- Коэволюция и эволюция экосистем

Содержание
- 2. Коэволюция Эволюция разных видов и групп огранизмов не происходит независимо друг от друга. Изменения внутри одного
- 3. Коэволюция хищника и жертвы Эволюция (например, дизруптивный отбор) может изменять стандартную схему колебаний между численностью хищника
- 4. Коэволюция хищника и жертвы В популяции, где присутствуют белки, размер шишек и толщина сосны больше, чем
- 5. Коэволюция паразита и хозяина Степень патогенности Устойчивость хозяина
- 6. Коэволюция паразита и хозяина В линиях паразита и хозяина видообразование может идти конгруэнтно (филогения паразитов =
- 7. Гипотеза Чёрной королевы (Red Queen hypothesis) Leigh Van Valen (1935 — 2010) Способность вида к выживанию
- 8. Обманщики и мутуалисты Бабочки откладывают небольшое число яиц в завязь цветка юкки, при этом опыляя цветок.
- 9. Коэволюция при мутуализме Благодаря питанию с помощью симбионтов у тлей стали нерабочими многие гены, участвующие в
- 10. Коэволюция при мутуализме Устойчивая передача симбионта от родителей к потомкам выравнивает уровни приспособленности обоих организмов, тем
- 11. Гильдии и конкуренция Гильдия — группа видов, использующая одни и те же ресурсы, занимающих сходные экологические
- 12. Классическая теория биоразнообразия
- 13. Нейтральная теория биоразнообразия Stephen Hubbell (род. 1942)
- 14. Изменения экосистем Специогенез — изменение структуры экосистемы за счет видообразования Экогенез — изменение структуры экосистемы за
- 15. Мелиттофильный комплекс Микромийофильный комплекс Комплекс растений с широким кругом опылителей Коадаптивные комплексы – комплексы видов, в
- 16. Совымирание
- 17. Court Jester vs Red Queen
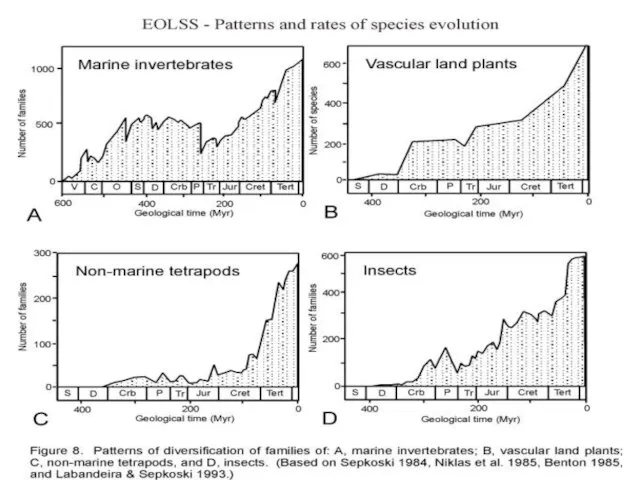
- 18. Джон Сепкоски Jack John Sepkoski (1948 – 1999) Массовые вымирания в истории жизни на земле: 0.
- 19. 0 1 2
- 20. 3 4 5
- 21. Лазарь (Элвис) Эндемик Исчезновение Появление Проходящий
- 22. Импактные гипотезы
- 23. Импактные гипотезы
- 24. Некоторые группы начинали регрессировать задолго до кризиса, а другие продолжали существовать и после него
- 26. «Экологические» гипотезы
- 27. Теория филоценогенеза Владимир Васильевич Жерихин (1945 — 2001) По теории филоценогенеза единицей эволюции экосистем является сукцессионный
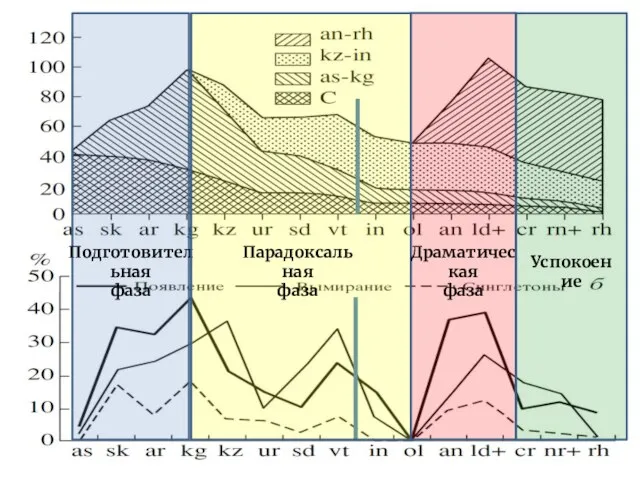
- 28. Теория экологических кризисов Владимир Васильевич Жерихин (1945 — 2001) Александр Павлович Расницын (р. 1936) Поскольку: а)
- 30. Теория экологических кризисов Подготовительная фаза В ходе подготовительной фазы происходит дробление адаптивных зон, рост числа специалистов,
- 31. Теория экологических кризисов III. Драматическая фаза Интенсификация появлений и вымираний новых таксонов, причём появления преобладают над
- 33. Скачать презентацию
Слайд 3Коэволюция хищника и жертвы
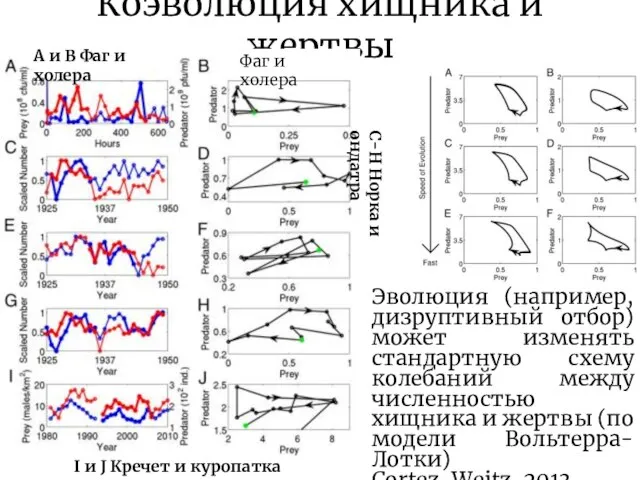
Эволюция (например, дизруптивный отбор) может изменять стандартную схему колебаний
Коэволюция хищника и жертвы
Эволюция (например, дизруптивный отбор) может изменять стандартную схему колебаний
Слайд 4Коэволюция хищника и жертвы
В популяции, где присутствуют белки, размер шишек и толщина
Коэволюция хищника и жертвы
В популяции, где присутствуют белки, размер шишек и толщина

