- Мышечная ткань

Содержание
- 2. План лекции Определение, происхождение и классификация мышечной ткани Строение, функции, регенерация гладкой мышечной ткани Строение, функции,
- 3. Мышечные ткани (МТ) представляют собой группу тканей различного происхождения и строения, но объединенных способностью к сокращению
- 4. Общая морфофункциональная характеристика мышечной ткани (МТ) Структурные элементы (клетки, волокна) обладают удлиненной формой; Наличие органелл специального
- 5. Классификация МТ 1. по морфофункциональной характеристике 1. Гладкая мышечная ткань (ГМТ) 2. Поперечно-полосатая мышечная ткань (ППМТ)
- 6. КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ТКАНЕЙ 2. Гистогенетическая классификация мышечных тканей ∙ Эктодермальные − эпителиально-мышечная ткань (мышечные клетки экзокринных
- 7. РАЗВИТИЕ МЫШЕЧНОЙ ТКАНИ
- 8. Классификация предложенная Николаем Григорьевичем Хлопиным: 1. Гладкая МТ. 2. Поперечно-полосатая МТ. 1)Поперечно-полосатая МТ соматического типа. 2)Поперечно-полосатая
- 9. Мышцы составляют большую массу тела и входят в состав многих органов. ТЕРМИНОЛОГИЯ: мышечная клетка = мышечное
- 10. Гладкая МТ входит в состав мышечных оболочек сосудов, кишечника, мочевыводящих, семявыводящих путей; обнаруживается в селезенке, коже
- 11. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ Микрофотография ткани ядро миофибриллы саркоплазма плазматическая мембрана Схема строения гладкомышечной клетки
- 12. Миоцит представляет собой чаще всего веретенообразную клетку (длиной 20—1000 мкм, диаметром 2—20 мкм). В матке клетки
- 13. Характеристика гадкого миоцита 9. Сарколемма ГМ окружена базальной мембраной, в которой вплетаются тонкие ретикулярные, коллагеновые, эластические
- 14. 11. Актиновые миофиламенты: располагаются продольно или под углом образуют трехмерную сеть в месте их контакта друг
- 15. Гладкая мышечная ткань продольный срез поперечный срез
- 16. СТРОЕНИЕ ГЛАДКОМЫШЕЧНОЙ КЛЕТКИ (схема) митохондрии гликоген ядро грЭПС аппарат Гольджи периферические пузырьки базальная пластина
- 17. Структурная единица – гладкий миоцит Мио-миоцитарный контакт миофиламенты митохондрия плотные тельца клеточный центр ядро миолемма эластические
- 18. В гладкой МТ толстые и тонкие филаменты ориентированы под углом к осям волокна и прикреплены к
- 19. Строение гладкого миоцита Фаза расслабления Фаза сокращения Плотные тельца: Плотные тельца Промежуточные филаменты Промежуточные филаменты ядро
- 20. Ядро в центре клетки, а периферическая часть клетки занята сократительным аппаратом: много актиновых (их больше) +
- 21. Гладкие мышцы Поперечные срезы гладких миоцитов Продольные срезы гладких миоцитов Перимизий Перимизий Эндомизий Межклеточное вещество Межклеточное
- 22. Гладкие мышечные ткани. Нейральные – Миоциты расположены пучками, Эндомизий и Перимизий тонкие, содержат Меланоциты и миопигментоциты
- 23. Особые типы мышечных клеток Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности. Окружают секреторные отделы и
- 24. Мионейральные клетки Мионейральная ткань входит в состав мышц радужной оболочки глаза – мышцы, суживающей зрачок, и
- 25. В миоидных клетках (миофибробластах) ядра вытянутые, палочковидные Миофибробласты относятся к рыхлой соединительной ткани и представляют собой
- 26. Эндокринные гладкие миоциты Эндокринные гладкие миоциты – это видоизмененные ГМК, представляющие основной компонент юкстагломерулярного аппарата почек.
- 27. Регенерация гладкой мышечной ткани осуществляется несколькими способами: посредством внутриклеточной регенерации гипертрофии при усилении функциональной нагрузки; посредством
- 28. 3. Строение, функции, регенерация поперечно-полосатой мышечной ткани (соматической)
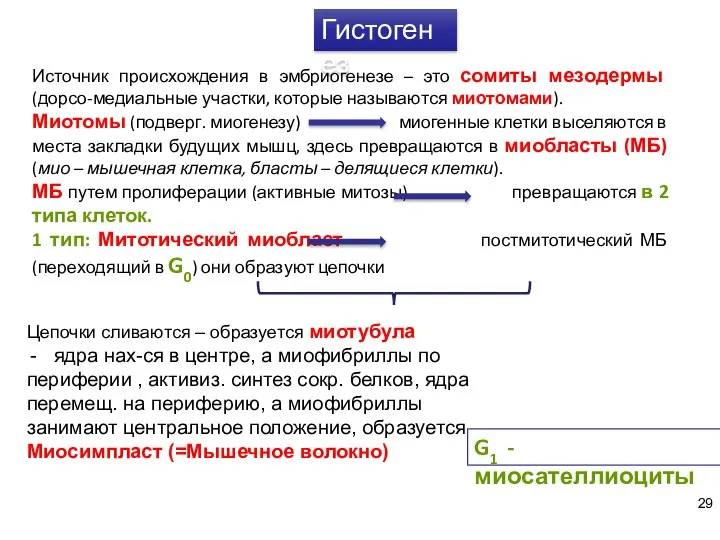
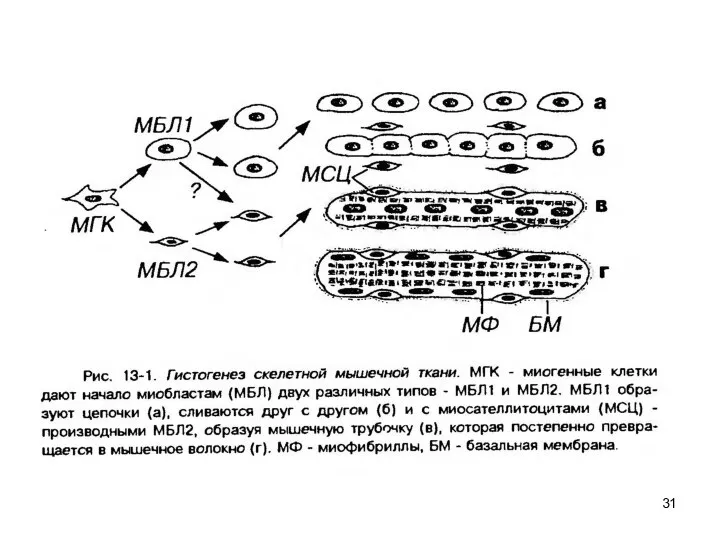
- 29. Гистогенез Источник происхождения в эмбриогенезе – это сомиты мезодермы (дорсо-медиальные участки, которые называются миотомами). Миотомы (подверг.
