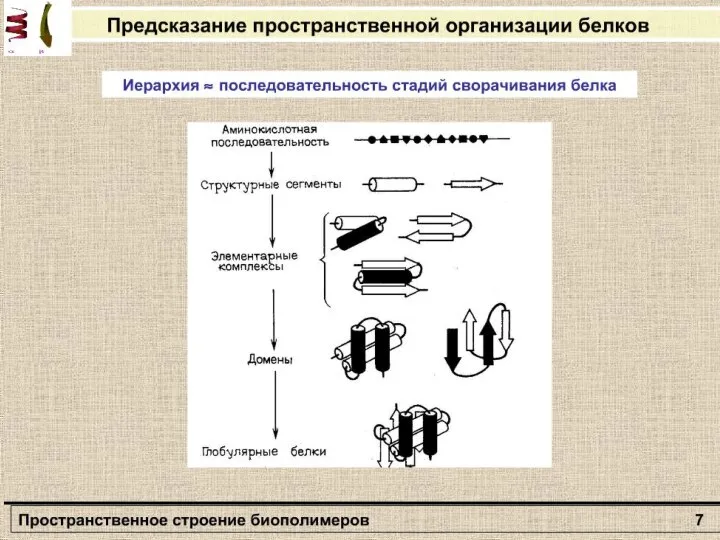
- Структура белка

Содержание
- 2. Конформационная энергия полипептида Общее строение полипептидной цепи, характеризуется тремя углами:
- 3. Пептидная связь – ω - угол Однако, двойной характер пептидной связи препятствует вращению вокруг нее: ω=const=180°.
- 4. 1. Компланарность – все атомы, входящие в пептидную группу, находятся в одной плоскости. 2. Способность к
- 5. Пространственная конфигурация полипептидной цепи Таким образом, пространственная конфигурация полипептидной цепи будет зависеть от кооперативного взаимодействия φ
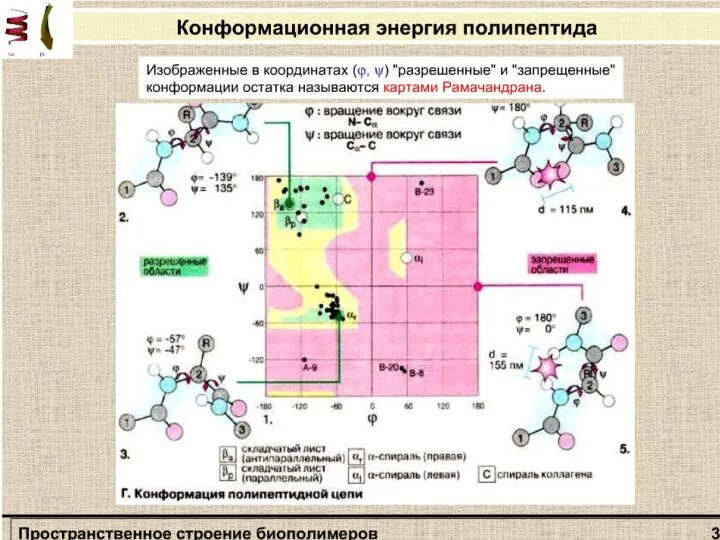
- 6. Не все значения углов ψ и φ допустимы вследствие возможного стерического несоответствия двух соседних аминокислотных остатков.
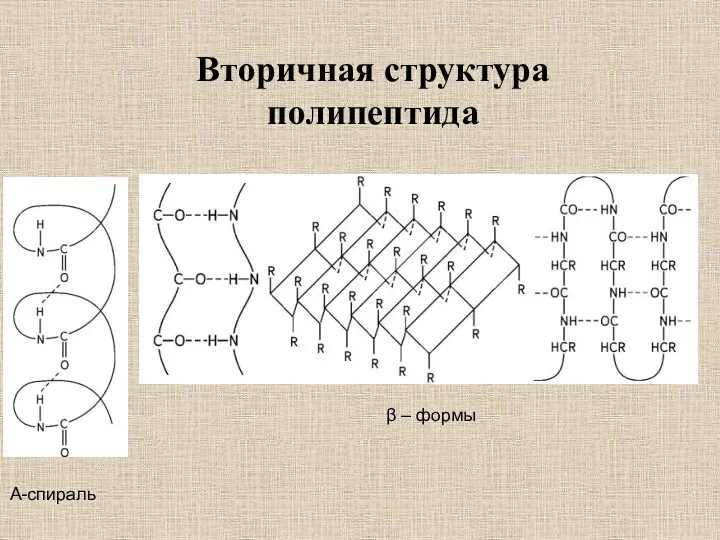
- 12. Вторичная структура полипептида Α-спираль β – формы
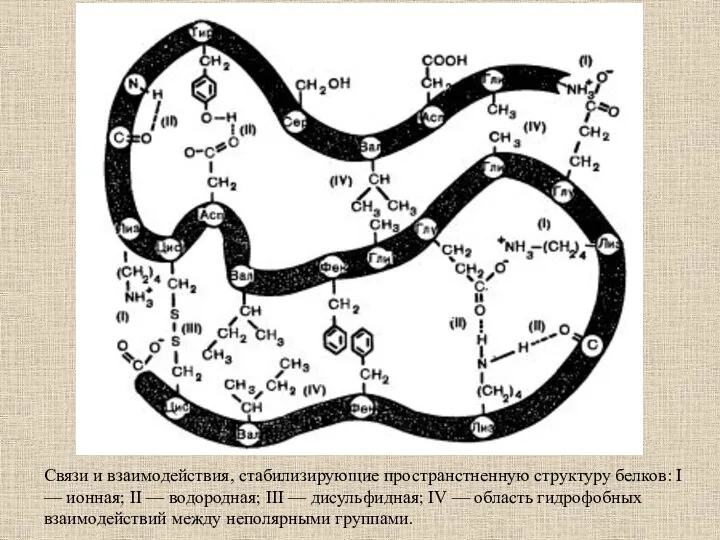
- 13. Связи и взаимодействия, стабилизирующие пространстненную структуру белков: I — ионная; II — водородная; III — дисульфидная;
- 27. Раздел: Молекулярная биофизика Тема: Динамика биополимеров
- 28. Пространственная структура миоглобина (кашалота) в проекции ху Миоглобин (переносчик кислорода в мышцах) содержит один гем и
- 29. Расщепление d-орбиталей в октаэдрическом комплексе (I): и распределение d-электронов по орбиталям высокоспинового (расщепление Δ мало) и
- 30. Структурные изменения, происходящие в гемоглобине при оксигенации (объяснение см. в тексте) (по Д. Мецлеру, 1980)
- 31. Переход из T в R форму у гемоглобина
- 33. Туннельный перенос электрона в биоструктурах
- 34. Методы изучения подвижности белков Люминесцентные методы ЭПР ЯМР ЯГР спектроскопия Метод изотопного обмена
- 35. Кривые потенциальной энергии основного (So) и синглетного возбужденного (S*) состояний двухатомной молекулы: U — потенциальная энергия;
- 36. Электронные уровни органической молекулы и переходы между ними (схема Яблонского): р — вероятность переходов в единицу
- 37. Люминесцентные методы Измерение внутримолекулярной подвижности белка по зависимости положения максимума люминесценции метки, введенной в белок, либо
- 38. Зависимость положения спектра флуорисценции водного раствора (3-лактоглобулина (I) и нейтротоксина II кобры (II) от температуры при
- 39. Методы радиоспектроскопии ЭПР и ЯМР Расщепление энергетических уровней электрона (протона) в магнитном поле (Н)
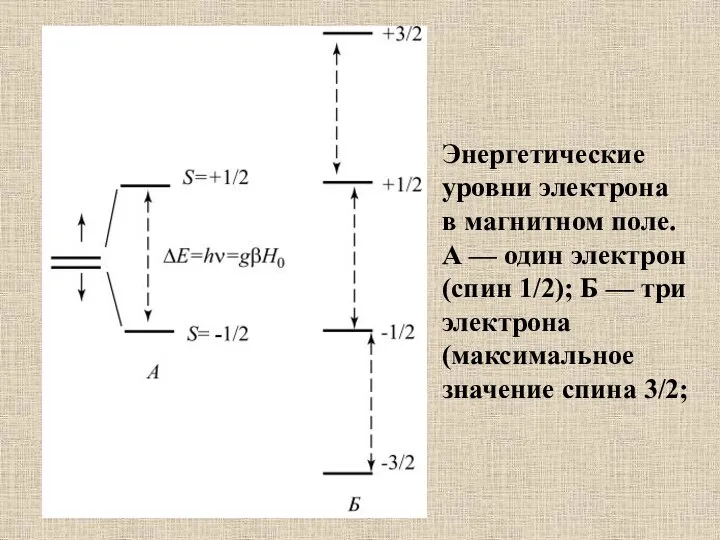
- 40. Энергетические уровни электрона в магнитном поле. А — один электрон (спин 1/2); Б — три электрона
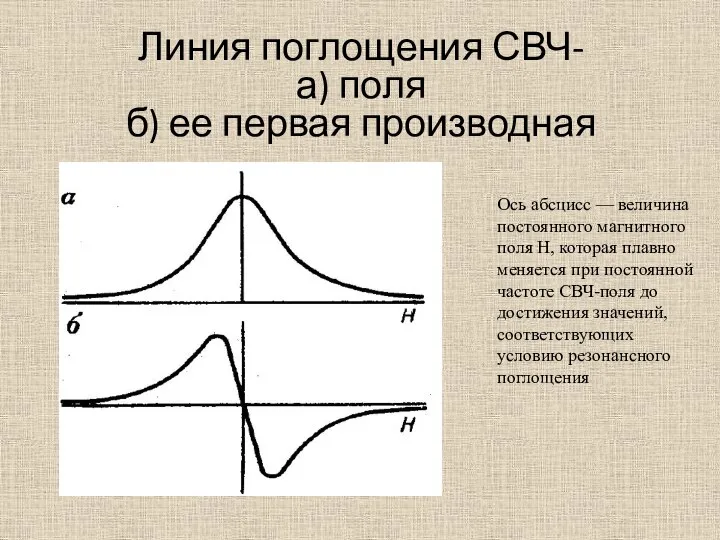
- 41. Линия поглощения СВЧ- а) поля б) ее первая производная Ось абсцисс — величина постоянного магнитного поля