Слайд 5Коэволюция паразита и хозяина
Степень патогенности
Устойчивость хозяина
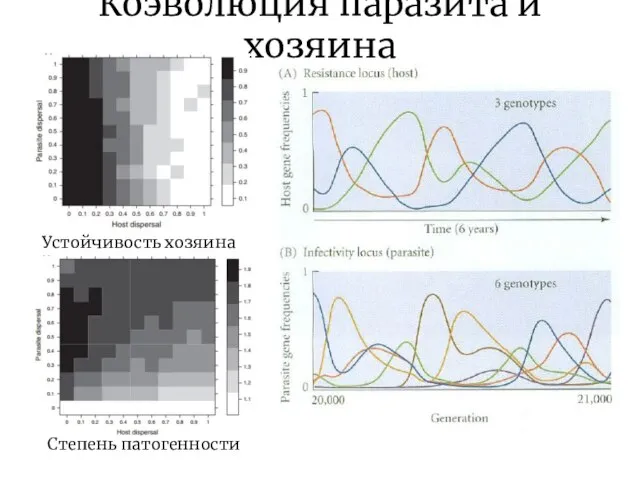
Коэволюция паразита и хозяина
Степень патогенности
Устойчивость хозяина
Слайд 6Коэволюция паразита и хозяина
В линиях паразита и хозяина видообразование может идти конгруэнтно
Коэволюция паразита и хозяина
В линиях паразита и хозяина видообразование может идти конгруэнтно