- 30. Миотомы и развитие системы мышц.
- 32. 2 тип: Митотический миобласт переходит в состояние G1 – готовы всегда к делению – это миосателлиты
- 33. Скелетная мышечная ткань (соматического типа). Основные гистологические элементы: скелетные мышечные волокна (функция сокращения) и клетки-сателлиты (камбиальный
- 34. Строение скелетной мышечной ткани Рассмотрим строение мышечной ткани на нескольких уровнях организации живого: - на органном
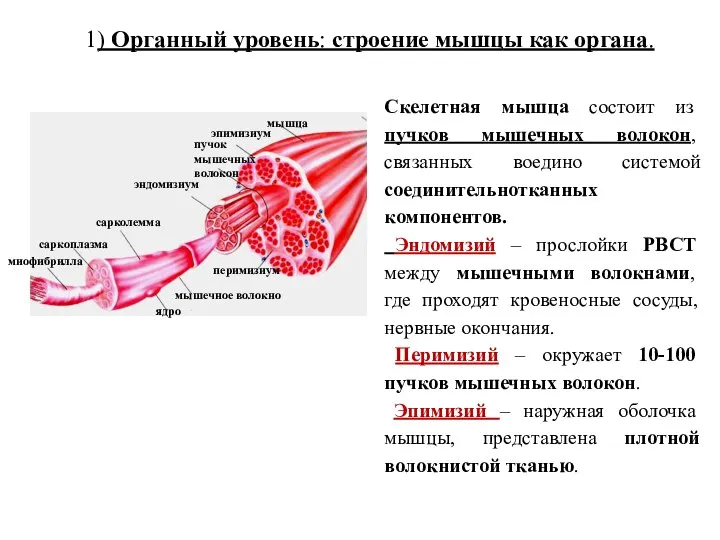
- 35. 1) Органный уровень: строение мышцы как органа. Скелетная мышца состоит из пучков мышечных волокон, связанных воедино
- 36. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ Анатомия скелетной мускулатуры сухожилие мышца эпимизиум пучок мышечных волокон перимизиум
- 37. Структурно-функциональной единицей скелетной поперечнополосатой (исчерченной) мышечной ткани является мышечное волокно (симпласт) – цилиндрической формы образование диаметром
- 38. Клеточный уровень - строение мышечного волокна (миосимпласта). Термин «мышечное волокно» подразумевает «миосимпласт», поскольку миосимпласт обеспечивает функцию
- 39. Субклеточный уровень: строение миофибриллы I-диск содержит только тонкие нити. В середине I-диска проходит Z-линия - которая
- 40. Субклеточный уровень: строение миофибриллы. Саркомер содержит один А-диск (тёмный) и две половины I-диска (светлого). Толстые миозиновые
- 41. В мышечном волокне (миосимпласте) различают функциональные аппараты 1. Мембранный 2. Фибриллярный (сократительный) и 3. Трофический. Трофический
- 42. Каждая миофибрилла окружена регулярно повторяющимися элементами сaркоплазматического ретикулума - анастомозирующими мембранными трубочками (L-трубочки), заканчивающимися терминальными цистернами.
- 43. Молекулярный уровень: строение актиновых и миозиновых филаментов. Под электронным микроскопом миофибриллы представляют агрегаты из толстых, или
- 44. Тяжелые цепи состоят из шести полипептидных цепей, две из них тяжелые и представляют собой две спирально
- 45. МИКРОФИЛАМЕНТЫ Миозин как представитель микрофиламентов тяжелая цепь миозина Схема строения молекулы миозина тяжелая цепь миозина легкие
- 46. : Тонкие филаменты, или актиновые нити, (диаметр 7-8 нм), образованы тремя белками: актином, тропонином и тропомиозином.