- 42. Линия резонанса ЭПР Ширина: Т1 – время передачи энергии окружающей среде Т2 – время спин-спинового взаимодействия
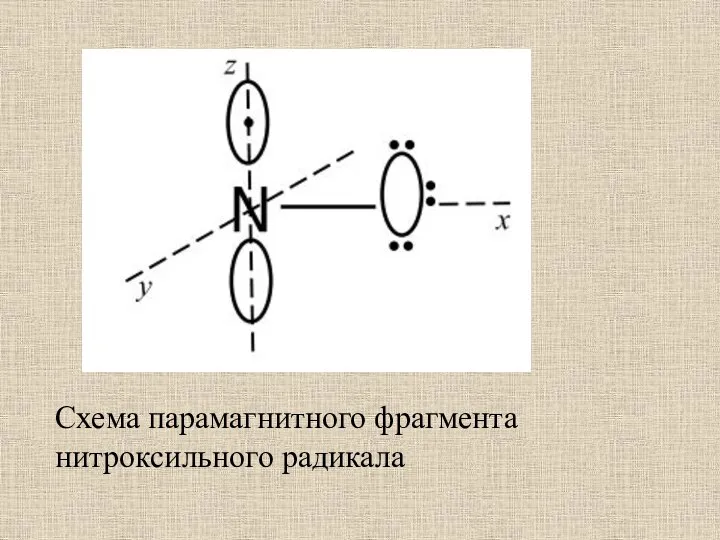
- 43. Схема парамагнитного фрагмента нитроксильного радикала
- 44. Спектр ЭПР парамагнитной метки, присоединенной к гис-15 лизоцима (рН 7,0; t=26°C) (по Г.И. Лихтенштейну, 1971): h-1,
- 45. ЯМР-спектроскопия Измерение времени релаксации Т1 и Т2 по ширине линии резонанса. Определение времени вращения метки, на
- 46. Спектр ЯМР ацетальдегида СН3СНО СНО химический сдвиг СН3 по А.Керрингтону, Э. Мак-Лечлану, 1970
- 47. ЯГР спектроскопия Дает информацию не только о временных, а также амплитудных характеристиках движений в белке (средние
- 48. Эффект Мёссбауэра Уширение спектра обусловлено диффузией молекул белка. Изменение частоты уширения пропорционально скорости источника (v)
- 51. Ферментативный катализ
- 52. Ферменты (от латинского fermentum – закваска), энзимы (от греческого en – "в" и zyme – "закваска")
- 53. Ферменты отличаются от других катализаторов тремя уникальными свойствами: - высокой эффективностью действия, - специфичностью действия, -
- 59. Взаимодействия, которые играют главную роль при связывании субстрата в активном центре фермента и образовании комплекса фермент-субстрат
- 61. Основные механизмы управления каталитической активностью фермента связаны со следующими явлениями и процессами: 1) аллостерия; 2) кооперативность;
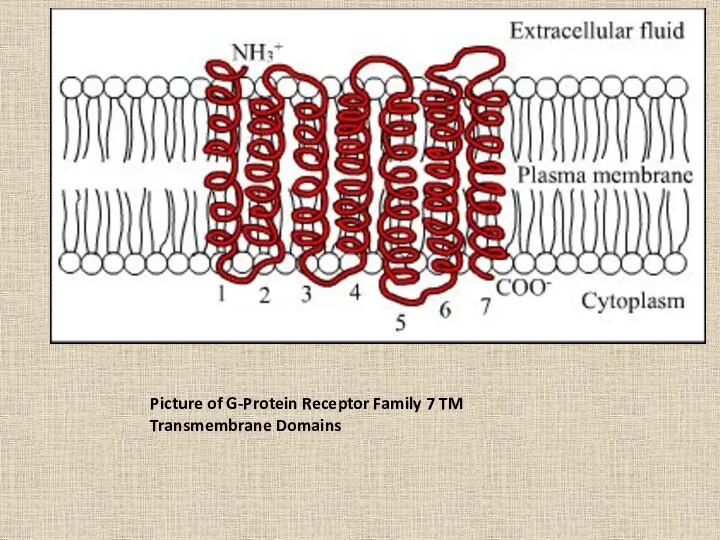
- 65. Picture of G-Protein Receptor Family 7 TM Transmembrane Domains
- 71. Скачать презентацию
Слайд 2Конформационная энергия полипептида
Общее строение полипептидной цепи,
характеризуется тремя углами:
Конформационная энергия полипептида
Общее строение полипептидной цепи,
характеризуется тремя углами:
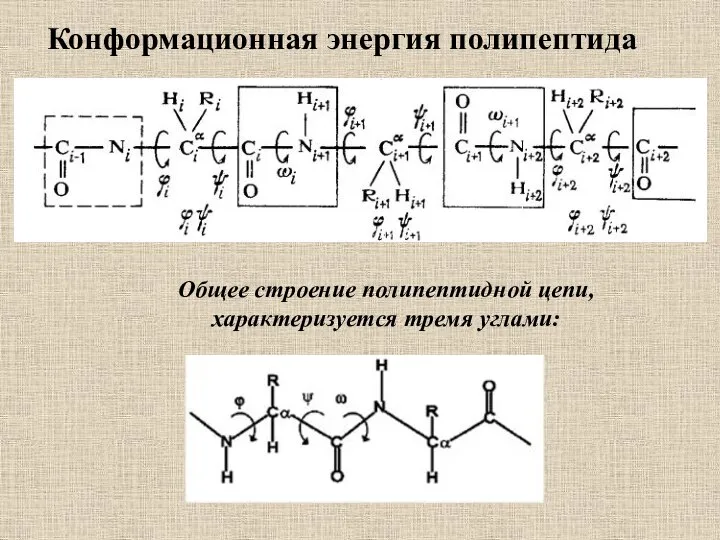
Слайд 3Пептидная связь – ω - угол
Однако, двойной характер пептидной связи препятствует вращению
Пептидная связь – ω - угол
Однако, двойной характер пептидной связи препятствует вращению
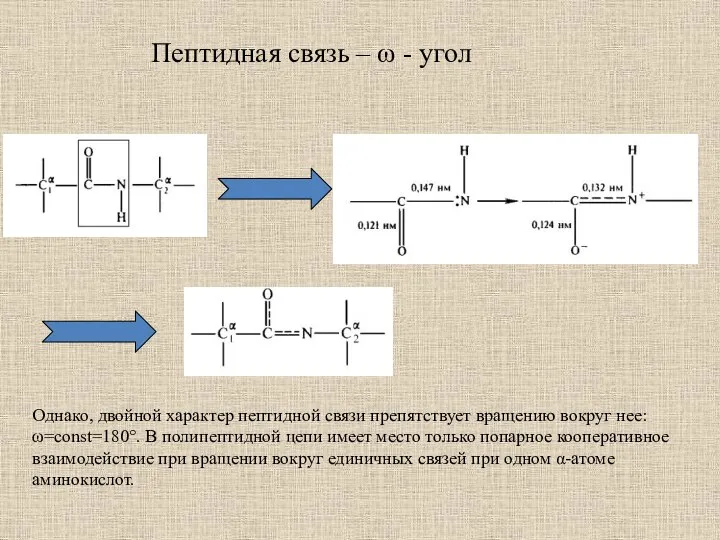
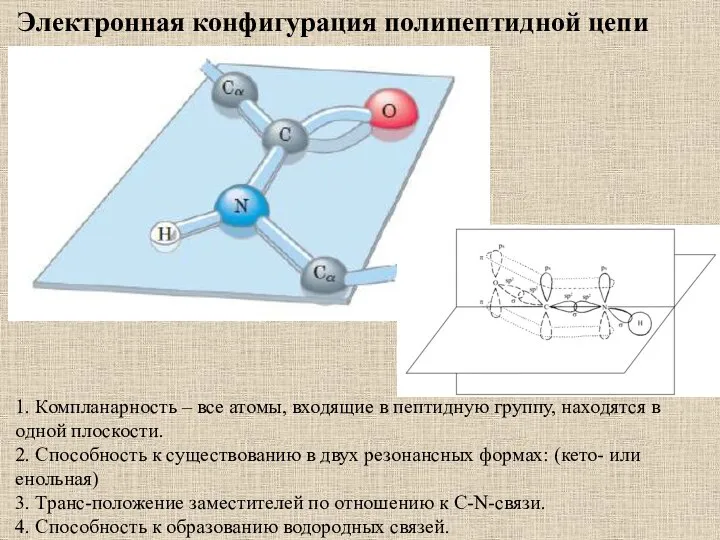
Слайд 41. Компланарность – все атомы, входящие в пептидную группу, находятся в одной
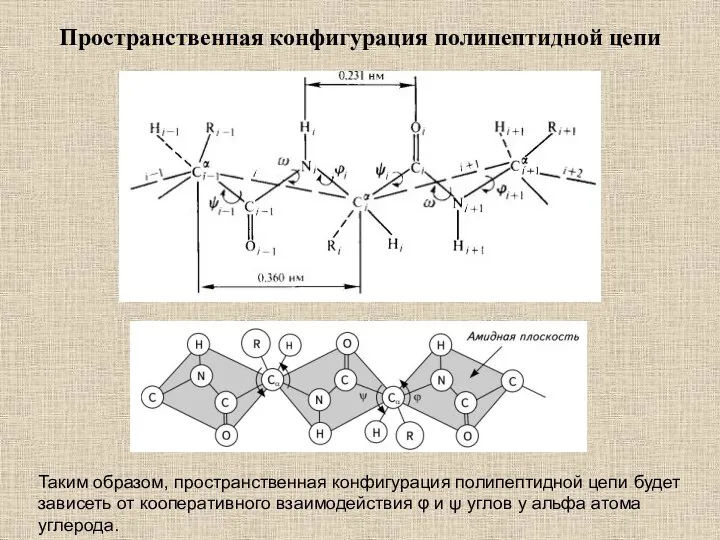
Слайд 5Пространственная конфигурация полипептидной цепи
Таким образом, пространственная конфигурация полипептидной цепи будет зависеть от
Пространственная конфигурация полипептидной цепи
Таким образом, пространственная конфигурация полипептидной цепи будет зависеть от
Слайд 6Не все значения углов ψ и φ допустимы вследствие возможного стерического несоответствия
Не все значения углов ψ и φ допустимы вследствие возможного стерического несоответствия




