- Мышечная и соед-я ткань (1)

Содержание
- 2. Мышечная ткань
- 3. Одним из свойств живой системы является способность к движению. Выделяют три уровня развития двигательной активности: 1)
- 4. Мышечные ткани – это группа возбудимых тканей, различных по происхождению и строению, но имеющих сходную функцию
- 5. Общие функции мышечных тканей 1) сократительная 2) защитная 3) формообразующая 4) терморегуляция 5) трофическая
- 6. Общая морфофункциональная характеристика мышечных тканей структурные элементы (клетки, волокна) обладают удлиненной формой, наличие органелл специального назначения
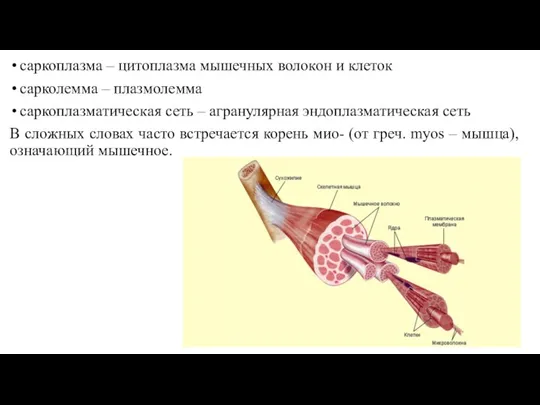
- 9. саркоплазма – цитоплазма мышечных волокон и клеток сарколемма – плазмолемма саркоплазматическая сеть – агранулярная эндоплазматическая сеть
- 10. Классификация мышечных тканей Классификация мышечных тканей основана: на признаках их строения и функции (морфофункциональная классификация) происхождения
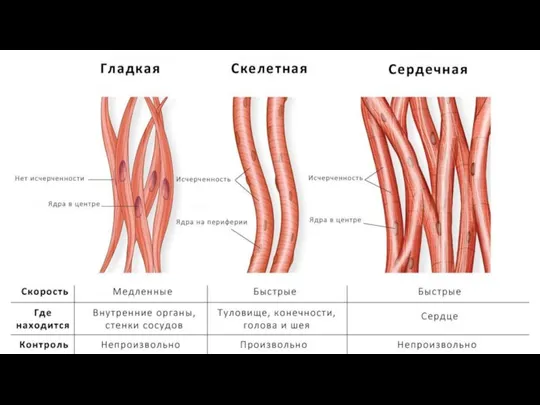
- 11. Морфофункциональная классификация мышечных тканей 1. Гладкие мышечные ткани – состоят из клеток, не имеющих поперечной исчерченности.
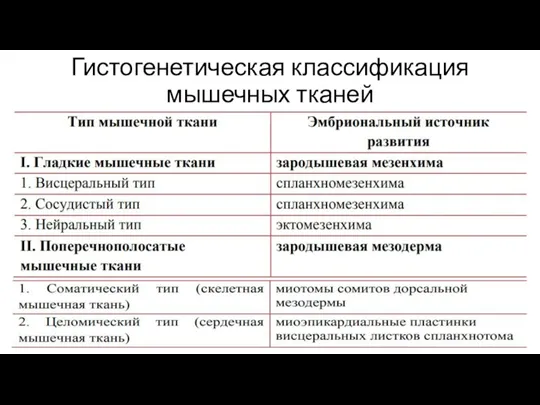
- 13. Гистогенетическая классификация мышечных тканей
- 14. Гладкая мышечная ткань
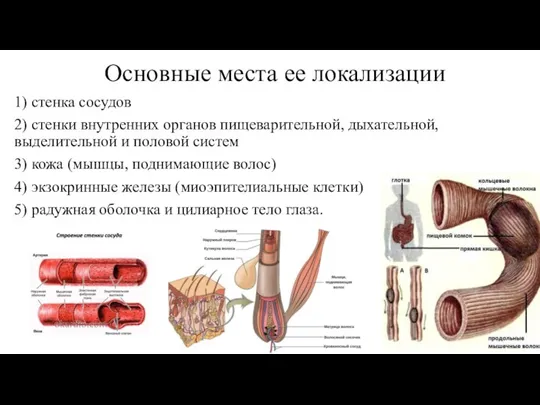
- 15. Основные места ее локализации 1) стенка сосудов 2) стенки внутренних органов пищеварительной, дыхательной, выделительной и половой
- 16. Гистогенез гладкой мышечной ткани 1 стадия – закладки – размножение стволовых клеток (СК), образовавшихся из спланхномезенхимы;
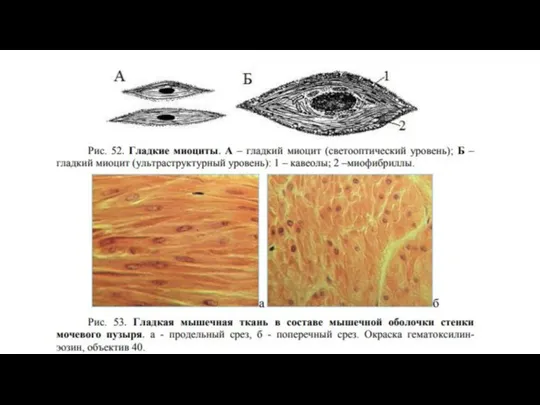
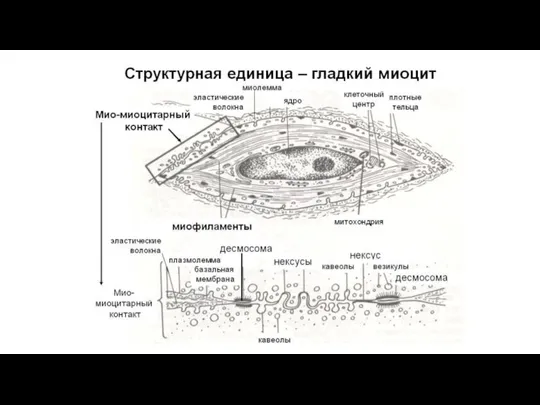
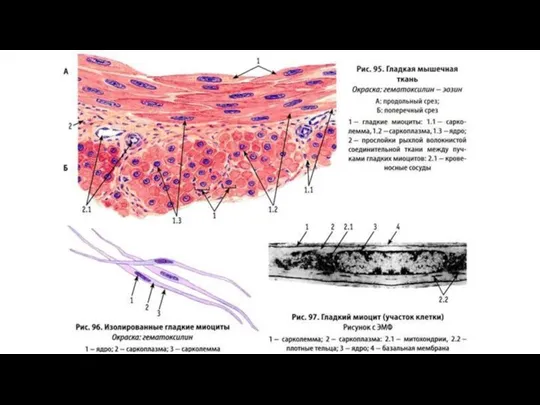
- 17. Строение гладкой мышечной ткани Структурно-функциональной единицей гладкой мышечной ткани является гладкий миоцит (ГМ). Это одноядерные клетки,
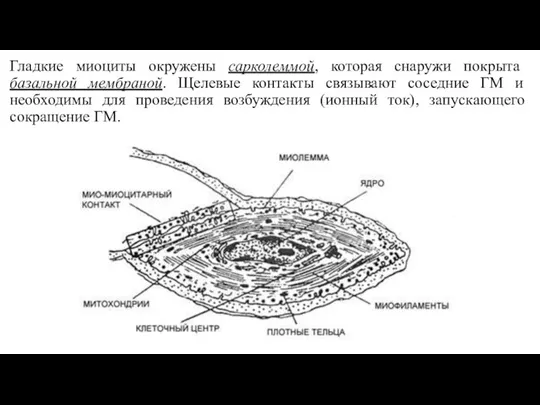
- 19. Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной. Щелевые контакты связывают соседние ГМ и необходимы
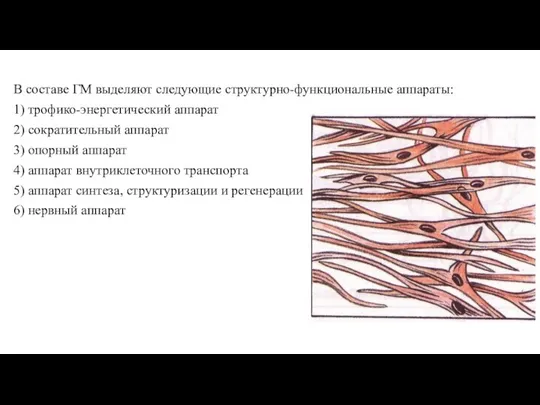
- 21. В составе ГМ выделяют следующие структурно-функциональные аппараты: 1) трофико-энергетический аппарат 2) сократительный аппарат 3) опорный аппарат
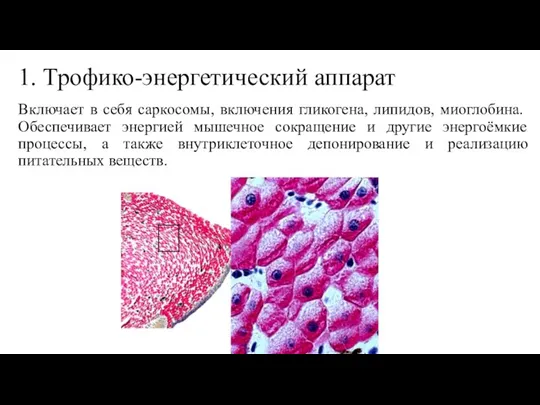
- 22. 1. Трофико-энергетический аппарат Включает в себя саркосомы, включения гликогена, липидов, миоглобина. Обеспечивает энергией мышечное сокращение и
- 23. 2. Сократительный аппарат Представлен тонкими (актиновыми) и толстыми (миозиновыми) филаментами, которые не образуют миофибрилл, в отличие
- 24. Сокращение гладких миоцитов индуцируется притоком в саркоплазму ионов кальция и происходит в соответствии с моделью скользящих
- 25. 3. Опорный аппарат Включает: Сарколемму Цитоскелет промежуточные филаменты плотные тельца межклеточные контакты (десмосомы, нексусы). Обеспечивает: Формообразовательную
- 26. 4. Аппарат внутриклеточного транспорта К нему относятся: кавеолы – колбовидные впячивания сарколеммы диаметром 50-90 нм, расположенные
- 27. 5. Аппарат синтеза, структуризации и регенерации Включает: свободные рибосомы ЭПС комплекс Гольджи лизосомы. Обеспечивает процессы внутриклеточной
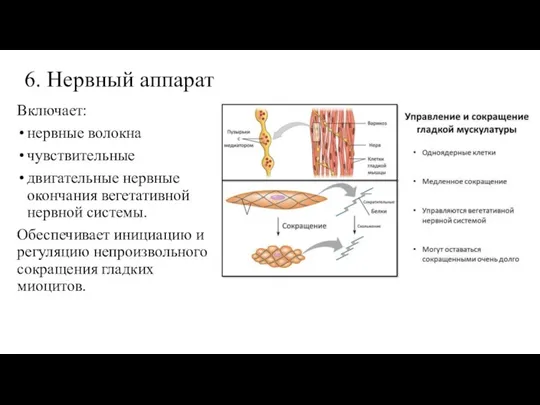
- 28. 6. Нервный аппарат Включает: нервные волокна чувствительные двигательные нервные окончания вегетативной нервной системы. Обеспечивает инициацию и
- 30. Регенерация гладкой мышечной ткани А. Механизмы 1. Эндорепродукция – внутриклеточное образование, обновление и восстановление структур (в
- 31. Б. Виды 1. Физиологическая регенерация (постэмбриональный миогенез) • Протекает постоянно. • Усиливается при повышении физиологической нагрузки
- 32. Гладкая мышечная ткань в составе органов В органах гладкая мышечная ткань преимущественно представлена: пластами слоями пучками.
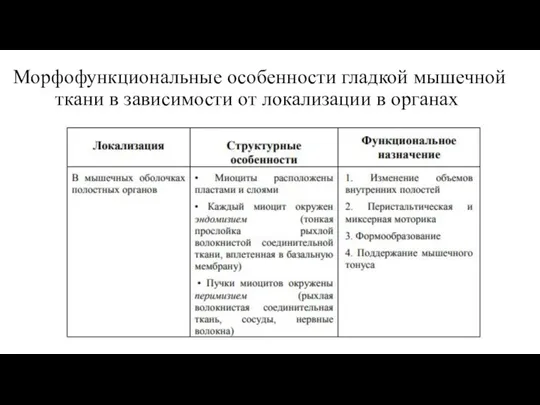
- 33. Морфофункциональные особенности гладкой мышечной ткани в зависимости от локализации в органах
- 35. Поперечнополосатые мышечные ткани
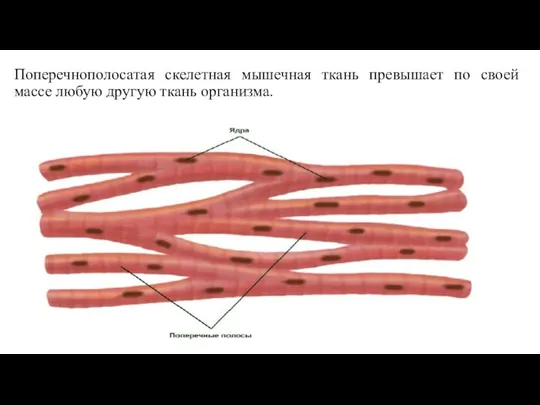
- 36. Поперечнополосатая скелетная мышечная ткань превышает по своей массе любую другую ткань организма.
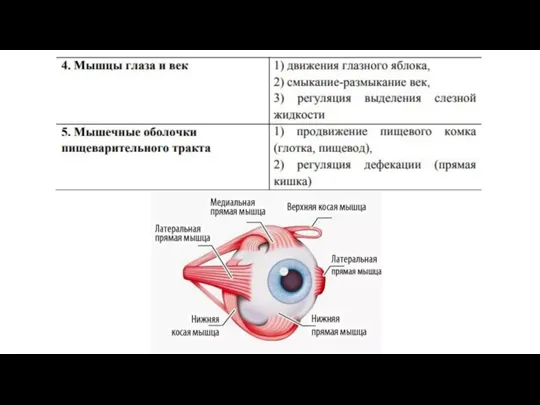
- 37. Локализация и функциональное назначение скелетной мышечной ткани
- 39. Гистогенез поперечнополосатой скелетной мышечной ткани Источник развития поперечнополосатой скелетной мышечной ткани – клетки миотомов сомитов дорсальной
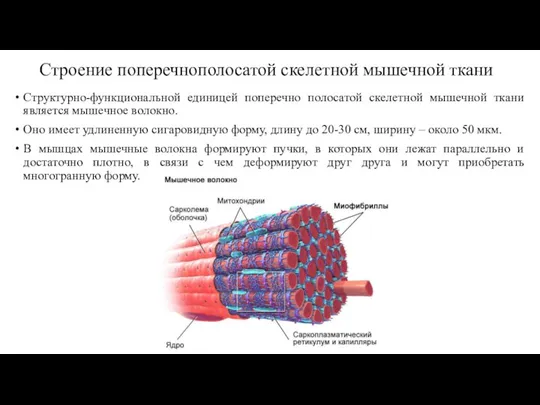
- 41. Строение поперечнополосатой скелетной мышечной ткани Структурно-функциональной единицей поперечно полосатой скелетной мышечной ткани является мышечное волокно. Оно
- 43. На светооптическом уровне после окраски в мышечном волокне наблюдается чередование участков разной интенсивности – темных и
- 44. Компоненты мышечного волокна: 1) миосимпласт – занимает основной объем и ограничен сарколеммой; 2) миосателлитоциты – мелкие,
- 46. Структурно-функциональные аппараты мышечного волокна: 1) трофико-энергетический 2) аппарат синтеза, структуризации и регенерации 3) сократительный аппарат 4)
- 47. Трофико-энергетический аппарат представлен митохондриями и трофическими включениями. Митохондрии вырабатывают энергию, необходимую для сокращения, а также для
- 48. Аппарат синтеза, структуризации и регенерации Основными элементами волокна, обеспечивающими процессы синтеза и структуризации, являются свободные рибосомы,
- 49. Сократительный аппарат представлен специальными органеллами – миофибриллами, которые располагаются продольно в центральной части миосимпласта и отделяются
- 50. Миофибриллы имеют вид нитей диаметром 1-2 мкм. Они обладают собственной поперечной исчерченностью и располагаются в мышечном
- 51. Транспортный аппарат Обеспечивает депонирование и выделение ионов кальция, представлен следующими элементами: 1) поперечные мембранные канальцы (Т-система),
- 52. Опорный аппарат. В его состав входят базальная мембрана, сарколемма, цитоскелет, телофрагмы, мезофрагмы, краевые зоны мышечного волокна.
- 53. Нервный аппарат обеспечивает инициацию и регуляцию произвольного сокращения мышечных волокон. Эфферентные нервные волокна (аксоны α-мотонейронов) образуют
- 54. Типы мышечных волокон В зависимости от количества миоглобина и митохондрий выделяют три типа мышечных волокон.
- 55. Красные (первый тип) малый диаметр волокна много миосателлитоцитов тонкие миофибриллы много миоглобина, мало гликогена много митохондрий
- 56. Белые (второй тип) большой диаметр волокна мало миосателлитоцитов толстые миофибриллы мало миоглобина, много гликогена мало митохондрий