Слайд 7Гипотеза Чёрной королевы (Red Queen hypothesis)
Leigh Van Valen
(1935 — 2010)
Способность вида к
Гипотеза Чёрной королевы (Red Queen hypothesis)
Leigh Van Valen
(1935 — 2010)
Способность вида к

Слайд 8Обманщики и мутуалисты
Бабочки откладывают небольшое число яиц в завязь цветка юкки, при
Обманщики и мутуалисты
Бабочки откладывают небольшое число яиц в завязь цветка юкки, при

Слайд 9Коэволюция при мутуализме
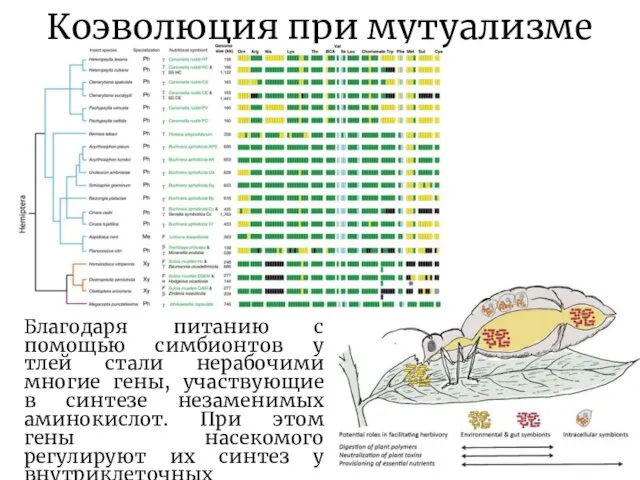
Благодаря питанию с помощью симбионтов у тлей стали нерабочими многие
Коэволюция при мутуализме
Благодаря питанию с помощью симбионтов у тлей стали нерабочими многие
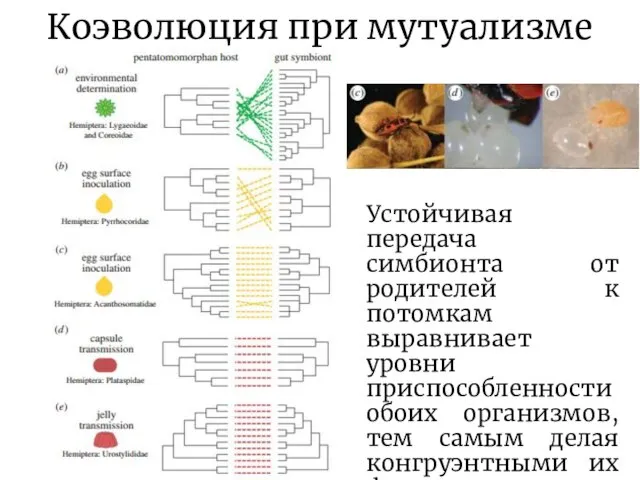
Слайд 10Коэволюция при мутуализме
Устойчивая передача симбионта от родителей к потомкам выравнивает уровни приспособленности
Коэволюция при мутуализме
Устойчивая передача симбионта от родителей к потомкам выравнивает уровни приспособленности
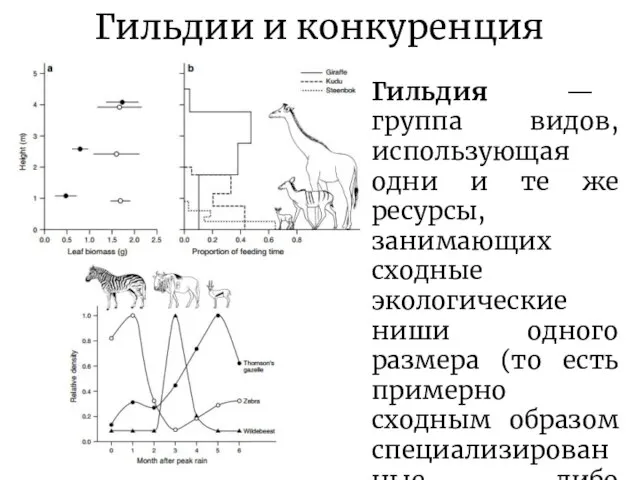
Слайд 11Гильдии и конкуренция