- 47. Молекулярный уровень: строение актиновых филаментов В покое взаимодействие тонких и толстых нитей (миофиламентов) невозможно, т.к. миозин-связывающие
- 48. саркомер 1 — плазмолемма; 2 — саркоплазма; 3 — ядра миосимпласта; 4 — миофибриллы; 5 —
- 49. МИКРОФИЛАМЕНТЫ Структура акто-миозиновых комплексов Схема строения саркомера Z диск титин тропомодулин М линия миозиновая нить Z
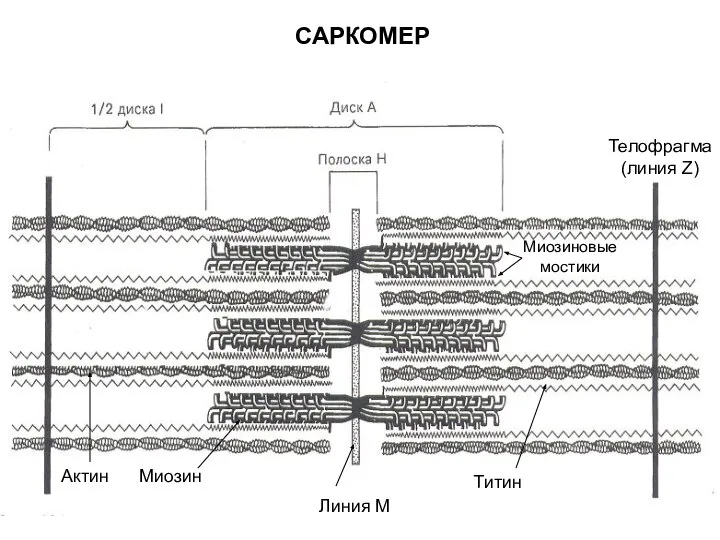
- 50. САРКОМЕР Линия М Миозин Актин Телофрагма (линия Z) Титин Миозиновые мостики
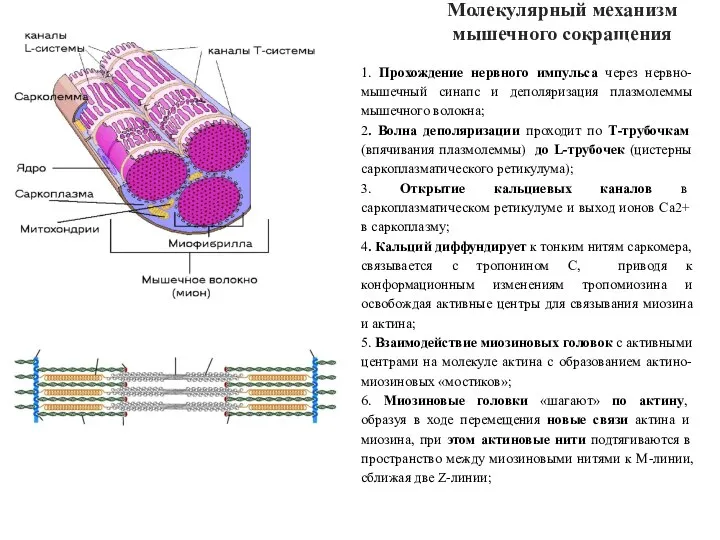
- 51. Молекулярный механизм мышечного сокращения 1. Прохождение нервного импульса через нервно-мышечный синапс и деполяризация плазмолеммы мышечного волокна;
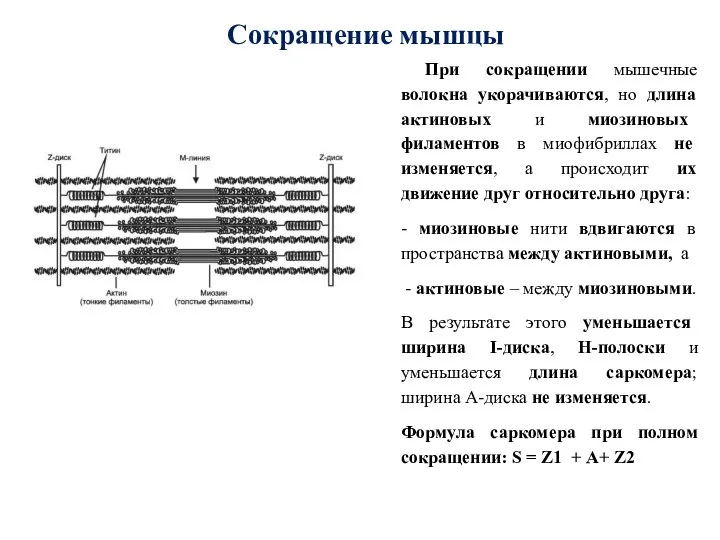
- 52. Сокращение мышцы При сокращении мышечные волокна укорачиваются, но длина актиновых и миозиновых филаментов в миофибриллах не
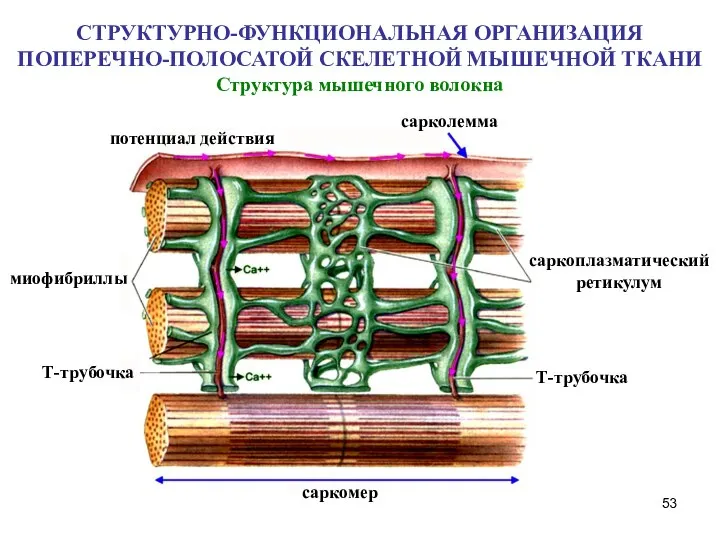
- 53. Структура мышечного волокна СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ потенциал действия сарколемма саркоплазматический ретикулум Т-трубочка Т-трубочка
- 54. Актин Механизм сокращения Головка миозинового мостика Миозиновый мостик Отсутствие связи миозина с актином Акто-миозиновый комплекс Изменение
- 55. Структурные элементы саркомера (S) в расслабленном состоянии можно выразить формулой: S = Z1 +1/2I1 +1/2A1 +
- 56. Стадия расслабления Стадия сокращения Саркомер миофибриллы
- 57. Скелетная мышечная ткань Эндомизий Миосимпласт в поперечном срезе Миосимпласт в продольном срезе Саркоплазма Сарколемма Ядра миосимпластов
- 58. Микрофотография продольного среза поперечно-полосатой мышечной ткани мышечные волокна ядра СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
- 59. Скелетная мышечная ткань
- 60. НЕРВНО-МЫШЕЧНЫЙ КОНТАКТ