Слайд 12Вторичная структура полипептида
Α-спираль
β – формы
Вторичная структура полипептида
Α-спираль
β – формы
Слайд 13Связи и взаимодействия, стабилизирующие пространстненную структуру белков: I — ионная; II —
Связи и взаимодействия, стабилизирующие пространстненную структуру белков: I — ионная; II —
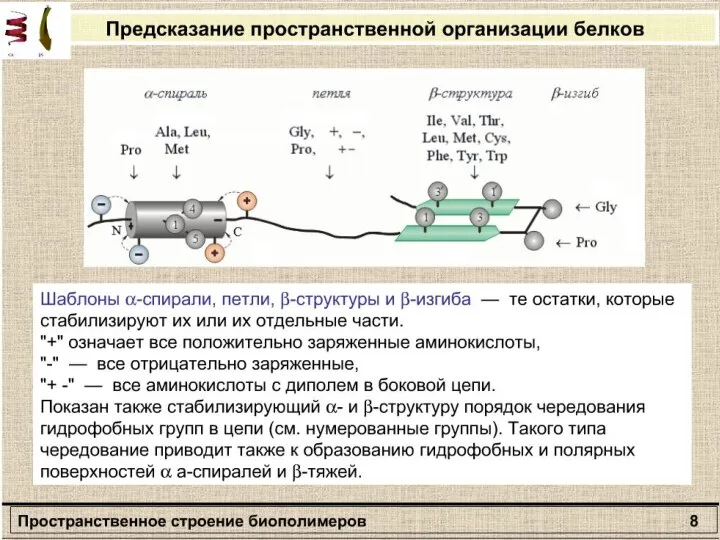

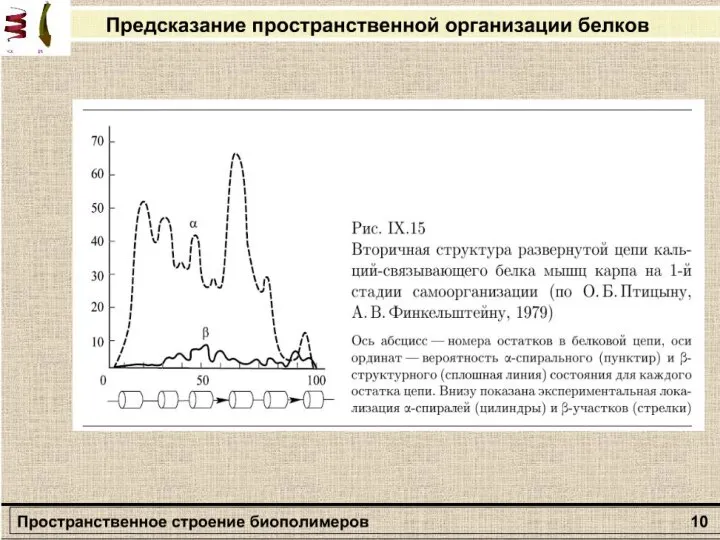
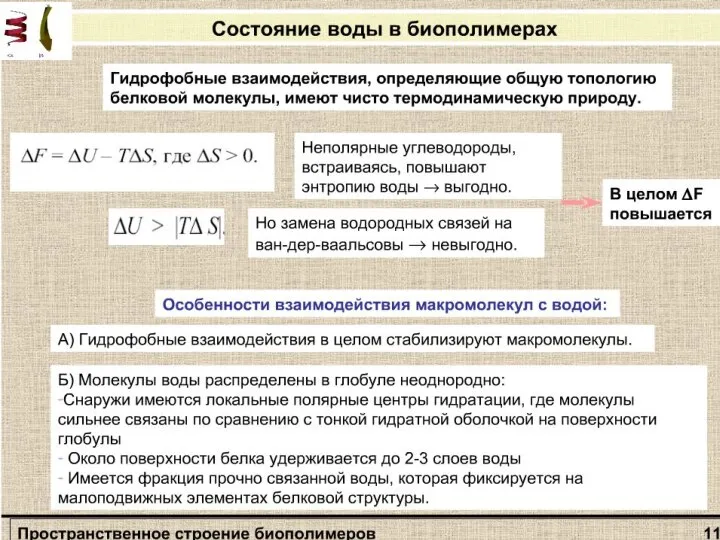
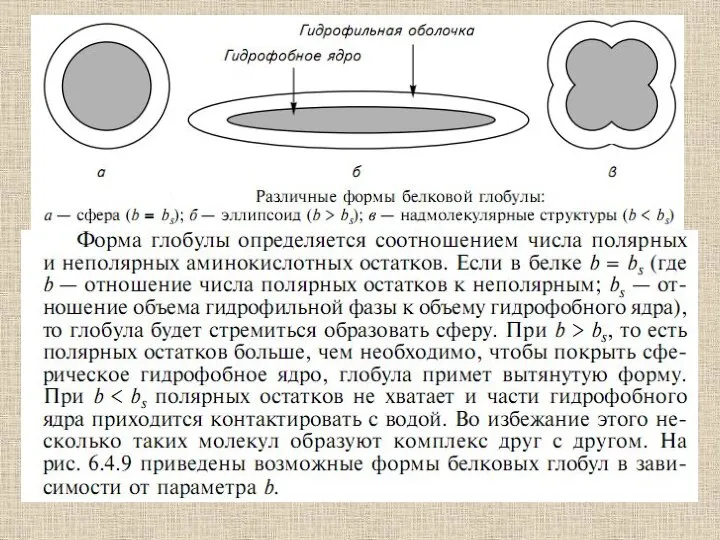