- 57. Строение скелетной мышцы как органа Мышцы человека содержат все типы мышечных волокон. Каждое мышечное волокно окружено
- 59. Регенерация скелетной поперечнополосатой мышечной ткани А. Механизмы 1. Эндорепродукция – внутрисимпластическое образование, обновление и восстановление структур
- 60. Б. Виды Физиологическая регенерация (постэмбриональный миогенез) • Протекает постоянно, состоит в самообновлении органелл и других структурных
- 61. Сердечная мышечная ткань
- 62. Сердечная мышечная ткань локализуется в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов.
- 63. Функциональные особенности сердечной мышечной ткани: 1) автоматизм 2) ритмичность 3) непроизвольность 4) малая утомляемость На функциональную
- 64. Гистогенез сердечной мышечной ткани Источником развития сердечной мышечной ткани является миоэпикардиальная пластинка висцерального листка спланхнотома. В
- 65. Строение сердечной мышечной ткани Как уже отмечалось, сердечная мышечная ткань образована клетками - кардиомиоцитами, связанными друг
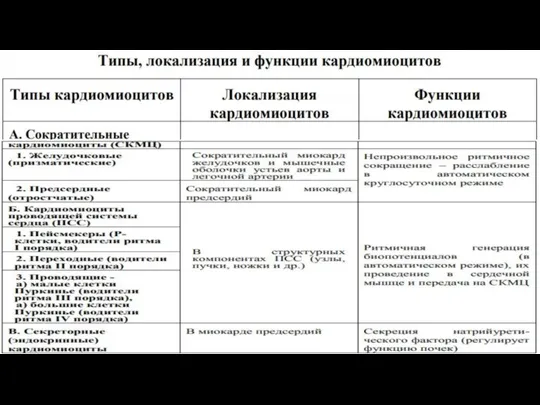
- 66. Разновидности кардиомиоцитов 1. сократительные 1) желудочковые (призматические) 2) предсердные (отростчатые) 2. кардиомиоциты проводящей системы сердца пейсмекеры
- 69. Особенности строения сократительных кардиомиоцитов (СКМЦ) Удлинённая призматическая или отростчатая форма. Размер: в длину 40-120 мкм, в
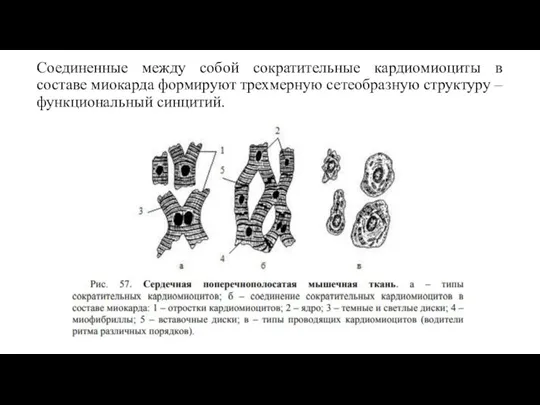
- 70. Соединенные между собой сократительные кардиомиоциты в составе миокарда формируют трехмерную сетеобразную структуру – функциональный синцитий.
- 72. Особенности строения кардиомиоцитов проводящей системы сердца (ПСС) неправильную призматическую форму в длину 8-20 мкм, в ширину
- 73. Особенности строения секреторных кардиомиоцитов отростчатая форма размер в длину 15-20 мкм, в ширину 2-5 мкм. развиты
- 74. Структурно-функциональные аппараты кардиомиоцитов
- 75. 1. Сократительный аппарат (наиболее развит в СКМЦ) Представлен миофибриллами, каждая из которых состоит из тысяч последовательно
- 76. 2. Транспортный аппарат (развит в СКМЦ) - аналогичен таковому в скелетных мышечных волокнах, но имеет ряд
- 77. 3. Опорный аппарат Представлен: Сарколеммой вставочными дисками полосками слипания Анастомозами Цитоскелетом Телофрагмами мезофрагмами. Обеспечивает формообразовательную, каркасную,
- 78. 4. Трофико-энергетический аппарат – представлен саркосомами и включениями гликогена, миоглобина и липидов. 5. Аппарат синтеза, структуризации
- 79. Регенерация сердечной мышечной ткани А. Механизмы Эндорепродукция 2. Синтез компонентов базальной мембраны 3. Пролиферация кардиомиоцитов (возможна
- 81. Нервная ткань
- 82. Функции нервной ткани 1. Восприятие раздражения 2. Генерация и проведение нервного импульса 3. Передача импульса на
- 83. Происхождение нервной ткани Возникает из участка нейроэктодермы – нервной пластинки; Нервная пластинка прогибается внутрь и образуется
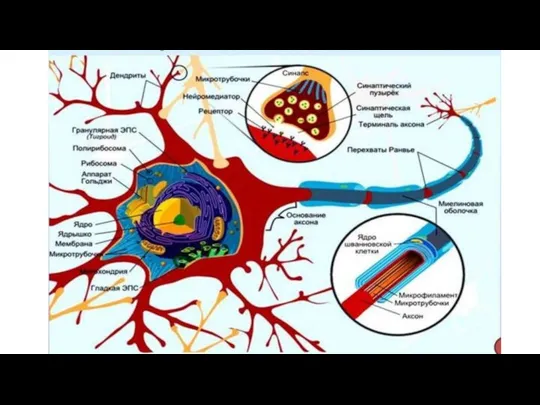
- 84. Нейрон (главная структурно-функциональная единица нервной ткани) Размеры : d = 4-5 мкм – 140 мкм Количество:
- 85. Тело нейрона Тела различных типов нейронов могут иметь круглую, овальную, уплощенную, яйцевидную или пирамидальную форму. •
- 86. Органоиды цитоплазмы нейрона Гранулярная эндоплазматическая сеть грЭПС или вещество Ниссля, или хромофильная субстанция, или тигроид -
- 87. Элементы цитоскелета • Нейрофибриллы (нф) - пучки филаментов (нейрофиламентов), диаметр которых около 3 мкм; химический состав
- 88. Отростки нейрона Аксон передает нервные импульсы на другие нейроны или клетки рабочих органов • Единственный, есть
- 89. Дендриты - отростки нейрона, которые проводят импульсы к телу нейрона, полученные от других нейронов • Количество
- 90. Классификация нейронов Морфологическая (по количеству отростков) • Униполярные – только аксон (фоторецепторы); • Биполярные – аксон
- 91. Функциональная • Чувствительные (рецепторные, сенсорные, афферентные, аффекторные) – на дендрите располагается рецептор, воспринимают раздражение и преобразуют
- 92. Классификации нейронов Функциональная: Афферентные (чувствительные) Ассоциативные (вставочные) Эфферентные (двигательные) Нейроэндокринные (секреторные) Биохимическая: Холинергические Адренергические Серотонинергические Дофаминергические
- 95. Возможно лишь одно направление проведения нервного импульса по нейрону: дендрит ? тело ? аксон
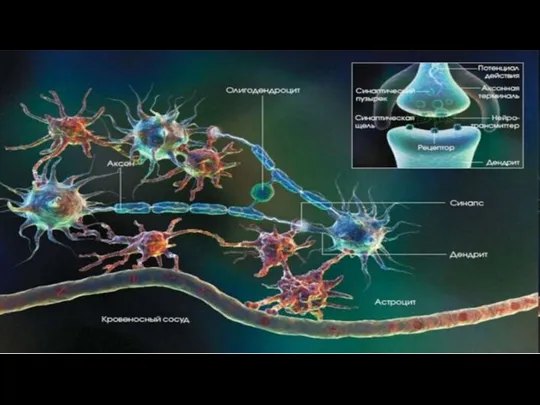
- 96. Глиоциты (нейроглия) Не проводят нервный импульс Функции гиоцитов: опорная поддержание тел и отростков нейронов, обеспечивая их
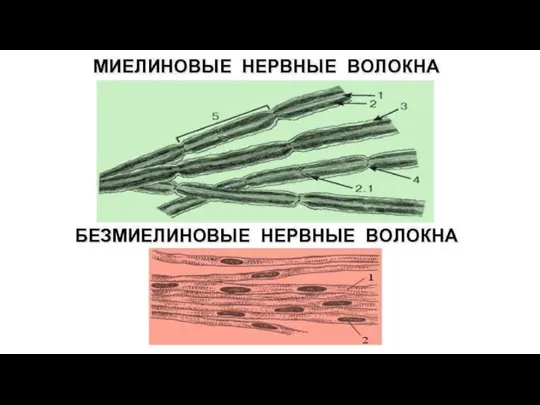
- 98. Нервные волокна В основе нервного волокна лежит отросток нервной клетки (чаще аксон) – осевой цилиндр. Каждое
- 99. Немиелинизированное волокно Не имеют миелиновой оболочки. Образуются путем погружения осевого цилиндра (аксона - А) в цитоплазму
- 100. Миелинизированное волокно Белые, имеют жироподобную миелиновую оболочку (М); Миелин – липопротеидный комплекс (холестерин, фосфолипиды, гликолипиды, белки),
- 103. Нервные окончания Концевые аппараты нервных волокон - нервные окончания - различны по их функциональному значению. Различают
- 104. Эффекторное (двигательное) нервное окончание (моторная бляшка) Моторные бляшки - комплекс взаимосвязанных структур нервной и мышечной тканей.
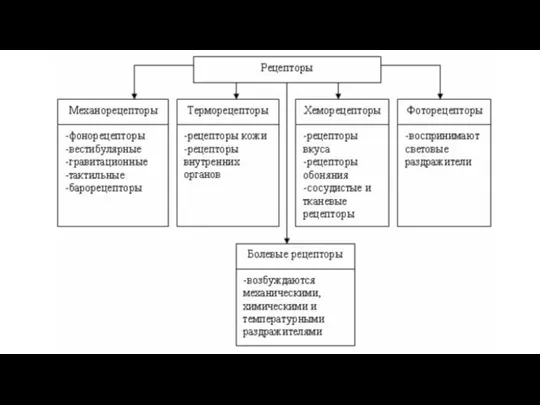
- 108. Рецепторные (чувствительные ) нервные окончания Подразделяются на: свободные нервные окончания (из терминальных ветвей дендрита чувствительной клетки)
- 110. Рецепторы кожи
- 112. Несвободное инкапсулированное нервное окончание скелетной мускулатуры (нервно-мышечное веретено) Регистрирует растяжение волокон поперечно-полосатых мышц. Представляют собой сложные
- 114. Несвободное инкапсулированное нервное окончание скелетной мускулатуры (нервно-мышечное веретено) Веретено покрыто тонкой соединительнотканной капсулой (продолжение периневрия) с
- 115. Межнейронные контакты (синапсы) Межнейрональный синапс - специализированный контакт двух нейронов, обеспечивающий одностороннее проведение нервного возбуждения. Пресинаптический
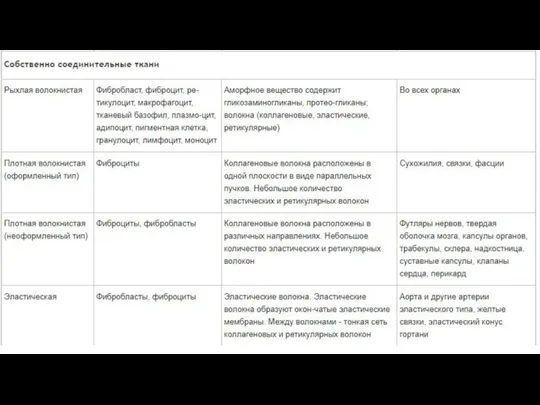
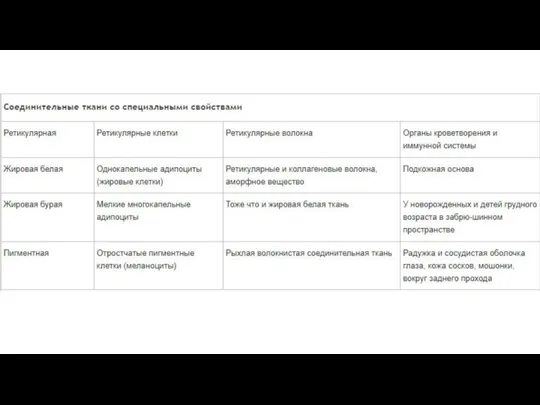
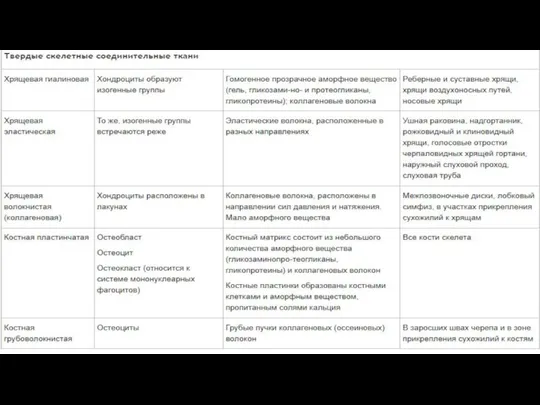
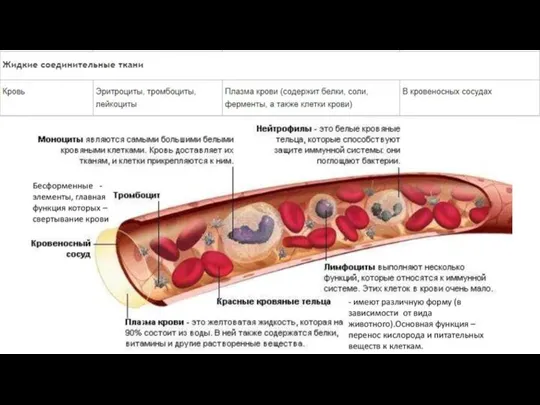
- 118. Соединительная ткань
- 121. Много межклеточного вещества, мало клеток Многообразие клеточных форм Имеются кровеносные сосуды (исключение хрящевая ткань) Общий источник
- 122. Функции соединительной ткани Опорная (соединяет клетки и ткани, создает каркас организма) Защитная (механическая, участие в иммунных
- 129. Скачать презентацию