- 61. Типы мышц: КРАСНЫЕ БЕЛЫЕ ПЕРЕХОДНЫЕ Больше миоглобина и митохондрий, меньше гликогена и миофибрилл Меньше миоглобина и
- 62. ТИПЫ МЫШЕЧНЫХ ВОЛОКОН Красные (окислительные) мышечные волокна небольшого диаметра, окружены массой капилляров, содержат много миоглобина. Их
- 63. Красные и белые мионы Различия белых и красных мионов
- 64. ИННЕРВАЦИЯ: Скелетные мышцы получают двигательную, чувствительную и трофическую (вегетативную) иннервацию. Двигательную (эфферентную) иннервацию скелетные мышцы туловища
- 65. КРОВОСНАБЖЕНИЕ Скелетные мышцы богато снабжаются кровью. В рыхлой соединительной ткани перимизия в большом количестве содержатся артерии
- 66. Рабочие кардиомиоциты. Гематоксилин – эозин, 200х 3. Поперечно-полосатая сердечная мышечная ткань
- 67. Поперечнополосатые мышечные ткани сердечного типа 2. Сердечная мышечная ткань (целомического типа) Целомический (сердечный)----Висцеральный листок спланхнотома (миоэпикардиальные
- 68. Сердечная мышечная ткань Продольные срезы мышечных волокон Поперечные срезы мышечных волокон Эндомизий с сосудами МИОКАРД
- 69. СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ. Встречается в стенке сердца, проксимальной части аорты, в верхней полой вене. Структурная единица
- 70. КАРДИОМИОЦИТЫ Клетки расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры бассейна венечных сосудов
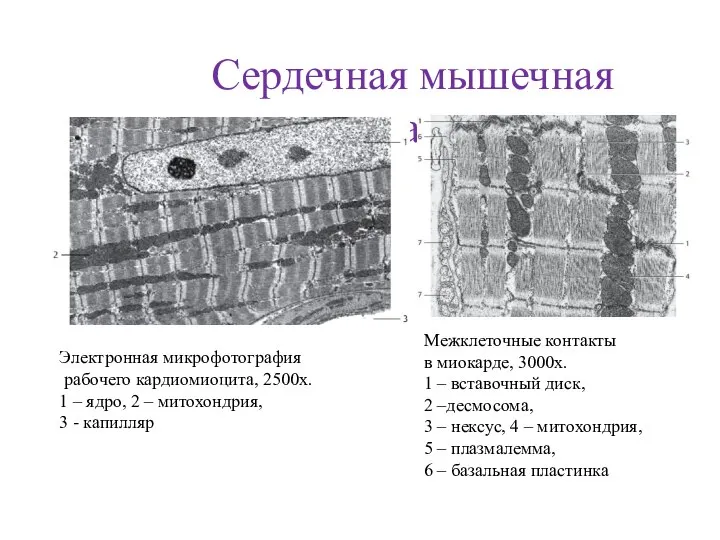
- 71. Сердечная мышечная ткань Электронная микрофотография рабочего кардиомиоцита, 2500х. 1 – ядро, 2 – митохондрия, 3 -
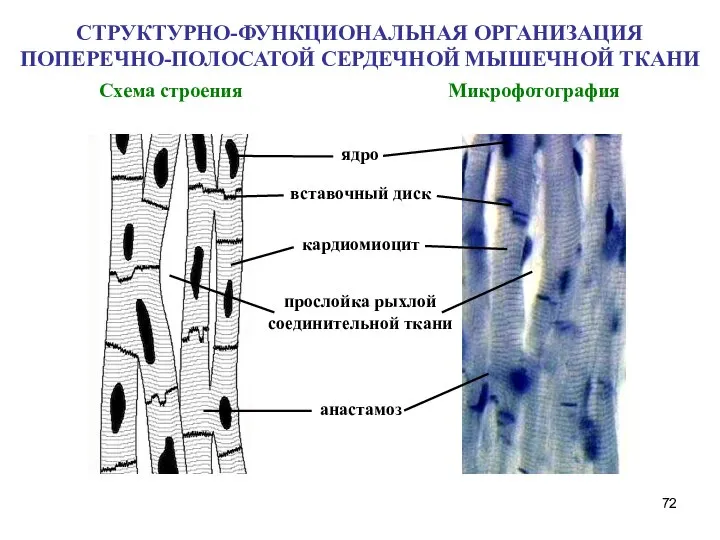
- 72. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ Схема строения Микрофотография ядро вставочный диск кардиомиоцит анастамоз прослойка рыхлой
- 73. Типичные кардиомиоциты (рабочие, сократительные) Типичные (рабочие, сократительные) кардиомиоциты – клетки цилиндрической формы, длиной до 100-150 мкм