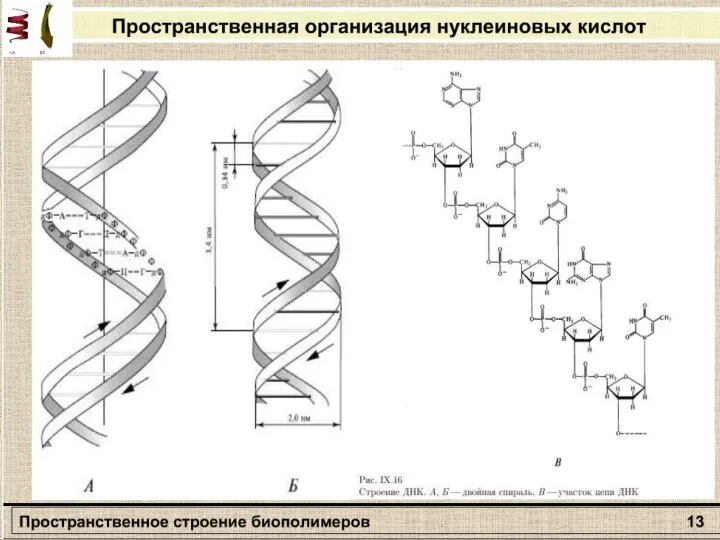
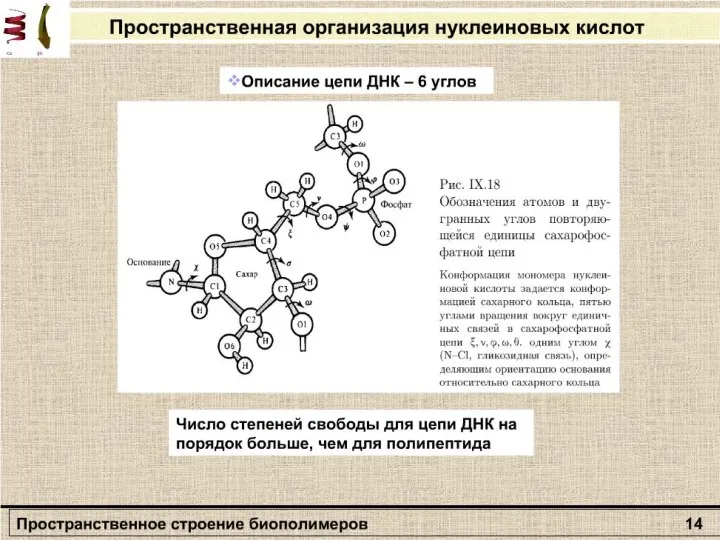


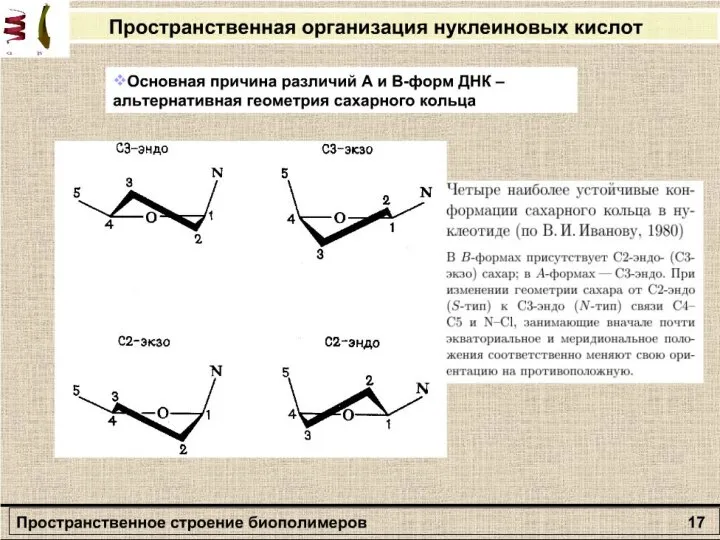
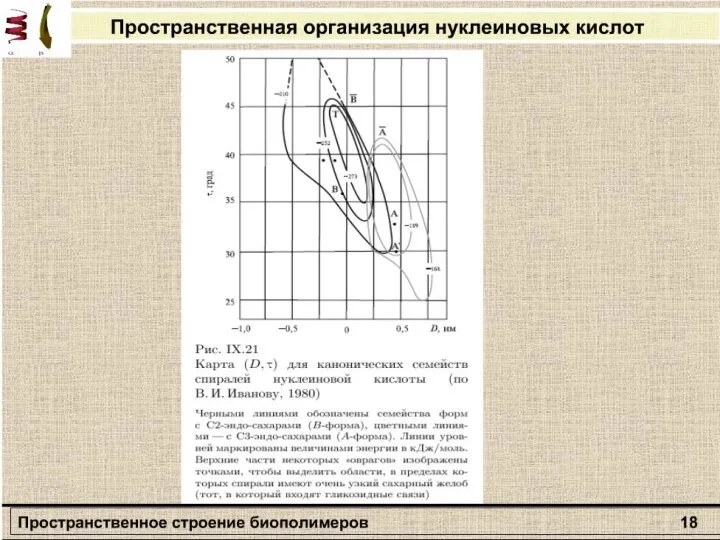

Слайд 27Раздел: Молекулярная биофизика
Тема: Динамика биополимеров
Раздел: Молекулярная биофизика
Тема: Динамика биополимеров

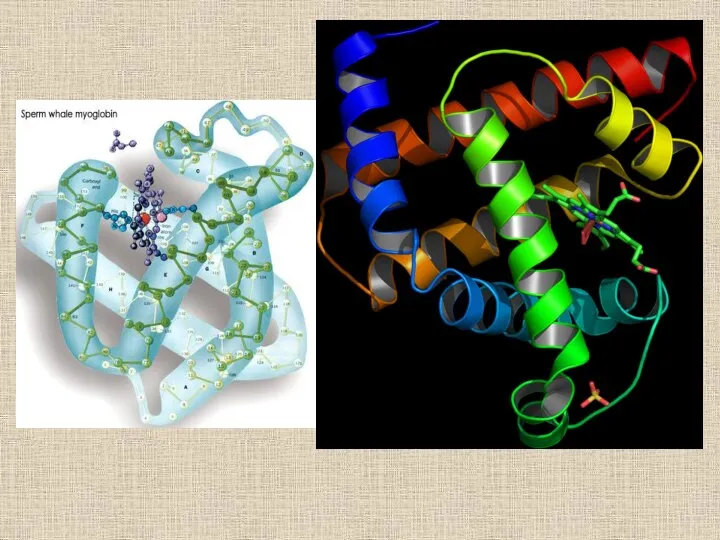
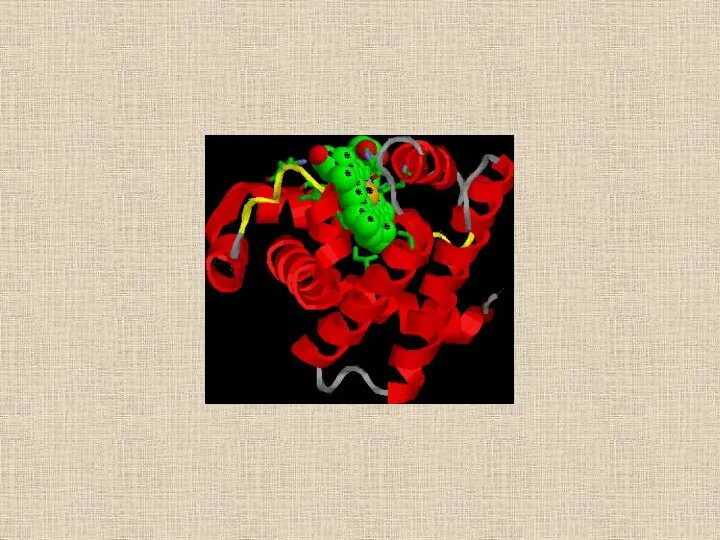
Слайд 28Пространственная структура миоглобина (кашалота) в проекции ху
Миоглобин (переносчик кислорода в мышцах) содержит
Пространственная структура миоглобина (кашалота) в проекции ху
Миоглобин (переносчик кислорода в мышцах) содержит