Гильдия — группа видов, использующая одни и те же ресурсы,
Гильдии и конкуренция
Гильдия — группа видов, использующая одни и те же ресурсы,
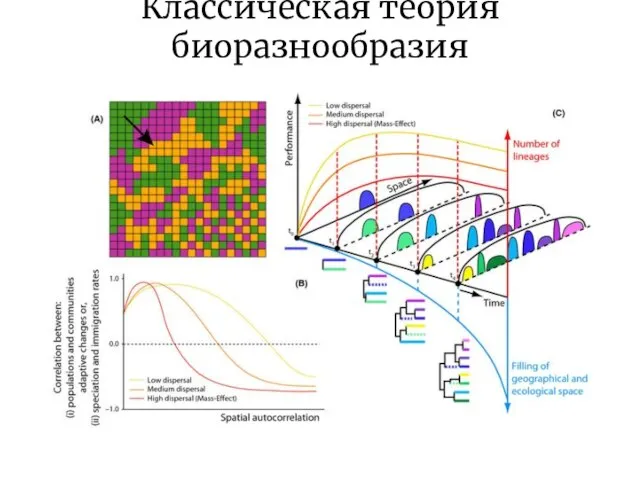
Слайд 12Классическая теория биоразнообразия
Классическая теория биоразнообразия
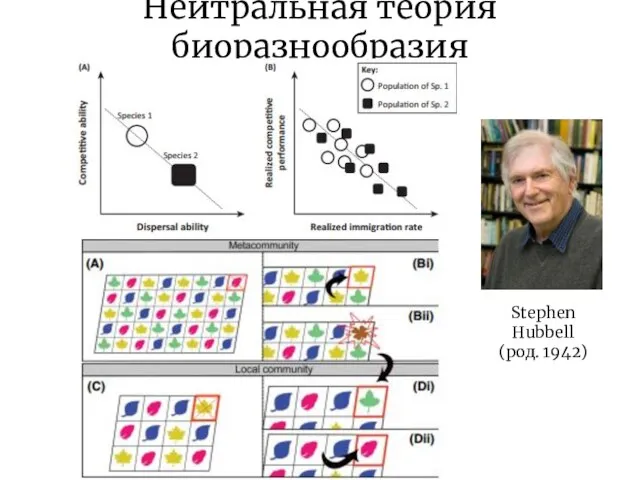
Слайд 13Нейтральная теория биоразнообразия
Stephen Hubbell
(род. 1942)
Нейтральная теория биоразнообразия
Stephen Hubbell
(род. 1942)
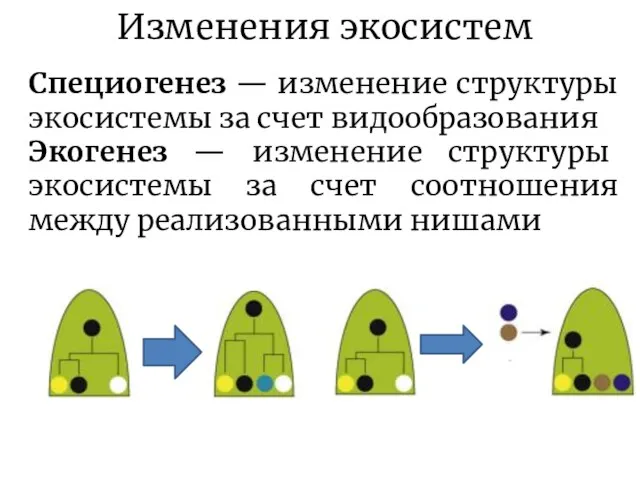
Слайд 14Изменения экосистем
Специогенез — изменение структуры экосистемы за счет видообразования
Экогенез — изменение структуры
Изменения экосистем
Специогенез — изменение структуры экосистемы за счет видообразования
Экогенез — изменение структуры
Слайд 15Мелиттофильный комплекс
Микромийофильный комплекс
Комплекс растений с широким кругом опылителей
Коадаптивные комплексы – комплексы видов,
Мелиттофильный комплекс
Микромийофильный комплекс
Комплекс растений с широким кругом опылителей
Коадаптивные комплексы – комплексы видов,