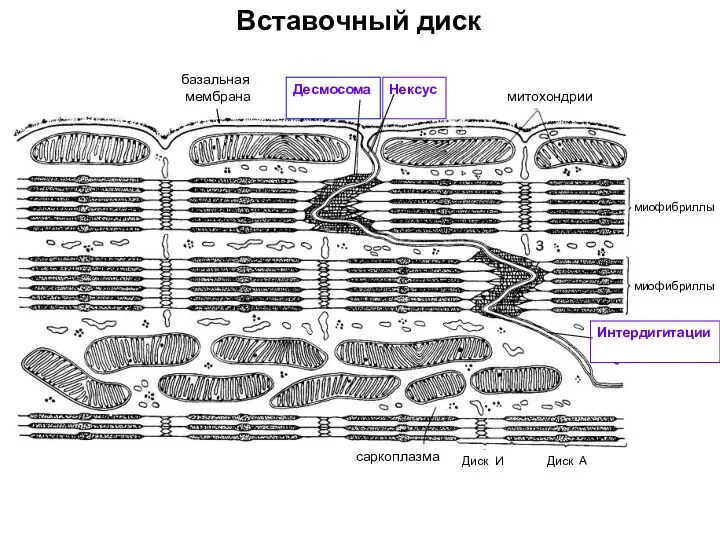
- 74. Вставочный диск базальная мембрана Десмосома Нексус митохондрии миофибриллы миофибриллы саркоплазма Диск И Диск А Интердигитации
- 75. Мембранный аппарат Мембранный аппарат: каждая клетка покрыта оболочкой, состоящей из комплекса плазмолеммы и базальной мембраны. Оболочка
- 76. Фибриллярный (сократительный) аппарат. Фибриллярный (сократительный) аппарат. Большую часть цитоплазмы кардиомиоцита занимают органеллы специального назначения – миофибриллы,
- 77. Сократительные кардиомиоциты Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются
- 78. Предсердные и желудочковые кардиомиоциты Предсердные и желудочковые кардиомиоциты относятся к разным популяциям рабочих кардиомиоцитов. Предсердные кардиомиоциты
- 79. Секреторные кардиомиоциты Секреторные кардиомиоциты выполняют особую функцию. Все кардиомиоциты покрыты базальной мембраной. В части кардиомиоцитов предсердий
- 80. Проводящие (атипичные) кардиомиоциты Проводящие (атипичные) кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они
- 81. Водители ритма (пейсмейкерные клетки) ПРОВОДЯЩИЕ: - водители ритма (пейсмейкерные к-ки) – небольшого размера, мало гликогена, миофибриллы
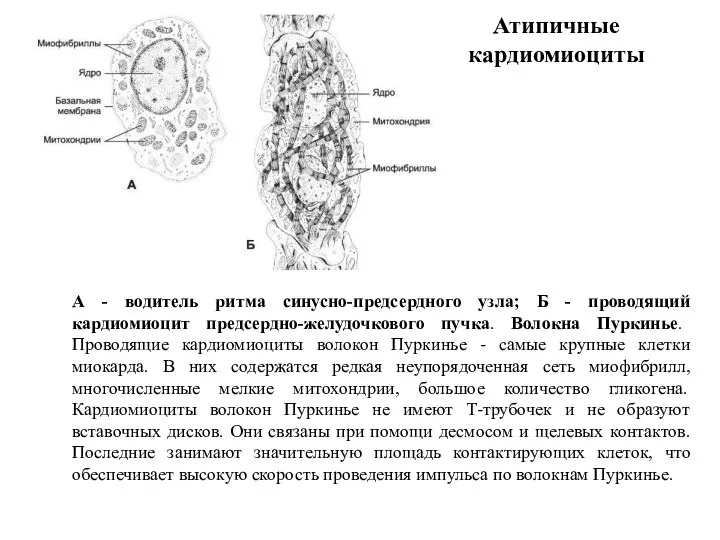
- 82. Атипичные кардиомиоциты А - водитель ритма синусно-предсердного узла; Б - проводящий кардиомиоцит предсердно-желудочкового пучка. Волокна Пуркинье.
- 83. ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА На деятельность сердца - сложной авторегуляторной и регулируемой системы - оказывает модулирующее влияние
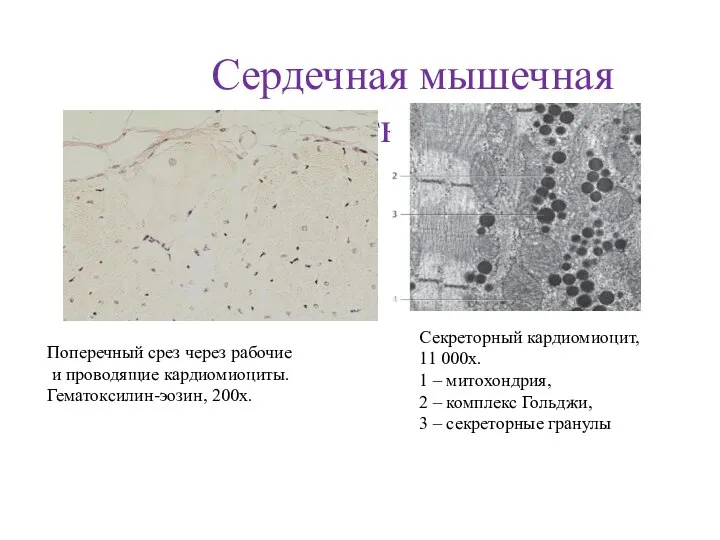
- 84. Сердечная мышечная ткань Поперечный срез через рабочие и проводящие кардиомиоциты. Гематоксилин-эозин, 200х. Секреторный кардиомиоцит, 11 000х.