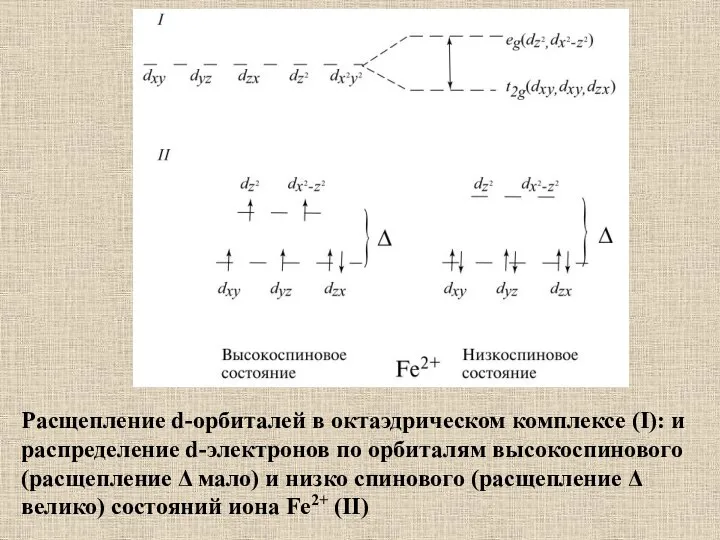
Слайд 29Расщепление d-орбиталей в октаэдрическом комплексе (I): и распределение d-электронов по орбиталям высокоспинового
Расщепление d-орбиталей в октаэдрическом комплексе (I): и распределение d-электронов по орбиталям высокоспинового
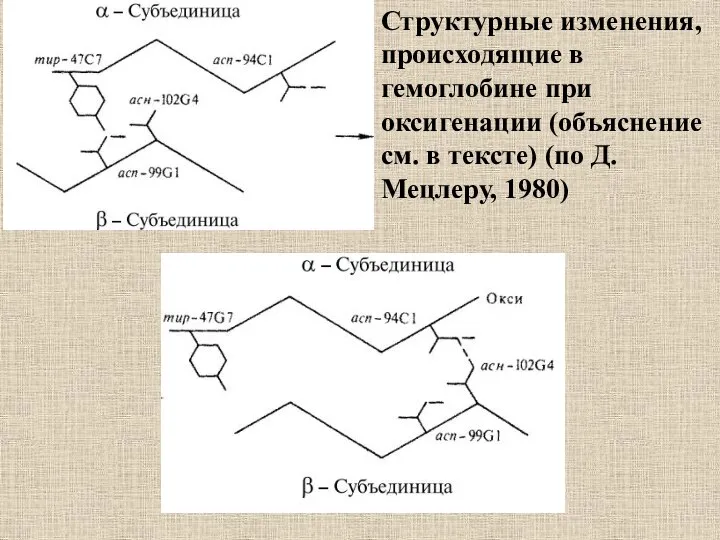
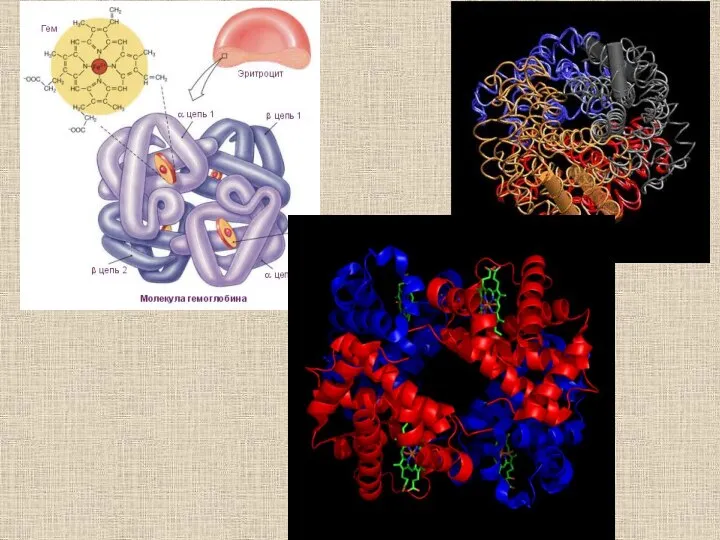
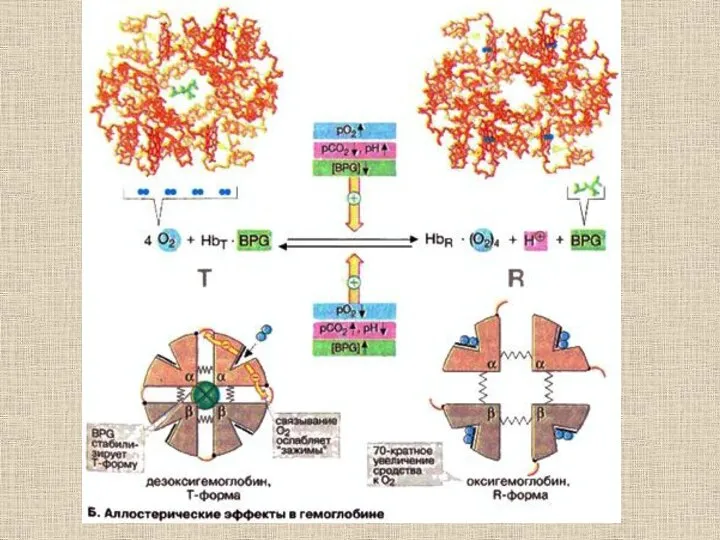
Слайд 30Структурные изменения, происходящие в гемоглобине при оксигенации (объяснение см. в тексте) (по
Структурные изменения, происходящие в гемоглобине при оксигенации (объяснение см. в тексте) (по
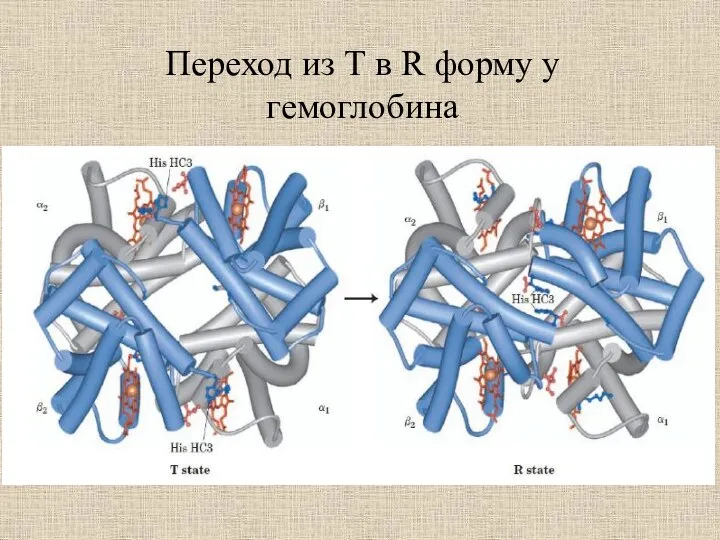
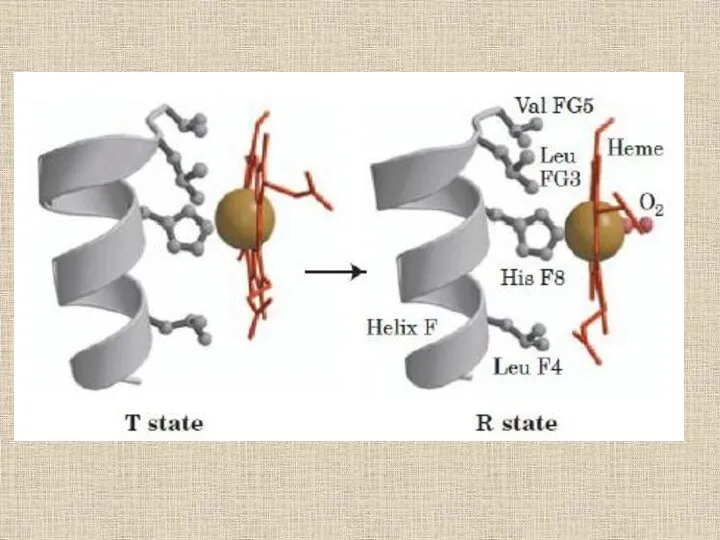
Слайд 31Переход из T в R форму у гемоглобина
Переход из T в R форму у гемоглобина
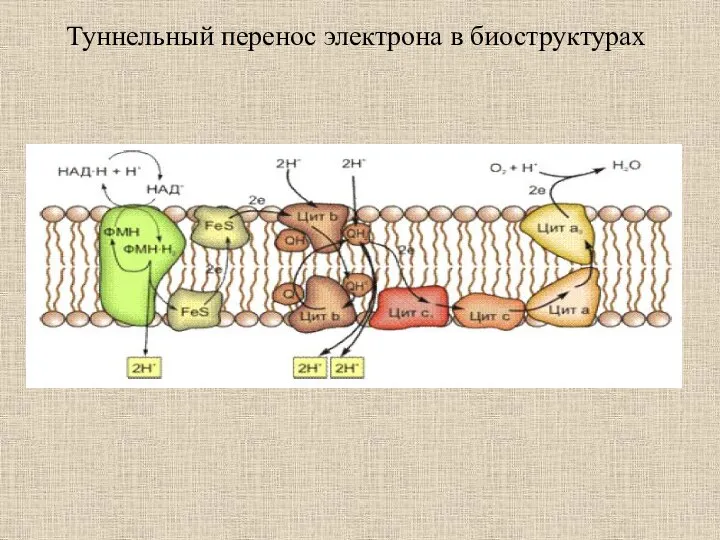
Слайд 33Туннельный перенос электрона в биоструктурах
Туннельный перенос электрона в биоструктурах
Слайд 34Методы изучения подвижности белков
Люминесцентные методы
ЭПР
ЯМР
ЯГР спектроскопия
Метод изотопного обмена
Методы изучения подвижности белков
Люминесцентные методы
ЭПР
ЯМР
ЯГР спектроскопия
Метод изотопного обмена