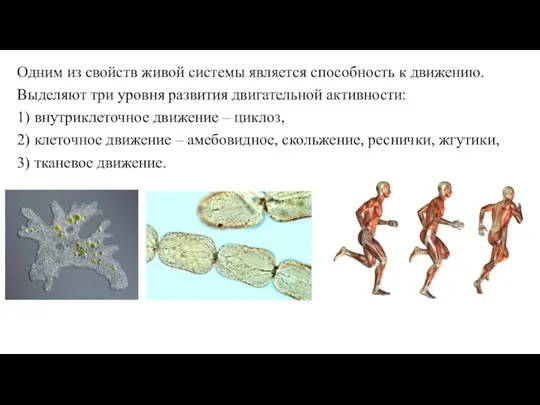
Слайд 3Одним из свойств живой системы является способность к движению.
Выделяют три уровня развития
Одним из свойств живой системы является способность к движению.
Выделяют три уровня развития
Слайд 4Мышечные ткани – это группа возбудимых тканей, различных по происхождению и строению,
Мышечные ткани – это группа возбудимых тканей, различных по происхождению и строению,
Слайд 5Общие функции мышечных тканей
1) сократительная
2) защитная
3) формообразующая
4) терморегуляция
5) трофическая
Общие функции мышечных тканей
1) сократительная
2) защитная
3) формообразующая
4) терморегуляция
5) трофическая
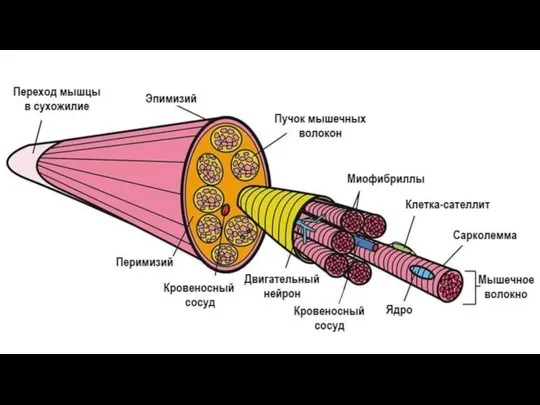
Слайд 6Общая морфофункциональная характеристика мышечных тканей
структурные элементы (клетки, волокна) обладают удлиненной формой,
наличие
Общая морфофункциональная характеристика мышечных тканей
структурные элементы (клетки, волокна) обладают удлиненной формой,
наличие

