиология речевых нарушений В мире цветов
В мире цветов Развитие животного мира на Земле
Развитие животного мира на Земле Лист — часть побега, внешнее и внутреннее строение листа. 7 класс
Лист — часть побега, внешнее и внутреннее строение листа. 7 класс Строение и функции кожного покрова
Строение и функции кожного покрова Строение среднего, промежуточного и концевого мозга. Оболочки и пространства, артерии и вены головного мозга
Строение среднего, промежуточного и концевого мозга. Оболочки и пространства, артерии и вены головного мозга Размножение
Размножение Черёмуха Маака. История школьного парка
Черёмуха Маака. История школьного парка Плоские черви. Урок 3
Плоские черви. Урок 3 054 урок Побеги и почки
054 урок Побеги и почки Морфология собственно-соединительных тканей
Морфология собственно-соединительных тканей Программа биологического мониторинга. (Тема 1)
Программа биологического мониторинга. (Тема 1) Растения Красной книги России
Растения Красной книги России Презентация на тему Пресмыкающиеся и их особенности
Презентация на тему Пресмыкающиеся и их особенности  Биочипирование. Чипы сегодня
Биочипирование. Чипы сегодня Біосинтез і біотрансформація холестеролу. Метаболізм кетонових тіл. Регуляція та порушення ліпідного обміну
Біосинтез і біотрансформація холестеролу. Метаболізм кетонових тіл. Регуляція та порушення ліпідного обміну Животные, истребленные человеком
Животные, истребленные человеком Влияние внешних и внутренних факторов на транспирацию
Влияние внешних и внутренних факторов на транспирацию Фотосинтез
Фотосинтез 20140107_zhivotnye_saratovskoy_oblasti
20140107_zhivotnye_saratovskoy_oblasti Теория сильных электролитов. Роль электролитов в жизнедеятельности организмов
Теория сильных электролитов. Роль электролитов в жизнедеятельности организмов Частная патология
Частная патология