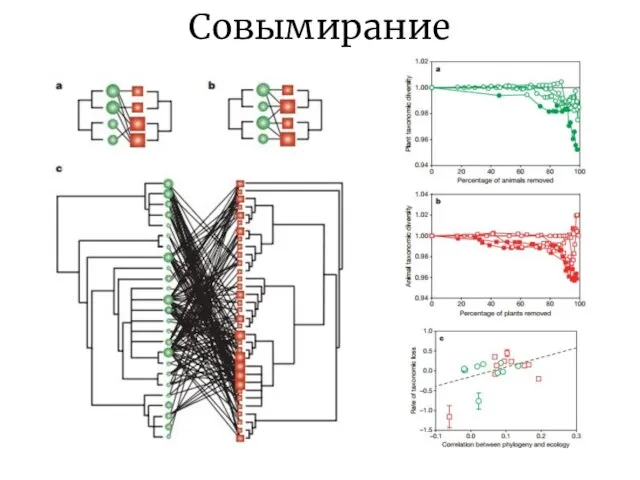
Слайд 16Совымирание
Совымирание
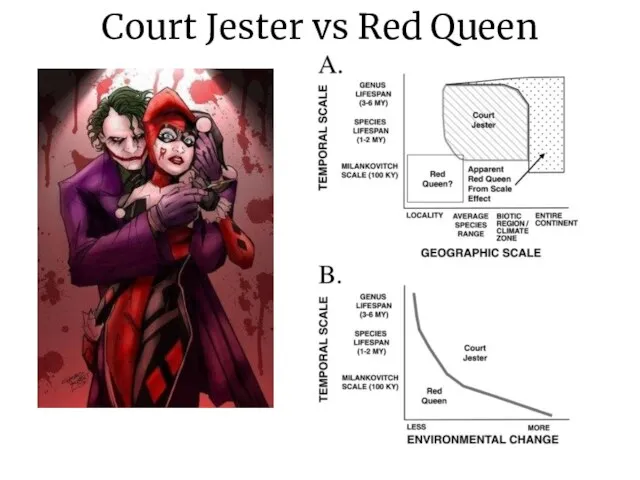
Слайд 17Court Jester vs Red Queen
Court Jester vs Red Queen
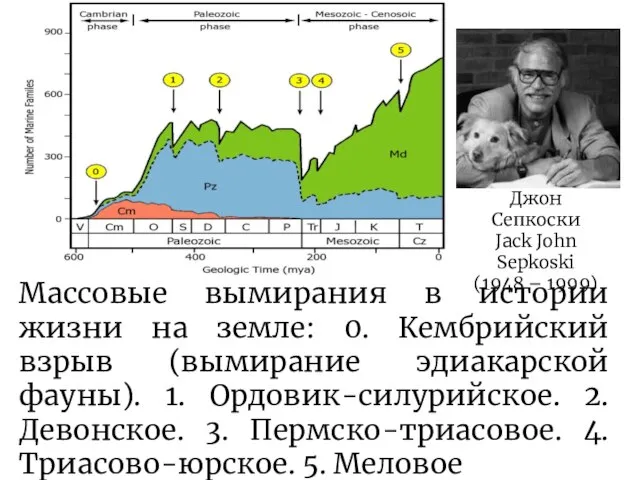
Слайд 18Джон Сепкоски
Jack John Sepkoski
(1948 – 1999)
Массовые вымирания в истории жизни на земле:
Джон Сепкоски
Jack John Sepkoski
(1948 – 1999)
Массовые вымирания в истории жизни на земле:
Слайд 190
1
2
0
1
2