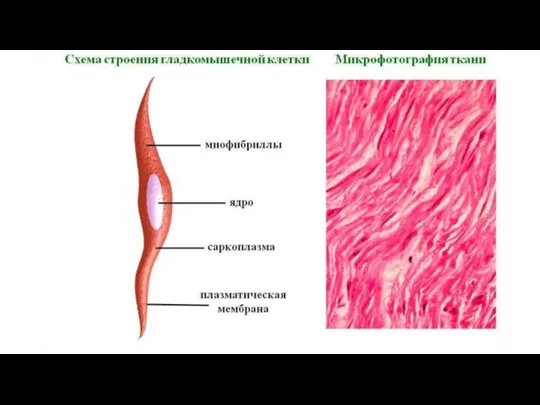
Слайд 9саркоплазма – цитоплазма мышечных волокон и клеток
сарколемма – плазмолемма
саркоплазматическая сеть
саркоплазма – цитоплазма мышечных волокон и клеток
сарколемма – плазмолемма
саркоплазматическая сеть
Слайд 10Классификация мышечных тканей
Классификация мышечных тканей основана:
на признаках их строения и функции
Классификация мышечных тканей
Классификация мышечных тканей основана:
на признаках их строения и функции

Слайд 11Морфофункциональная классификация мышечных тканей
1. Гладкие мышечные ткани – состоят из клеток, не
Морфофункциональная классификация мышечных тканей
1. Гладкие мышечные ткани – состоят из клеток, не

Слайд 13Гистогенетическая классификация мышечных тканей
Гистогенетическая классификация мышечных тканей
Слайд 14Гладкая мышечная ткань
Гладкая мышечная ткань

Слайд 15Основные места ее локализации
1) стенка сосудов
2) стенки внутренних органов пищеварительной, дыхательной, выделительной
Основные места ее локализации
1) стенка сосудов
2) стенки внутренних органов пищеварительной, дыхательной, выделительной
Слайд 16Гистогенез гладкой мышечной ткани
1 стадия – закладки – размножение стволовых клеток (СК),
Гистогенез гладкой мышечной ткани
1 стадия – закладки – размножение стволовых клеток (СК),

Слайд 17Строение гладкой мышечной ткани
Структурно-функциональной единицей гладкой мышечной ткани является гладкий миоцит (ГМ).
Строение гладкой мышечной ткани
Структурно-функциональной единицей гладкой мышечной ткани является гладкий миоцит (ГМ).