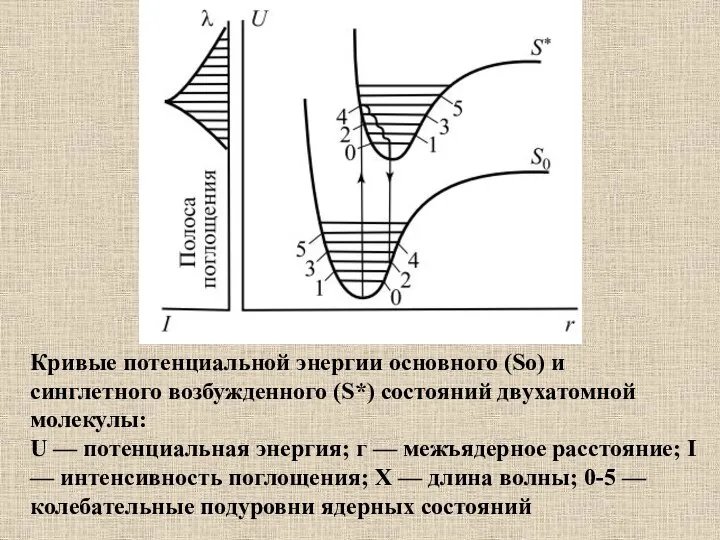
Слайд 35Кривые потенциальной энергии основного (So) и синглетного возбужденного (S*) состояний двухатомной молекулы:
U
Кривые потенциальной энергии основного (So) и синглетного возбужденного (S*) состояний двухатомной молекулы:
U
Слайд 36Электронные уровни органической молекулы и переходы между ними (схема Яблонского): р —
Электронные уровни органической молекулы и переходы между ними (схема Яблонского): р —

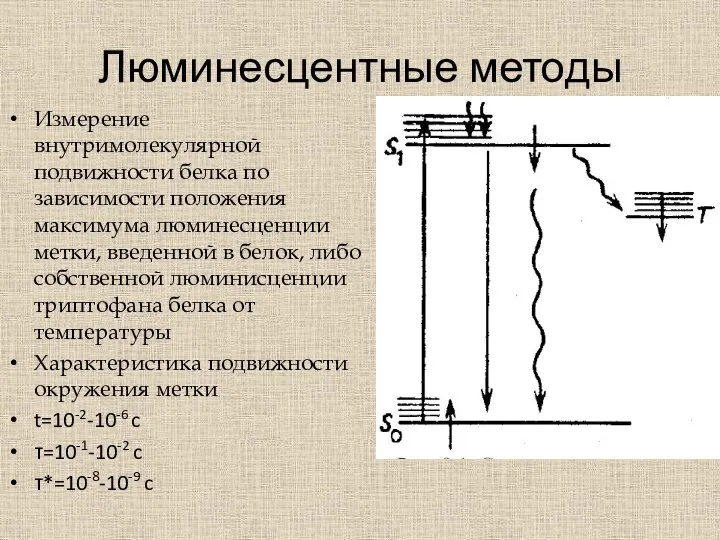
Слайд 37Люминесцентные методы
Измерение внутримолекулярной подвижности белка по зависимости положения максимума люминесценции метки, введенной
Люминесцентные методы
Измерение внутримолекулярной подвижности белка по зависимости положения максимума люминесценции метки, введенной
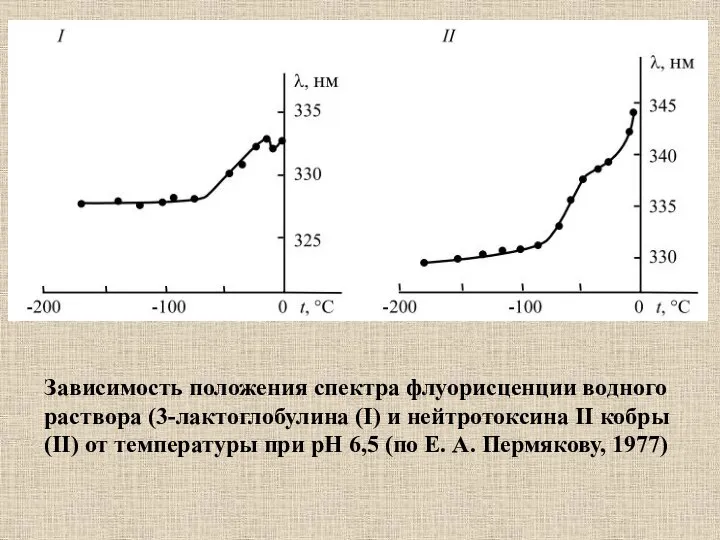
Слайд 38Зависимость положения спектра флуорисценции водного раствора (3-лактоглобулина (I) и нейтротоксина II кобры
Зависимость положения спектра флуорисценции водного раствора (3-лактоглобулина (I) и нейтротоксина II кобры
Слайд 39Методы радиоспектроскопии
ЭПР и ЯМР
Расщепление энергетических уровней электрона (протона) в магнитном поле (Н)
Методы радиоспектроскопии
ЭПР и ЯМР
Расщепление энергетических уровней электрона (протона) в магнитном поле (Н)
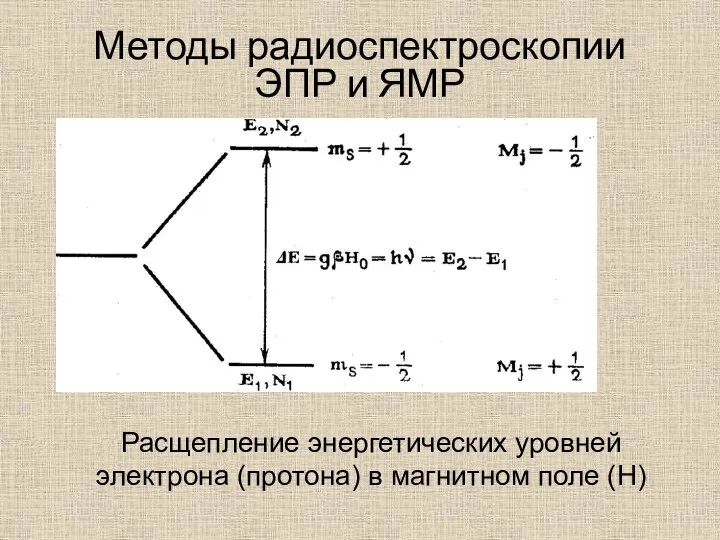
Слайд 40Энергетические уровни электрона в магнитном поле. А — один электрон (спин 1/2);
Энергетические уровни электрона в магнитном поле. А — один электрон (спин 1/2);
Слайд 41Линия поглощения СВЧ-
а) поля
б) ее первая производная
Ось абсцисс — величина постоянного
Линия поглощения СВЧ-
а) поля
б) ее первая производная
Ось абсцисс — величина постоянного
Слайд 42Линия резонанса ЭПР
Ширина:
Т1 – время передачи энергии окружающей среде
Т2 – время
Линия резонанса ЭПР
Ширина:
Т1 – время передачи энергии окружающей среде
Т2 – время