Слайд 203
4
5
3
4
5

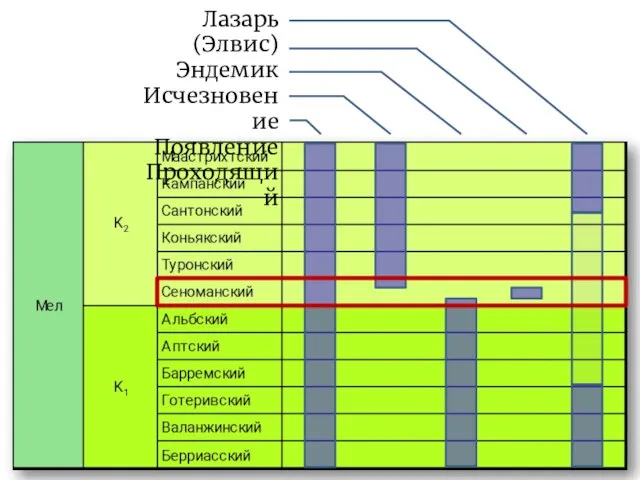
Слайд 21Лазарь (Элвис)
Эндемик
Исчезновение
Появление
Проходящий
Лазарь (Элвис)
Эндемик
Исчезновение
Появление
Проходящий
Слайд 22Импактные гипотезы
Импактные гипотезы

Слайд 23Импактные гипотезы
Импактные гипотезы

Слайд 24Некоторые группы начинали регрессировать задолго до кризиса, а другие продолжали существовать и
Некоторые группы начинали регрессировать задолго до кризиса, а другие продолжали существовать и

Слайд 26«Экологические» гипотезы
«Экологические» гипотезы

Слайд 27Теория филоценогенеза
Владимир Васильевич Жерихин
(1945 — 2001)
По теории филоценогенеза единицей эволюции экосистем является
Теория филоценогенеза
Владимир Васильевич Жерихин
(1945 — 2001)
По теории филоценогенеза единицей эволюции экосистем является

Слайд 28Теория экологических кризисов
Владимир Васильевич Жерихин
(1945 — 2001)
Александр Павлович Расницын
(р. 1936)
Поскольку:
а) массовые вымирания
Теория экологических кризисов
Владимир Васильевич Жерихин
(1945 — 2001)
Александр Павлович Расницын
(р. 1936)
Поскольку:
а) массовые вымирания

Слайд 30Теория экологических кризисов
Подготовительная фаза
В ходе подготовительной фазы происходит дробление адаптивных зон,
Теория экологических кризисов
Подготовительная фаза
В ходе подготовительной фазы происходит дробление адаптивных зон,

Слайд 31Теория экологических кризисов
III. Драматическая фаза
Интенсификация появлений и вымираний новых таксонов, причём появления
Теория экологических кризисов
III. Драматическая фаза
Интенсификация появлений и вымираний новых таксонов, причём появления