Слайд 19Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной. Щелевые контакты связывают
Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной. Щелевые контакты связывают
Слайд 21В составе ГМ выделяют следующие структурно-функциональные аппараты:
1) трофико-энергетический аппарат
2) сократительный аппарат
3)
В составе ГМ выделяют следующие структурно-функциональные аппараты:
1) трофико-энергетический аппарат
2) сократительный аппарат
3)
Слайд 221. Трофико-энергетический аппарат
Включает в себя саркосомы, включения гликогена, липидов, миоглобина. Обеспечивает энергией
1. Трофико-энергетический аппарат
Включает в себя саркосомы, включения гликогена, липидов, миоглобина. Обеспечивает энергией
Слайд 232. Сократительный аппарат
Представлен тонкими (актиновыми) и толстыми (миозиновыми) филаментами, которые не образуют
2. Сократительный аппарат
Представлен тонкими (актиновыми) и толстыми (миозиновыми) филаментами, которые не образуют

Слайд 24Сокращение гладких миоцитов индуцируется притоком в саркоплазму ионов кальция и происходит в
Сокращение гладких миоцитов индуцируется притоком в саркоплазму ионов кальция и происходит в

Слайд 253. Опорный аппарат
Включает:
Сарколемму
Цитоскелет
промежуточные филаменты
плотные тельца
межклеточные контакты (десмосомы, нексусы).
Обеспечивает:
Формообразовательную
Каркасную
локомоторную
3. Опорный аппарат
Включает:
Сарколемму
Цитоскелет
промежуточные филаменты
плотные тельца
межклеточные контакты (десмосомы, нексусы).
Обеспечивает:
Формообразовательную
Каркасную
локомоторную

Слайд 264. Аппарат внутриклеточного транспорта
К нему относятся:
кавеолы – колбовидные впячивания сарколеммы диаметром
4. Аппарат внутриклеточного транспорта
К нему относятся:
кавеолы – колбовидные впячивания сарколеммы диаметром

Слайд 275. Аппарат синтеза, структуризации и регенерации
Включает:
свободные рибосомы
ЭПС
комплекс Гольджи
лизосомы.
Обеспечивает процессы внутриклеточной
5. Аппарат синтеза, структуризации и регенерации
Включает:
свободные рибосомы
ЭПС
комплекс Гольджи
лизосомы.
Обеспечивает процессы внутриклеточной

Слайд 286. Нервный аппарат
Включает:
нервные волокна
чувствительные
двигательные нервные окончания вегетативной нервной системы.
Обеспечивает
6. Нервный аппарат
Включает:
нервные волокна
чувствительные
двигательные нервные окончания вегетативной нервной системы.
Обеспечивает
Слайд 30Регенерация гладкой мышечной ткани
А. Механизмы
1. Эндорепродукция – внутриклеточное образование, обновление и
Регенерация гладкой мышечной ткани
А. Механизмы
1. Эндорепродукция – внутриклеточное образование, обновление и

Слайд 31Б. Виды
1. Физиологическая регенерация (постэмбриональный миогенез)
• Протекает постоянно.
• Усиливается
Б. Виды
1. Физиологическая регенерация (постэмбриональный миогенез)
• Протекает постоянно.
• Усиливается

Слайд 32Гладкая мышечная ткань в составе органов
В органах гладкая мышечная ткань преимущественно представлена:
Гладкая мышечная ткань в составе органов
В органах гладкая мышечная ткань преимущественно представлена:

Слайд 33Морфофункциональные особенности гладкой мышечной ткани в зависимости от локализации в органах
Морфофункциональные особенности гладкой мышечной ткани в зависимости от локализации в органах

Слайд 35Поперечнополосатые мышечные ткани
Поперечнополосатые мышечные ткани

Слайд 36Поперечнополосатая скелетная мышечная ткань превышает по своей массе любую другую ткань организма.
Поперечнополосатая скелетная мышечная ткань превышает по своей массе любую другую ткань организма.
Слайд 37Локализация и функциональное назначение скелетной мышечной ткани
Локализация и функциональное назначение скелетной мышечной ткани

Слайд 39Гистогенез поперечнополосатой скелетной мышечной ткани
Источник развития поперечнополосатой скелетной мышечной ткани – клетки
Гистогенез поперечнополосатой скелетной мышечной ткани
Источник развития поперечнополосатой скелетной мышечной ткани – клетки


Слайд 41Строение поперечнополосатой скелетной мышечной ткани
Структурно-функциональной единицей поперечно полосатой скелетной мышечной ткани является
Строение поперечнополосатой скелетной мышечной ткани
Структурно-функциональной единицей поперечно полосатой скелетной мышечной ткани является
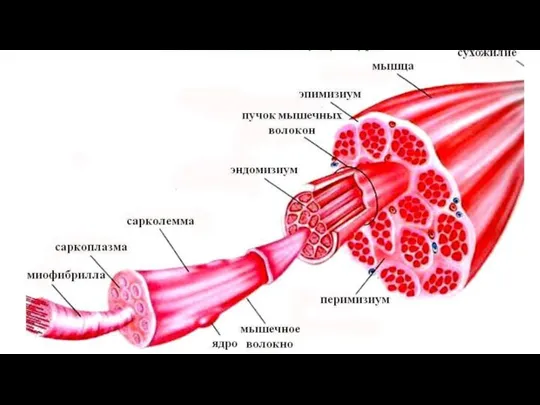
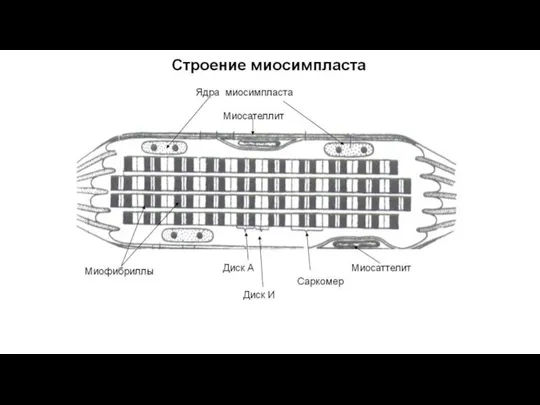
Слайд 43На светооптическом уровне после окраски в мышечном волокне наблюдается чередование участков разной
На светооптическом уровне после окраски в мышечном волокне наблюдается чередование участков разной

Слайд 44Компоненты мышечного волокна:
1) миосимпласт – занимает основной объем и ограничен сарколеммой;
Компоненты мышечного волокна:
1) миосимпласт – занимает основной объем и ограничен сарколеммой;
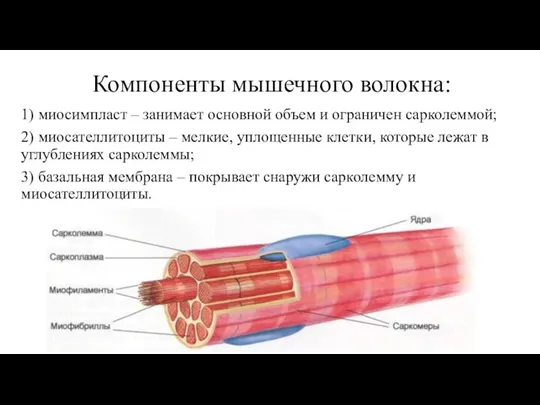
Слайд 46Структурно-функциональные аппараты мышечного волокна:
1) трофико-энергетический
2) аппарат синтеза, структуризации и регенерации
3) сократительный аппарат
4)
Структурно-функциональные аппараты мышечного волокна:
1) трофико-энергетический
2) аппарат синтеза, структуризации и регенерации
3) сократительный аппарат
4)

Слайд 47Трофико-энергетический аппарат
представлен митохондриями и трофическими включениями.
Митохондрии вырабатывают энергию, необходимую для
Трофико-энергетический аппарат
представлен митохондриями и трофическими включениями.
Митохондрии вырабатывают энергию, необходимую для

Слайд 48Аппарат синтеза, структуризации и регенерации
Основными элементами волокна, обеспечивающими процессы синтеза и
Аппарат синтеза, структуризации и регенерации
Основными элементами волокна, обеспечивающими процессы синтеза и

Слайд 49Сократительный аппарат представлен специальными органеллами – миофибриллами, которые располагаются продольно в центральной
Сократительный аппарат представлен специальными органеллами – миофибриллами, которые располагаются продольно в центральной

Слайд 50Миофибриллы имеют вид нитей диаметром 1-2 мкм.
Они обладают собственной поперечной исчерченностью
Миофибриллы имеют вид нитей диаметром 1-2 мкм.
Они обладают собственной поперечной исчерченностью

Слайд 51Транспортный аппарат
Обеспечивает депонирование и выделение ионов кальция, представлен следующими элементами: 1) поперечные
Транспортный аппарат
Обеспечивает депонирование и выделение ионов кальция, представлен следующими элементами: 1) поперечные

Слайд 52Опорный аппарат.
В его состав входят базальная мембрана, сарколемма, цитоскелет, телофрагмы, мезофрагмы, краевые
Опорный аппарат.
В его состав входят базальная мембрана, сарколемма, цитоскелет, телофрагмы, мезофрагмы, краевые

Слайд 53Нервный аппарат обеспечивает инициацию и регуляцию произвольного сокращения мышечных волокон.
Эфферентные нервные
Нервный аппарат обеспечивает инициацию и регуляцию произвольного сокращения мышечных волокон.
Эфферентные нервные