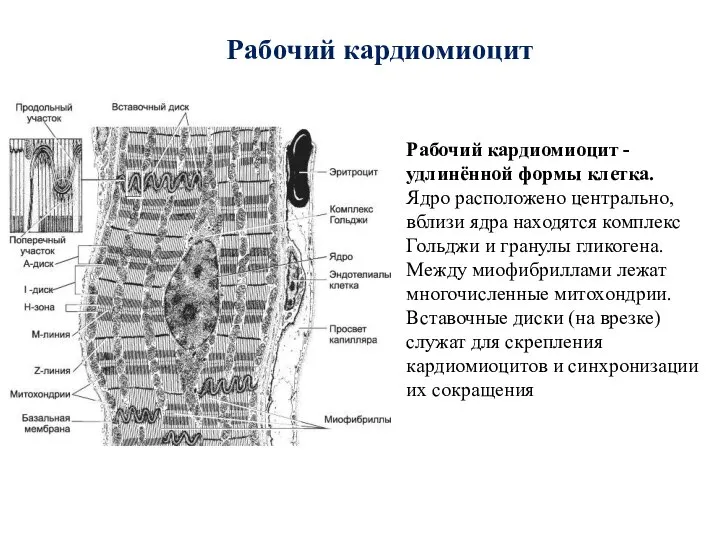
- 85. Рабочий кардиомиоцит Рабочий кардиомиоцит - удлинённой формы клетка. Ядро расположено центрально, вблизи ядра находятся комплекс Гольджи
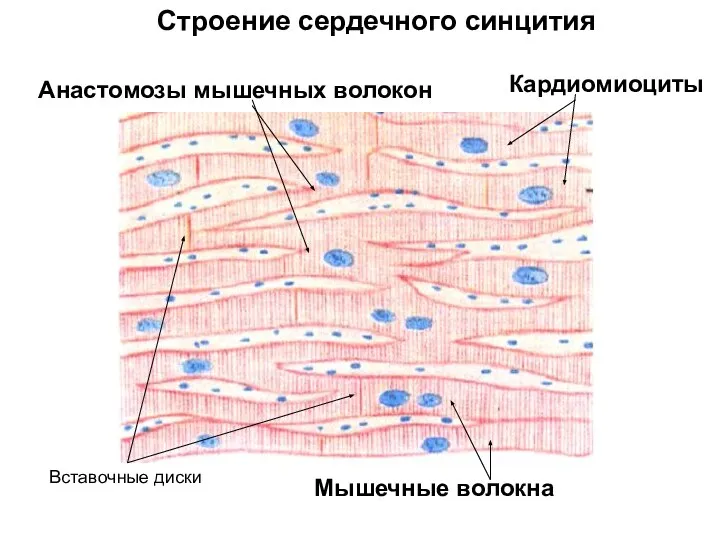
- 86. Анастомозы мышечных волокон Вставочные диски Строение сердечного синцития Кардиомиоциты Мышечные волокна
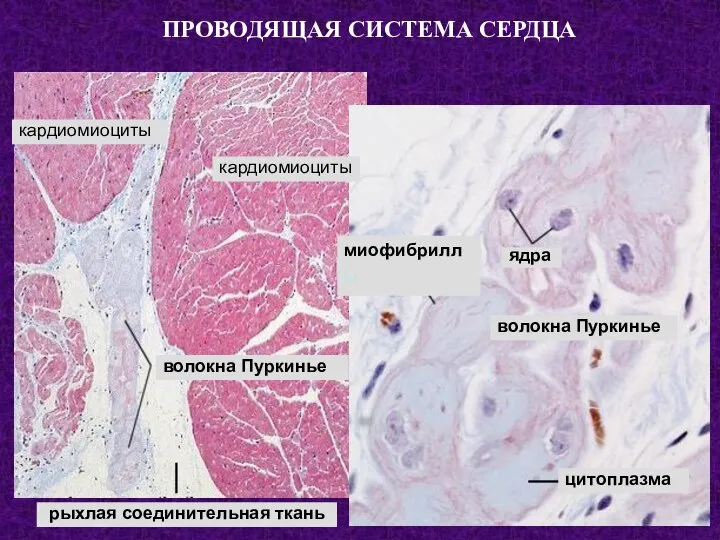
- 87. ПРОВОДЯЩАЯ СИСТЕМА СЕРДЦА кардиомиоциты кардиомиоциты волокна Пуркинье рыхлая соединительная ткань ядра волокна Пуркинье цитоплазма миофибриллы
- 89. Скачать презентацию
Слайд 2План лекции
Определение, происхождение и классификация мышечной ткани
Строение, функции, регенерация гладкой мышечной ткани
Строение,
План лекции
Определение, происхождение и классификация мышечной ткани
Строение, функции, регенерация гладкой мышечной ткани
Строение,

Слайд 3Мышечные ткани (МТ)
представляют собой группу тканей различного происхождения и строения, но объединенных
Мышечные ткани (МТ)
представляют собой группу тканей различного происхождения и строения, но объединенных

Слайд 4Общая морфофункциональная характеристика мышечной ткани (МТ)
Структурные элементы (клетки, волокна) обладают удлиненной формой;
Наличие
Общая морфофункциональная характеристика мышечной ткани (МТ)
Структурные элементы (клетки, волокна) обладают удлиненной формой;
Наличие

Слайд 5Классификация МТ
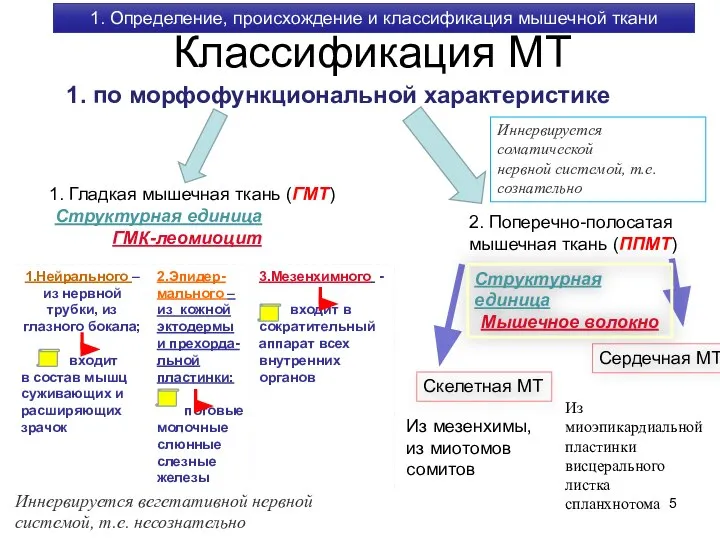
1. по морфофункциональной характеристике
1. Гладкая мышечная ткань (ГМТ)
2. Поперечно-полосатая
мышечная ткань
Классификация МТ
1. по морфофункциональной характеристике
1. Гладкая мышечная ткань (ГМТ)
2. Поперечно-полосатая