 Презентация на тему Подводный мир Черного моря
Презентация на тему Подводный мир Черного моря  Bloodreader. Персональный контролер биохимических параметров крови
Bloodreader. Персональный контролер биохимических параметров крови Молекулярно-генетический уровень жизни
Молекулярно-генетический уровень жизни Презентация на тему Биосинтез белка
Презентация на тему Биосинтез белка  Строение нервоной системы
Строение нервоной системы Тема 3
Тема 3 Презентация на тему Класс Земноводные 7 класс
Презентация на тему Класс Земноводные 7 класс  Гормональная регуляция обменных процессов: белков, воды, натрия, калия, кальция, фосфатов. Лекция № 22
Гормональная регуляция обменных процессов: белков, воды, натрия, калия, кальция, фосфатов. Лекция № 22 Coraux d’Hawaïï
Coraux d’Hawaïï Птицы Южного Урала
Птицы Южного Урала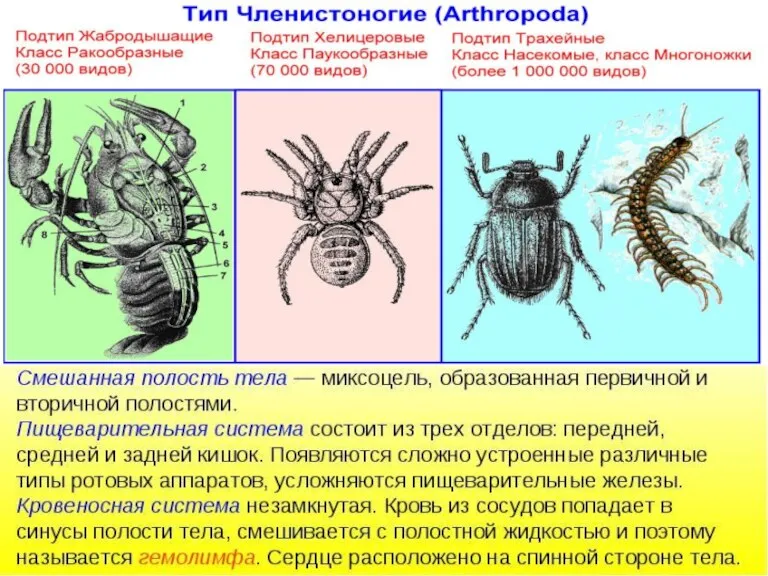 Тип Членистоногие
Тип Членистоногие Грегор Иоганн Мендель (1822 -1884). Законы генетики
Грегор Иоганн Мендель (1822 -1884). Законы генетики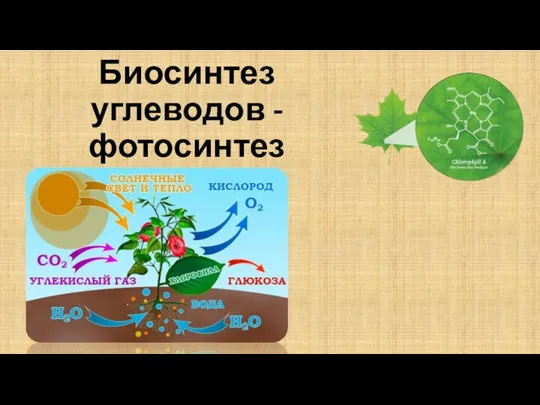 06. Биосинтез углеводов – фотосинтез
06. Биосинтез углеводов – фотосинтез Бактерии
Бактерии Охота в Рязанских лесах
Охота в Рязанских лесах Строение семян. Изучение строение семян двудольных растений
Строение семян. Изучение строение семян двудольных растений Индивидуальное развитие организма
Индивидуальное развитие организма Русский лес в кадре
Русский лес в кадре Васкулогенез и ангиогенез
Васкулогенез и ангиогенез Спинной и спинномозговые нервы
Спинной и спинномозговые нервы Растения субтропических областей Земного шара
Растения субтропических областей Земного шара Лекарственные растения
Лекарственные растения Пищеварительная система. Зубы, строение и функция
Пищеварительная система. Зубы, строение и функция Пингвины. Выразительность материалов для работы в объеме
Пингвины. Выразительность материалов для работы в объеме Поиск оптимальных условий непосредственно в процессе выращивания микроорганизмов
Поиск оптимальных условий непосредственно в процессе выращивания микроорганизмов Организм человека
Организм человека Транскрипція та реплікація
Транскрипція та реплікація Фотосинтез. Игра
Фотосинтез. Игра