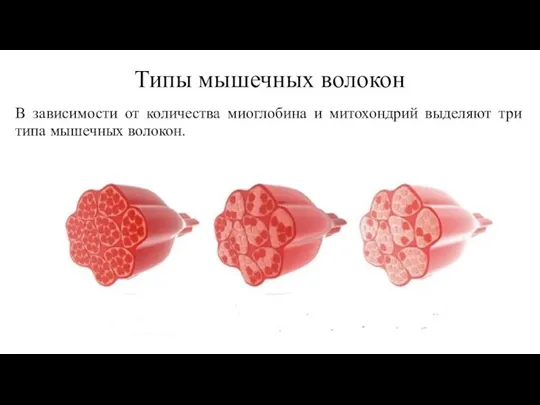
Слайд 54Типы мышечных волокон
В зависимости от количества миоглобина и митохондрий выделяют три типа
Типы мышечных волокон
В зависимости от количества миоглобина и митохондрий выделяют три типа
Слайд 55Красные (первый тип)
малый диаметр волокна
много миосателлитоцитов
тонкие миофибриллы
много миоглобина, мало
Красные (первый тип)
малый диаметр волокна
много миосателлитоцитов
тонкие миофибриллы
много миоглобина, мало

Слайд 56Белые (второй тип)
большой диаметр волокна
мало миосателлитоцитов
толстые миофибриллы
мало миоглобина, много
Белые (второй тип)
большой диаметр волокна
мало миосателлитоцитов
толстые миофибриллы
мало миоглобина, много

Слайд 57Строение скелетной мышцы как органа
Мышцы человека содержат все типы мышечных волокон.
Каждое
Строение скелетной мышцы как органа
Мышцы человека содержат все типы мышечных волокон.
Каждое

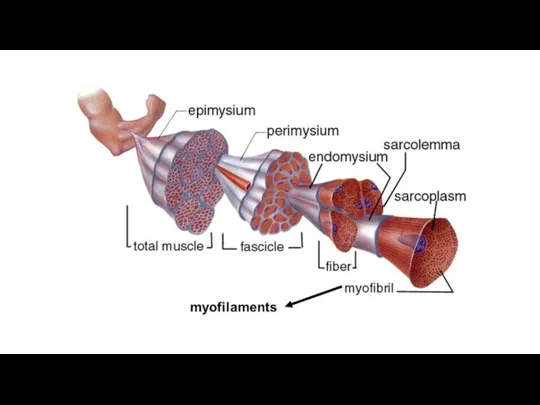
Слайд 59Регенерация скелетной поперечнополосатой мышечной ткани
А. Механизмы
1. Эндорепродукция – внутрисимпластическое образование, обновление
Регенерация скелетной поперечнополосатой мышечной ткани
А. Механизмы
1. Эндорепродукция – внутрисимпластическое образование, обновление

Слайд 60Б. Виды
Физиологическая регенерация (постэмбриональный миогенез)
• Протекает постоянно, состоит в самообновлении
Б. Виды
Физиологическая регенерация (постэмбриональный миогенез)
• Протекает постоянно, состоит в самообновлении

Слайд 61Сердечная мышечная ткань
Сердечная мышечная ткань
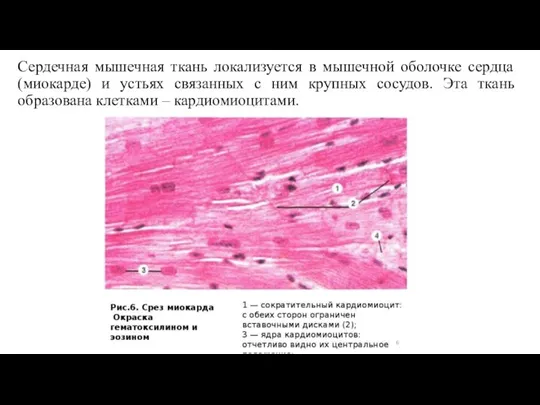
Слайд 62Сердечная мышечная ткань локализуется в мышечной оболочке сердца (миокарде) и устьях связанных
Сердечная мышечная ткань локализуется в мышечной оболочке сердца (миокарде) и устьях связанных
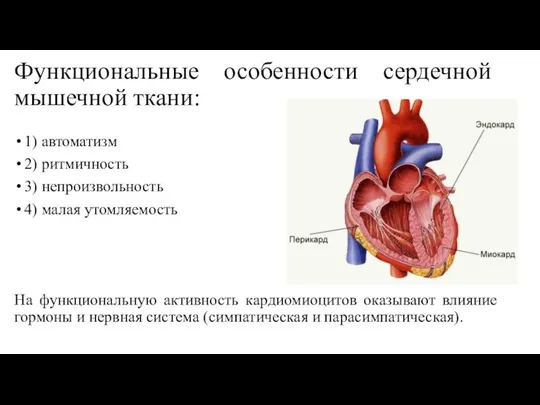
Слайд 63Функциональные особенности сердечной мышечной ткани:
1) автоматизм
2) ритмичность
3) непроизвольность
4) малая утомляемость
На функциональную активность
Функциональные особенности сердечной мышечной ткани:
1) автоматизм
2) ритмичность
3) непроизвольность
4) малая утомляемость
На функциональную активность
Слайд 64Гистогенез сердечной мышечной ткани
Источником развития сердечной мышечной ткани является миоэпикардиальная пластинка висцерального
Гистогенез сердечной мышечной ткани
Источником развития сердечной мышечной ткани является миоэпикардиальная пластинка висцерального

Слайд 65Строение сердечной мышечной ткани
Как уже отмечалось, сердечная мышечная ткань образована клетками -
Строение сердечной мышечной ткани
Как уже отмечалось, сердечная мышечная ткань образована клетками -

Слайд 66Разновидности кардиомиоцитов
1. сократительные
1) желудочковые (призматические)
2) предсердные (отростчатые)
2. кардиомиоциты проводящей
Разновидности кардиомиоцитов
1. сократительные
1) желудочковые (призматические)
2) предсердные (отростчатые)
2. кардиомиоциты проводящей

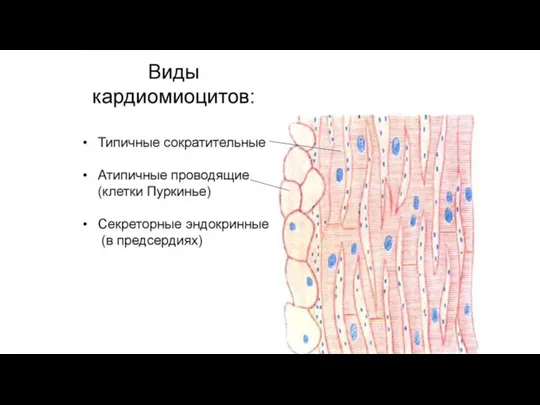
Слайд 69Особенности строения сократительных кардиомиоцитов (СКМЦ)
Удлинённая призматическая или отростчатая форма.
Размер: в длину
Особенности строения сократительных кардиомиоцитов (СКМЦ)
Удлинённая призматическая или отростчатая форма.
Размер: в длину

Слайд 70Соединенные между собой сократительные кардиомиоциты в составе миокарда формируют трехмерную сетеобразную структуру
Соединенные между собой сократительные кардиомиоциты в составе миокарда формируют трехмерную сетеобразную структуру

Слайд 72Особенности строения кардиомиоцитов проводящей системы сердца (ПСС)
неправильную призматическую форму
в длину 8-20 мкм,
Особенности строения кардиомиоцитов проводящей системы сердца (ПСС)
неправильную призматическую форму
в длину 8-20 мкм,

Слайд 73Особенности строения секреторных кардиомиоцитов
отростчатая форма
размер в длину 15-20 мкм, в ширину 2-5
Особенности строения секреторных кардиомиоцитов
отростчатая форма
размер в длину 15-20 мкм, в ширину 2-5

Слайд 74Структурно-функциональные аппараты кардиомиоцитов
Структурно-функциональные аппараты кардиомиоцитов

Слайд 751. Сократительный аппарат (наиболее развит в СКМЦ)
Представлен миофибриллами, каждая из которых состоит
1. Сократительный аппарат (наиболее развит в СКМЦ)
Представлен миофибриллами, каждая из которых состоит

Слайд 762. Транспортный аппарат (развит в СКМЦ) - аналогичен таковому в скелетных мышечных
2. Транспортный аппарат (развит в СКМЦ) - аналогичен таковому в скелетных мышечных

Слайд 773. Опорный аппарат
Представлен:
Сарколеммой
вставочными дисками
полосками слипания
Анастомозами
Цитоскелетом
Телофрагмами
мезофрагмами.
Обеспечивает формообразовательную, каркасную, локомоторную и интеграционную
3. Опорный аппарат
Представлен:
Сарколеммой
вставочными дисками
полосками слипания
Анастомозами
Цитоскелетом
Телофрагмами
мезофрагмами.
Обеспечивает формообразовательную, каркасную, локомоторную и интеграционную

Слайд 784. Трофико-энергетический аппарат – представлен саркосомами и включениями гликогена, миоглобина и липидов.
4. Трофико-энергетический аппарат – представлен саркосомами и включениями гликогена, миоглобина и липидов.

Слайд 79Регенерация сердечной мышечной ткани
А. Механизмы
Эндорепродукция
2. Синтез компонентов базальной мембраны
3.
Регенерация сердечной мышечной ткани
А. Механизмы
Эндорепродукция
2. Синтез компонентов базальной мембраны
3.