Слайд 43Схема парамагнитного фрагмента нитроксильного радикала
Схема парамагнитного фрагмента нитроксильного радикала
Слайд 44Спектр ЭПР парамагнитной метки, присоединенной к гис-15 лизоцима (рН 7,0; t=26°C) (по
Спектр ЭПР парамагнитной метки, присоединенной к гис-15 лизоцима (рН 7,0; t=26°C) (по
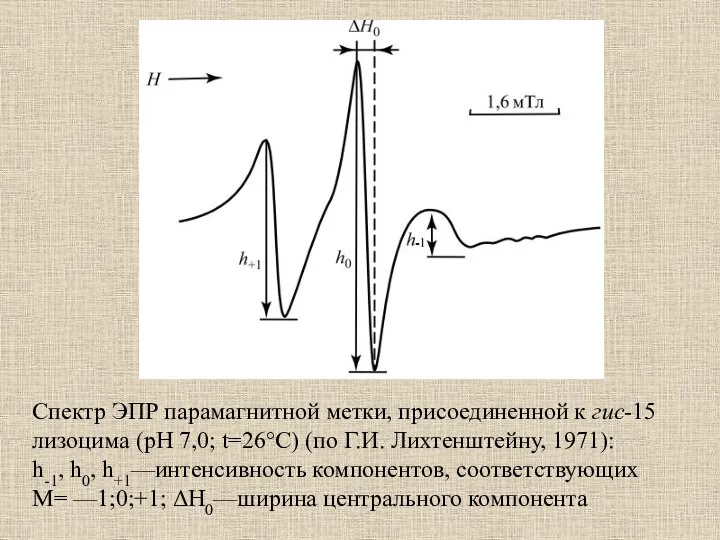
Слайд 45ЯМР-спектроскопия
Измерение времени релаксации Т1 и Т2 по ширине линии резонанса.
Определение времени вращения
ЯМР-спектроскопия
Измерение времени релаксации Т1 и Т2 по ширине линии резонанса.
Определение времени вращения

Слайд 46Спектр ЯМР ацетальдегида СН3СНО
СНО химический сдвиг СН3
по А.Керрингтону, Э. Мак-Лечлану, 1970
Спектр ЯМР ацетальдегида СН3СНО
СНО химический сдвиг СН3
по А.Керрингтону, Э. Мак-Лечлану, 1970
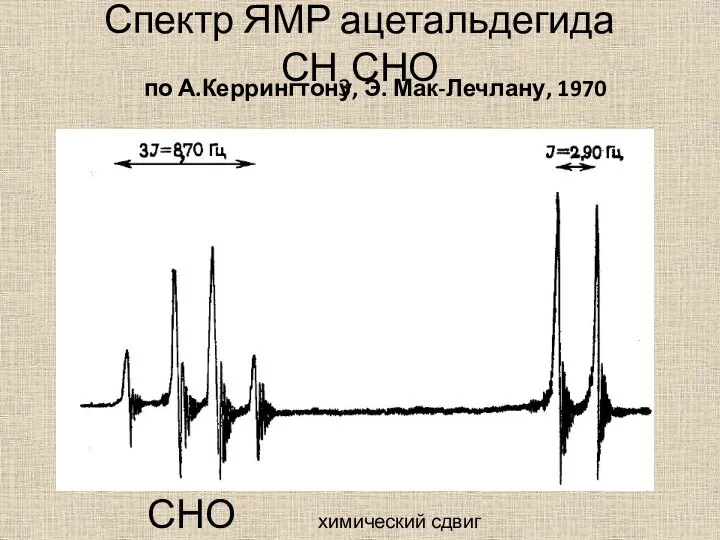
Слайд 47ЯГР спектроскопия
Дает информацию не только о временных, а также амплитудных характеристиках движений
ЯГР спектроскопия
Дает информацию не только о временных, а также амплитудных характеристиках движений

Слайд 48Эффект Мёссбауэра
Уширение спектра обусловлено диффузией молекул белка. Изменение частоты уширения пропорционально
Эффект Мёссбауэра
Уширение спектра обусловлено диффузией молекул белка. Изменение частоты уширения пропорционально


Слайд 51Ферментативный катализ
Ферментативный катализ

Слайд 52Ферменты (от латинского fermentum – закваска), энзимы (от греческого en – "в"
Ферменты (от латинского fermentum – закваска), энзимы (от греческого en – "в"

Слайд 53Ферменты отличаются от других катализаторов тремя уникальными свойствами:
- высокой эффективностью действия,
- специфичностью
Ферменты отличаются от других катализаторов тремя уникальными свойствами:
- высокой эффективностью действия,
- специфичностью

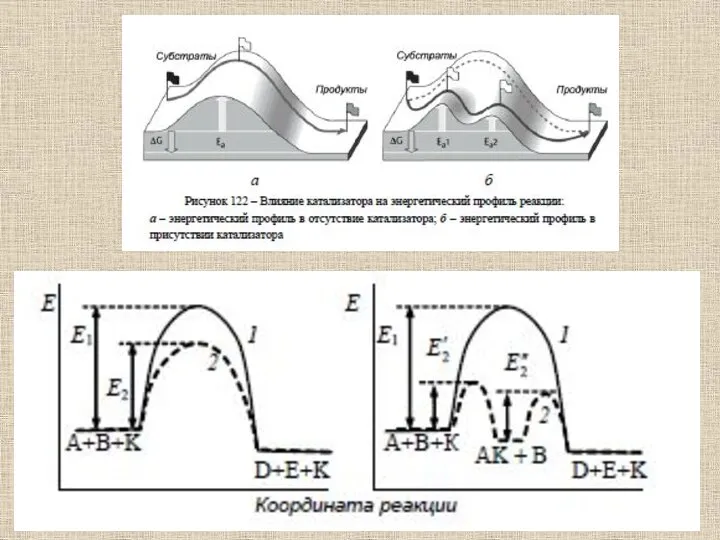
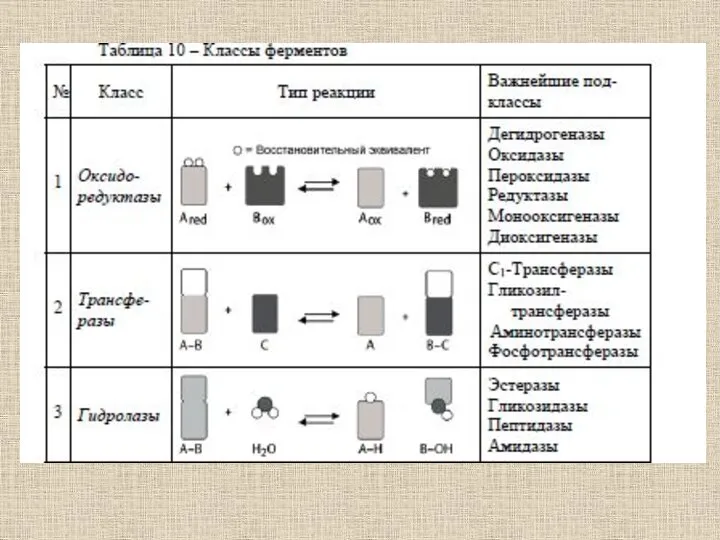
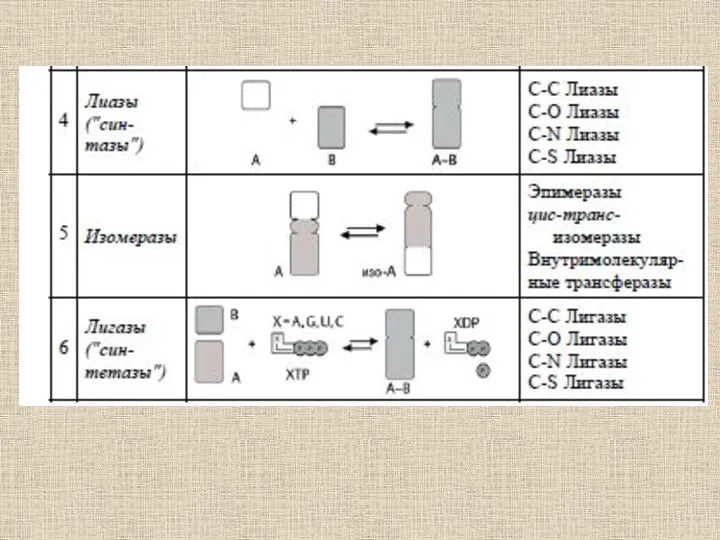
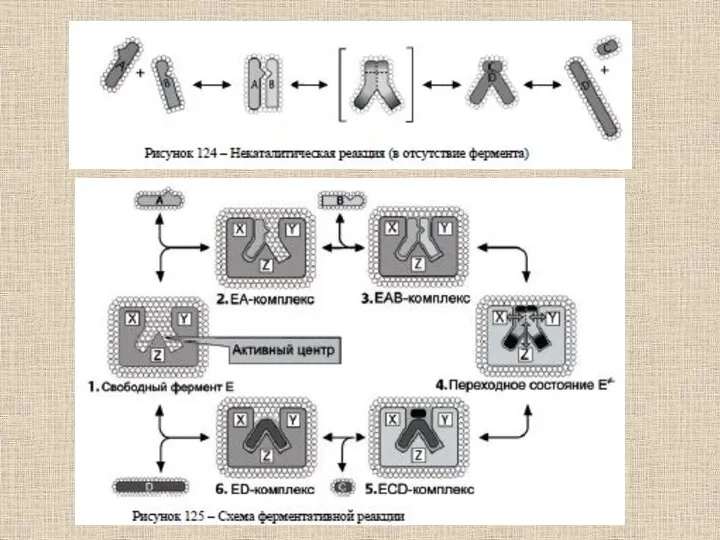
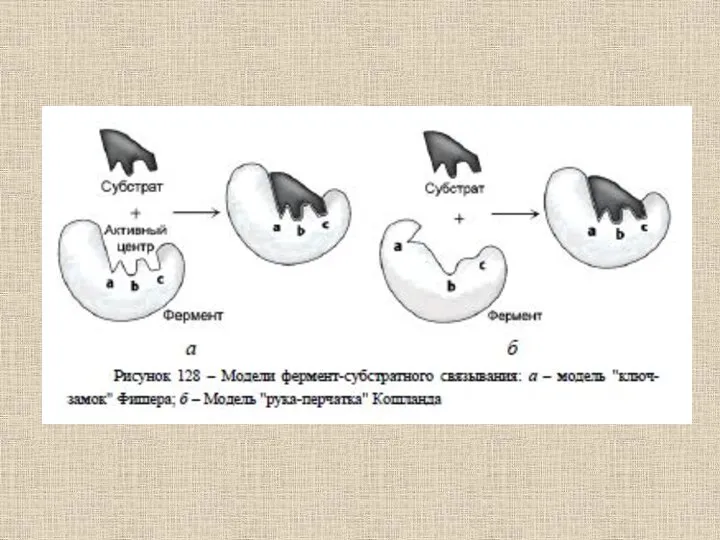
Слайд 59Взаимодействия, которые играют главную роль при связывании субстрата в активном центре фермента
Взаимодействия, которые играют главную роль при связывании субстрата в активном центре фермента