мышечная ткань
Слайд 6КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ТКАНЕЙ
2. Гистогенетическая классификация мышечных тканей
∙ Эктодермальные
− эпителиально-мышечная ткань (мышечные
КЛАССИФИКАЦИЯ МЫШЕЧНЫХ ТКАНЕЙ
2. Гистогенетическая классификация мышечных тканей
∙ Эктодермальные
− эпителиально-мышечная ткань (мышечные

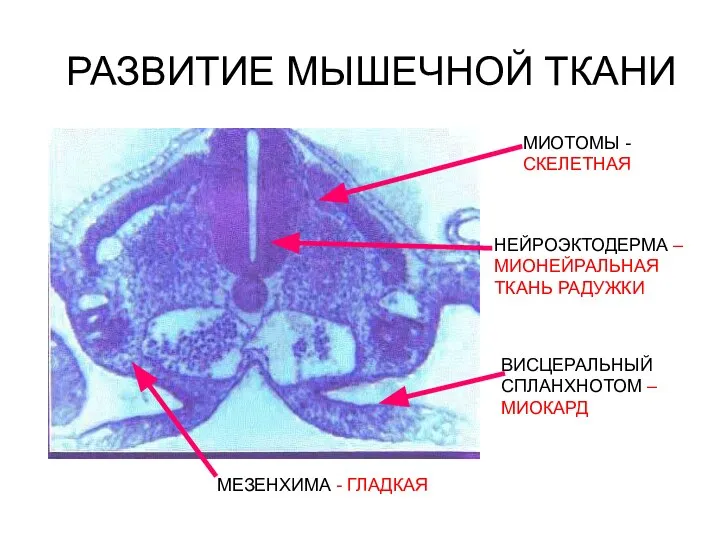
Слайд 7РАЗВИТИЕ МЫШЕЧНОЙ ТКАНИ
РАЗВИТИЕ МЫШЕЧНОЙ ТКАНИ
Слайд 8Классификация предложенная
Николаем Григорьевичем Хлопиным:
1. Гладкая МТ.
2. Поперечно-полосатая МТ.
1)Поперечно-полосатая МТ
Классификация предложенная
Николаем Григорьевичем Хлопиным:
1. Гладкая МТ.
2. Поперечно-полосатая МТ.
1)Поперечно-полосатая МТ

Слайд 9
Мышцы составляют большую массу тела и входят в состав многих органов.
ТЕРМИНОЛОГИЯ:
мышечная клетка
Мышцы составляют большую массу тела и входят в состав многих органов.
ТЕРМИНОЛОГИЯ:
мышечная клетка

Слайд 10Гладкая МТ входит в состав мышечных оболочек сосудов, кишечника, мочевыводящих, семявыводящих путей;
Гладкая МТ входит в состав мышечных оболочек сосудов, кишечника, мочевыводящих, семявыводящих путей;

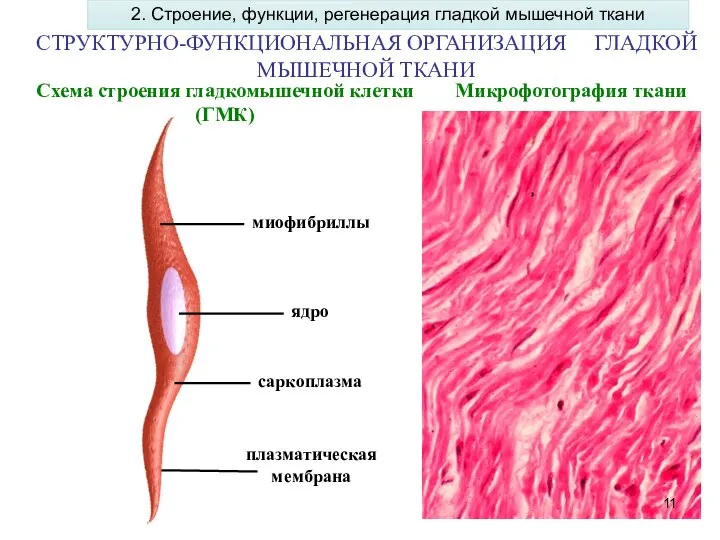
Слайд 11СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
Микрофотография ткани
ядро
миофибриллы
саркоплазма
плазматическая мембрана
Схема строения гладкомышечной клетки
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
Микрофотография ткани
ядро
миофибриллы
саркоплазма
плазматическая мембрана
Схема строения гладкомышечной клетки
Слайд 12Миоцит представляет собой чаще всего веретенообразную клетку (длиной 20—1000 мкм, диаметром 2—20