Слайд 81Нервная ткань
Нервная ткань
Слайд 82Функции нервной ткани
1. Восприятие раздражения
2. Генерация и проведение нервного импульса
3. Передача
Функции нервной ткани
1. Восприятие раздражения
2. Генерация и проведение нервного импульса
3. Передача
Слайд 83Происхождение нервной ткани
Возникает из участка нейроэктодермы – нервной пластинки;
Нервная пластинка прогибается
Происхождение нервной ткани
Возникает из участка нейроэктодермы – нервной пластинки;
Нервная пластинка прогибается

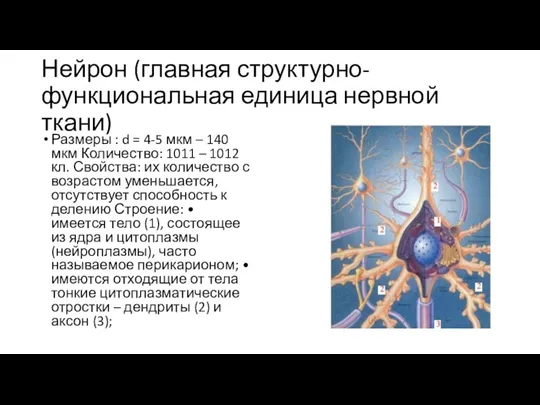
Слайд 84Нейрон (главная структурно-функциональная единица нервной ткани)
Размеры : d = 4-5 мкм –
Нейрон (главная структурно-функциональная единица нервной ткани)
Размеры : d = 4-5 мкм –
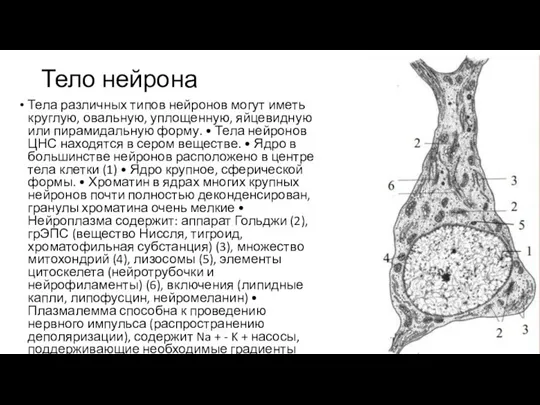
Слайд 85Тело нейрона
Тела различных типов нейронов могут иметь круглую, овальную, уплощенную, яйцевидную или
Тело нейрона
Тела различных типов нейронов могут иметь круглую, овальную, уплощенную, яйцевидную или
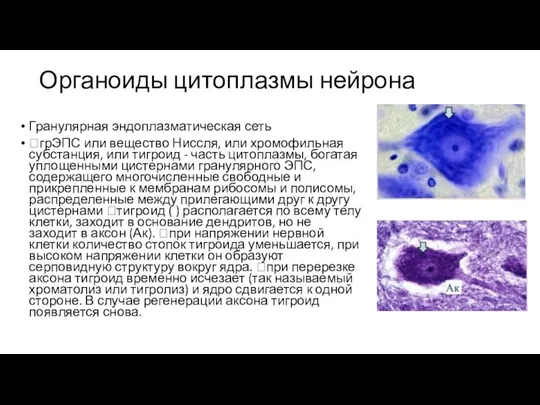
Слайд 86Органоиды цитоплазмы нейрона
Гранулярная эндоплазматическая сеть
грЭПС или вещество Ниссля, или хромофильная субстанция, или
Органоиды цитоплазмы нейрона
Гранулярная эндоплазматическая сеть
грЭПС или вещество Ниссля, или хромофильная субстанция, или
Слайд 87Элементы цитоскелета
• Нейрофибриллы (нф) - пучки филаментов (нейрофиламентов), диаметр которых около 3
Элементы цитоскелета
• Нейрофибриллы (нф) - пучки филаментов (нейрофиламентов), диаметр которых около 3

Слайд 88Отростки нейрона
Аксон передает нервные импульсы на другие нейроны или клетки рабочих органов
Отростки нейрона
Аксон передает нервные импульсы на другие нейроны или клетки рабочих органов

Слайд 89Дендриты - отростки нейрона, которые проводят импульсы к телу нейрона, полученные от
Дендриты - отростки нейрона, которые проводят импульсы к телу нейрона, полученные от

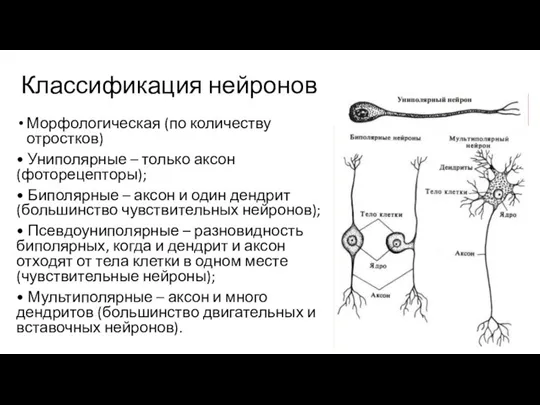
Слайд 90Классификация нейронов
Морфологическая (по количеству отростков)
• Униполярные – только аксон (фоторецепторы);
•
Классификация нейронов
Морфологическая (по количеству отростков)
• Униполярные – только аксон (фоторецепторы);
•
Слайд 91Функциональная
• Чувствительные (рецепторные, сенсорные, афферентные, аффекторные) – на дендрите располагается рецептор, воспринимают
Функциональная
• Чувствительные (рецепторные, сенсорные, афферентные, аффекторные) – на дендрите располагается рецептор, воспринимают

Слайд 92Классификации нейронов
Функциональная:
Афферентные (чувствительные)
Ассоциативные (вставочные)
Эфферентные (двигательные)
Нейроэндокринные (секреторные)
Биохимическая:
Холинергические
Адренергические
Серотонинергические
Классификации нейронов
Функциональная:
Афферентные (чувствительные)
Ассоциативные (вставочные)
Эфферентные (двигательные)
Нейроэндокринные (секреторные)
Биохимическая:
Холинергические
Адренергические
Серотонинергические


Слайд 95Возможно лишь одно направление проведения нервного импульса по нейрону: дендрит ? тело
Возможно лишь одно направление проведения нервного импульса по нейрону: дендрит ? тело

Слайд 96Глиоциты (нейроглия)
Не проводят нервный импульс
Функции гиоцитов:
опорная
поддержание тел и отростков
Глиоциты (нейроглия)
Не проводят нервный импульс
Функции гиоцитов:
опорная
поддержание тел и отростков

Слайд 98Нервные волокна
В основе нервного волокна лежит отросток нервной клетки (чаще аксон) –
Нервные волокна
В основе нервного волокна лежит отросток нервной клетки (чаще аксон) –

Слайд 99Немиелинизированное волокно
Не имеют миелиновой оболочки.
Образуются путем погружения осевого цилиндра (аксона
Немиелинизированное волокно
Не имеют миелиновой оболочки.
Образуются путем погружения осевого цилиндра (аксона

Слайд 100Миелинизированное волокно
Белые, имеют жироподобную миелиновую оболочку (М);
Миелин – липопротеидный комплекс (холестерин,
Миелинизированное волокно
Белые, имеют жироподобную миелиновую оболочку (М);
Миелин – липопротеидный комплекс (холестерин,


Слайд 103Нервные окончания
Концевые аппараты нервных волокон - нервные окончания - различны по их
Нервные окончания
Концевые аппараты нервных волокон - нервные окончания - различны по их

Слайд 104Эффекторное (двигательное) нервное окончание (моторная бляшка)
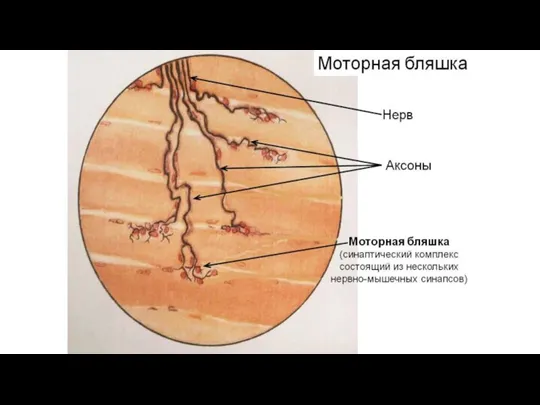
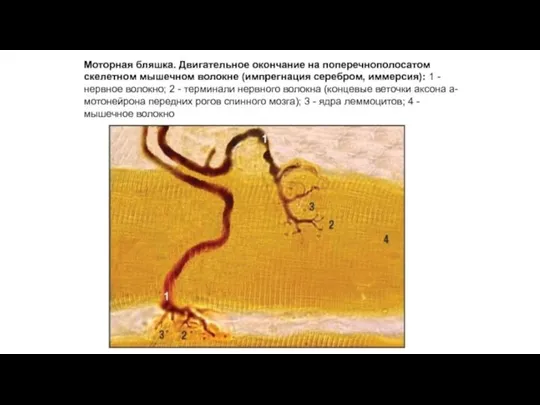
Моторные бляшки - комплекс взаимосвязанных структур нервной
Эффекторное (двигательное) нервное окончание (моторная бляшка)
Моторные бляшки - комплекс взаимосвязанных структур нервной

Слайд 108Рецепторные (чувствительные ) нервные окончания
Подразделяются на:
свободные нервные окончания (из терминальных ветвей
Рецепторные (чувствительные ) нервные окончания
Подразделяются на:
свободные нервные окончания (из терминальных ветвей