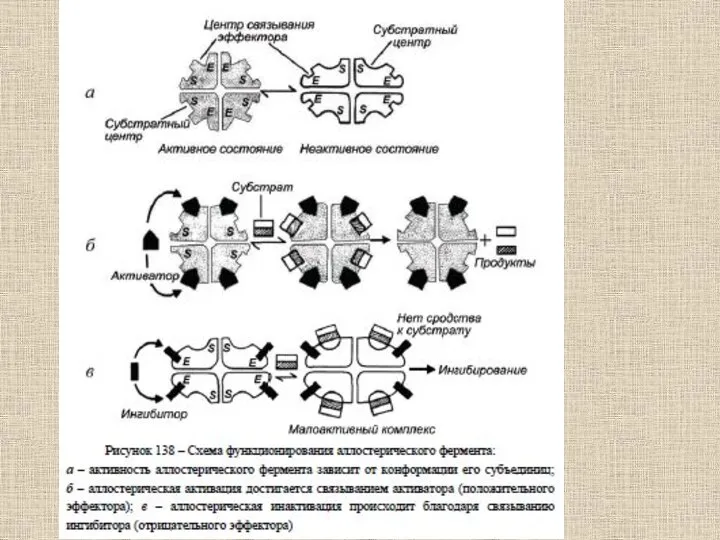
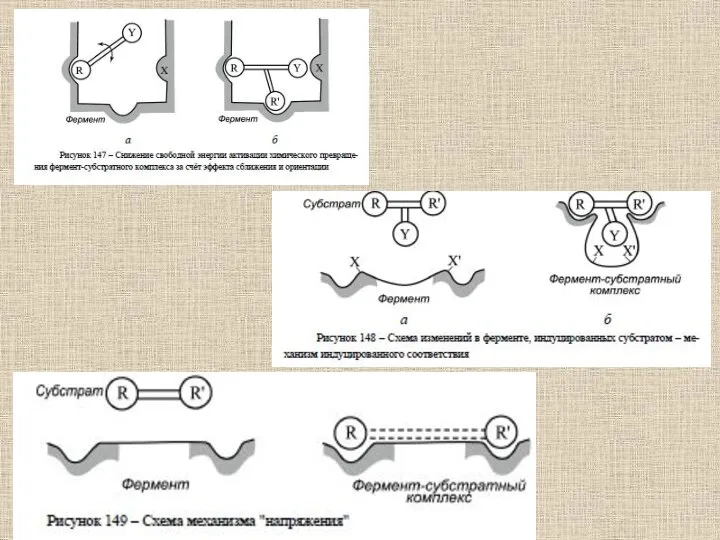
Слайд 61Основные механизмы управления каталитической активностью фермента связаны со следующими явлениями и процессами:
1)
Основные механизмы управления каталитической активностью фермента связаны со следующими явлениями и процессами:
1)

Слайд 65Picture of G-Protein Receptor Family 7 TM Transmembrane Domains
Picture of G-Protein Receptor Family 7 TM Transmembrane Domains

 Витамины в продуктах питания
Витамины в продуктах питания Мята. Лекарственные растения
Мята. Лекарственные растения Основы эндокринологии. Анатомия и физиология человека
Основы эндокринологии. Анатомия и физиология человека Основные признаки животных организмов
Основные признаки животных организмов List - nadzemný orgán rastliny
List - nadzemný orgán rastliny Органические вещества
Органические вещества At the fruit farm
At the fruit farm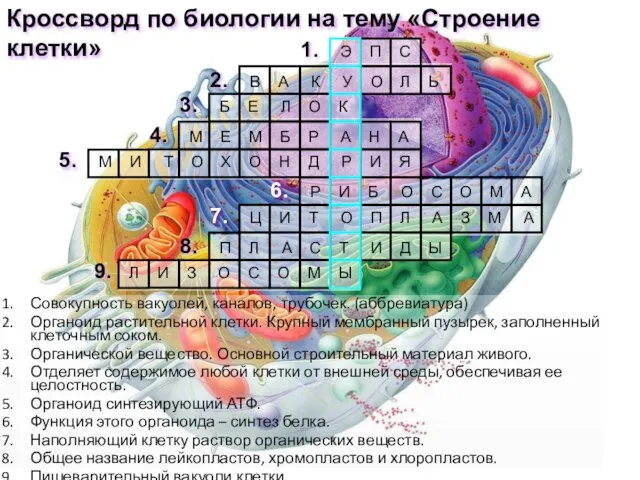 Презентация на тему Кроссворд. Строение клетки
Презентация на тему Кроссворд. Строение клетки  Фотосинтез. Ян ван Гельмонт
Фотосинтез. Ян ван Гельмонт Приручение животных
Приручение животных Выделительные системы позвоночных разных классов
Выделительные системы позвоночных разных классов Царство Грибы. Грибы-паразиты.
Царство Грибы. Грибы-паразиты. Основы гистологии
Основы гистологии Весна
Весна «Биогенная роль щелочных металлов (натрия и калия) в организме человека» Автор: учащающиеся 9 «А» класса МОУ СОШ № 41 Плиев Ал
«Биогенная роль щелочных металлов (натрия и калия) в организме человека» Автор: учащающиеся 9 «А» класса МОУ СОШ № 41 Плиев Ал Комнатные растения
Комнатные растения Функции нижних конечностей
Функции нижних конечностей В гостях у дикой природы. Лоси
В гостях у дикой природы. Лоси Викторина Что вы знаете о лесе?
Викторина Что вы знаете о лесе? Бактерии. 5 класс
Бактерии. 5 класс Типы питания живых организмов. 6 класс
Типы питания живых организмов. 6 класс Презентация на тему Влияние школьного портфеля на осанку
Презентация на тему Влияние школьного портфеля на осанку  История первых великих путешествий
История первых великих путешествий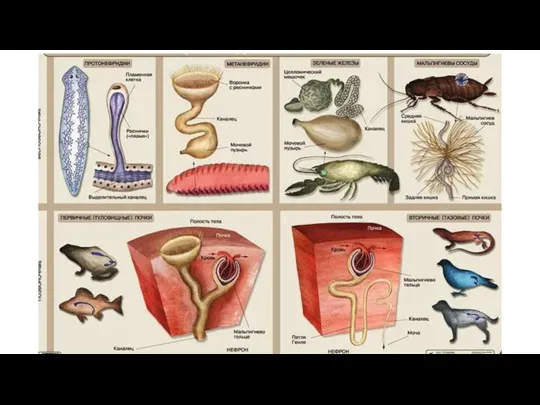 Внешний вид почек позвоночных животных
Внешний вид почек позвоночных животных Филогенез сердечно-сосудистой системы хордовых
Филогенез сердечно-сосудистой системы хордовых Работа с графической информацией на ВПР по биологии. Задание 6, 5 класс
Работа с графической информацией на ВПР по биологии. Задание 6, 5 класс Организм и его уровни организации (лекция 2)
Организм и его уровни организации (лекция 2) Размножение и оплодотворение у растений
Размножение и оплодотворение у растений