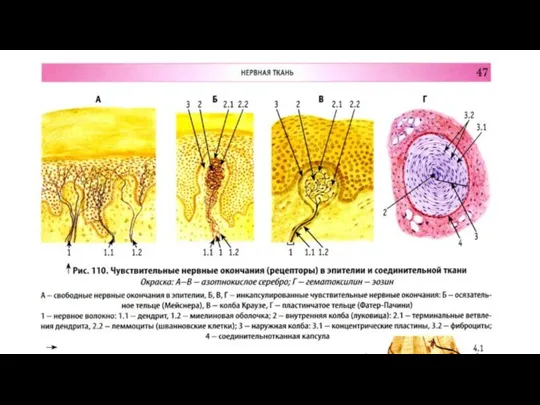
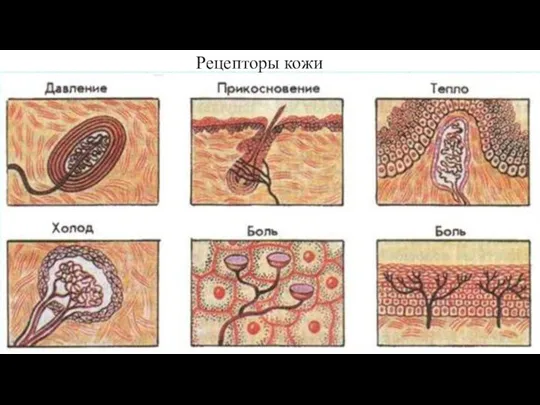
Слайд 110Рецепторы кожи
Рецепторы кожи
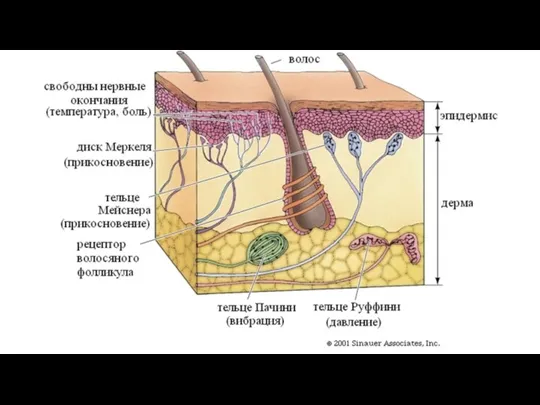
Слайд 112Несвободное инкапсулированное нервное окончание скелетной мускулатуры (нервно-мышечное веретено)
Регистрирует растяжение волокон поперечно-полосатых мышц.
Несвободное инкапсулированное нервное окончание скелетной мускулатуры (нервно-мышечное веретено)
Регистрирует растяжение волокон поперечно-полосатых мышц.

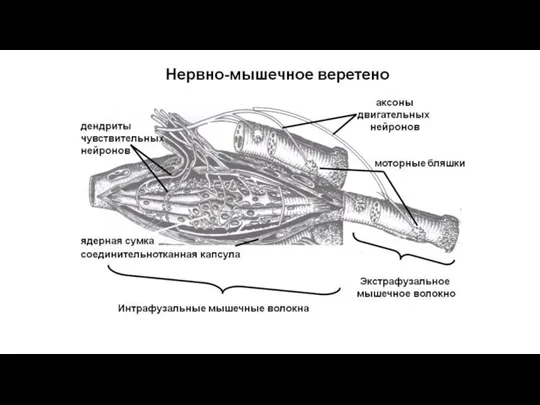
Слайд 114Несвободное инкапсулированное нервное окончание скелетной мускулатуры (нервно-мышечное веретено)
Веретено покрыто тонкой соединительнотканной капсулой
Несвободное инкапсулированное нервное окончание скелетной мускулатуры (нервно-мышечное веретено)
Веретено покрыто тонкой соединительнотканной капсулой

Слайд 115Межнейронные контакты (синапсы)
Межнейрональный синапс - специализированный контакт двух нейронов, обеспечивающий одностороннее проведение
Межнейронные контакты (синапсы)
Межнейрональный синапс - специализированный контакт двух нейронов, обеспечивающий одностороннее проведение

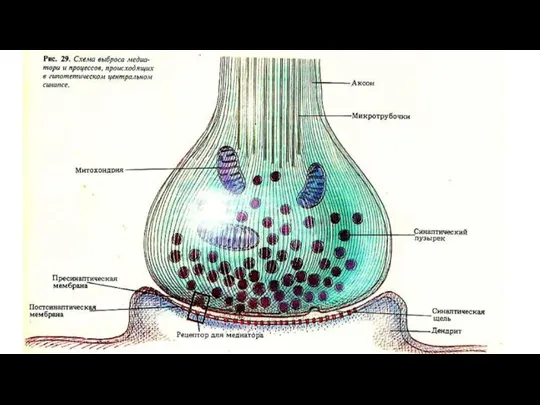

Слайд 118Соединительная ткань
Соединительная ткань

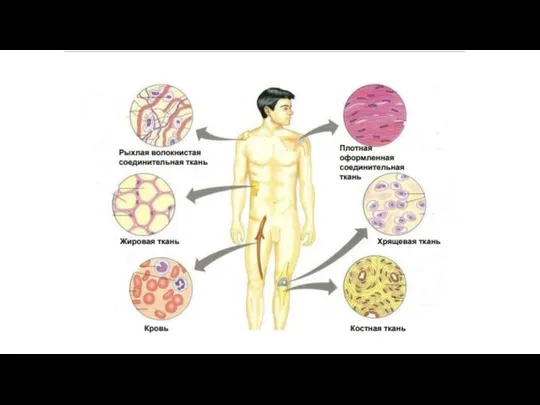
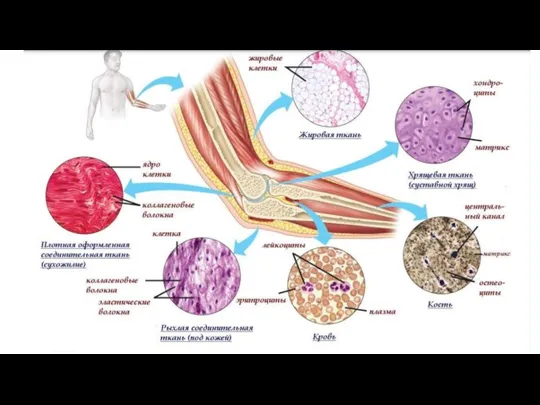
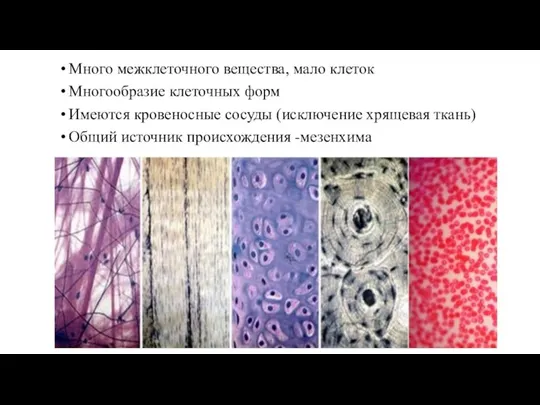
Слайд 121Много межклеточного вещества, мало клеток
Многообразие клеточных форм
Имеются кровеносные сосуды (исключение
Много межклеточного вещества, мало клеток
Многообразие клеточных форм
Имеются кровеносные сосуды (исключение
Слайд 122Функции соединительной ткани
Опорная (соединяет клетки и ткани, создает каркас организма)
Защитная (механическая,
Функции соединительной ткани
Опорная (соединяет клетки и ткани, создает каркас организма)
Защитная (механическая,

 pril
pril Разнообразие беспозвоночных животных. Викторина
Разнообразие беспозвоночных животных. Викторина Кислотно-основный состав крови
Кислотно-основный состав крови методы иссл
методы иссл Презентация на тему Методы изучения наследственности человека
Презентация на тему Методы изучения наследственности человека  Механические ткани в органах растений
Механические ткани в органах растений Тираннозавр (ящер-тиран)
Тираннозавр (ящер-тиран) Мышечная рецепция
Мышечная рецепция Тип Круглые черви
Тип Круглые черви Они тоже сражались за родину. Животный мир и война
Они тоже сражались за родину. Животный мир и война Корнеплоды и клубнеплоды
Корнеплоды и клубнеплоды Виды рыб Байкала
Виды рыб Байкала Окружающий мир. Животные
Окружающий мир. Животные Царство Грибы. Часть 1
Царство Грибы. Часть 1 Основные свойства жизни
Основные свойства жизни Домашние животные
Домашние животные Эмбриология. Строение половых клеток
Эмбриология. Строение половых клеток Клеточная теория
Клеточная теория Клетка. Разновидности клеток
Клетка. Разновидности клеток Животный мир Арктики
Животный мир Арктики Семена. Сортовой контроль
Семена. Сортовой контроль Строение головного мозга. Функции продолговатого мозга, моста, мозжечка и среднего мозга
Строение головного мозга. Функции продолговатого мозга, моста, мозжечка и среднего мозга Минеральное питание растений и значение воды
Минеральное питание растений и значение воды Побег, его строение и развитие
Побег, его строение и развитие Анатомия человека. Принципы организации тела человека
Анатомия человека. Принципы организации тела человека русский язык (FILEminimizer)
русский язык (FILEminimizer) konechnosti
konechnosti