Миоцит представляет собой чаще всего веретенообразную клетку (длиной 20—1000 мкм, диаметром 2—20

Слайд 13Характеристика гадкого миоцита
9. Сарколемма ГМ окружена базальной мембраной, в которой вплетаются тонкие
Характеристика гадкого миоцита
9. Сарколемма ГМ окружена базальной мембраной, в которой вплетаются тонкие

Слайд 1411. Актиновые миофиламенты:
располагаются продольно или под углом
образуют трехмерную сеть
в месте их контакта
11. Актиновые миофиламенты:
располагаются продольно или под углом
образуют трехмерную сеть
в месте их контакта

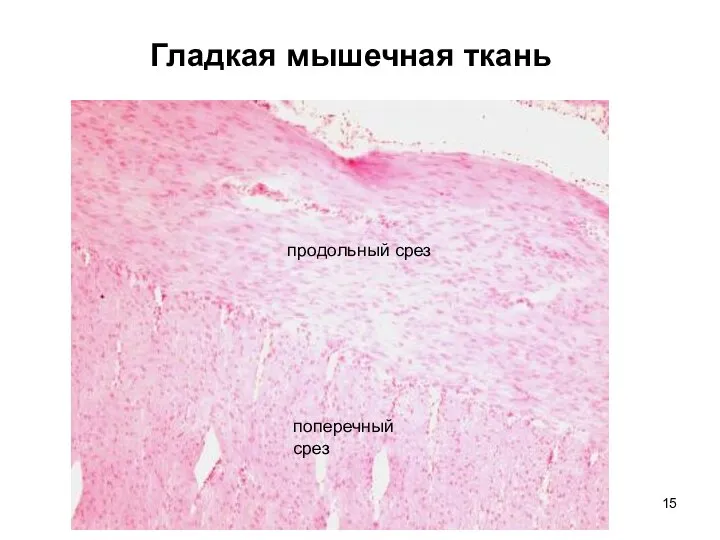
Слайд 15Гладкая мышечная ткань
продольный срез
поперечный
срез
Гладкая мышечная ткань
продольный срез
поперечный
срез
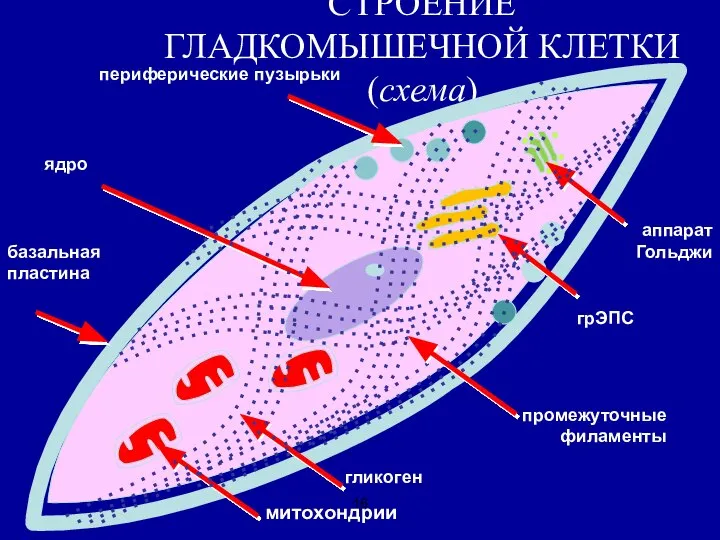
Слайд 16СТРОЕНИЕ ГЛАДКОМЫШЕЧНОЙ КЛЕТКИ (схема)
митохондрии
гликоген
ядро
грЭПС
аппарат
Гольджи
периферические пузырьки
базальная пластина
СТРОЕНИЕ ГЛАДКОМЫШЕЧНОЙ КЛЕТКИ (схема)
митохондрии
гликоген
ядро
грЭПС
аппарат
Гольджи
периферические пузырьки
базальная пластина
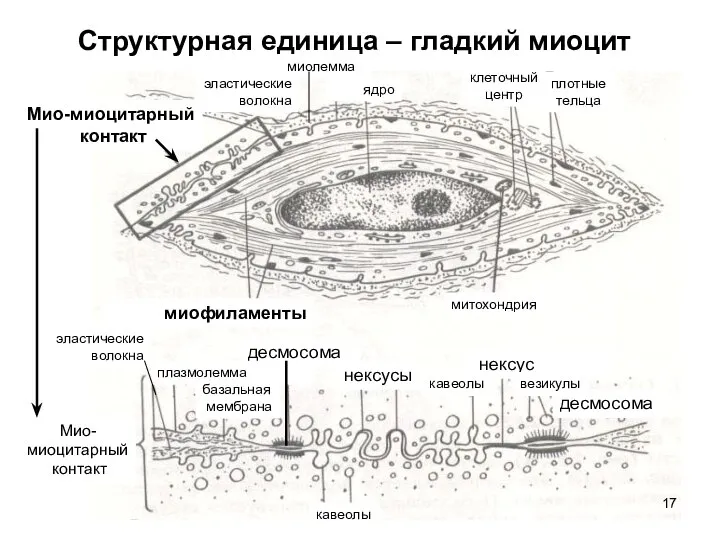
Слайд 17Структурная единица – гладкий миоцит
Мио-миоцитарный
контакт
миофиламенты
митохондрия
плотные
тельца
клеточный
центр
ядро
миолемма
эластические
волокна
эластические
волокна
плазмолемма
базальная
мембрана
десмосома
десмосома
кавеолы
кавеолы
нексусы
нексус
везикулы
Мио-
миоцитарный
контакт
Структурная единица – гладкий миоцит
Мио-миоцитарный
контакт
миофиламенты
митохондрия
плотные
тельца
клеточный
центр
ядро
миолемма
эластические
волокна
эластические
волокна
плазмолемма
базальная
мембрана
десмосома
десмосома
кавеолы
кавеолы
нексусы
нексус
везикулы
Мио-
миоцитарный
контакт
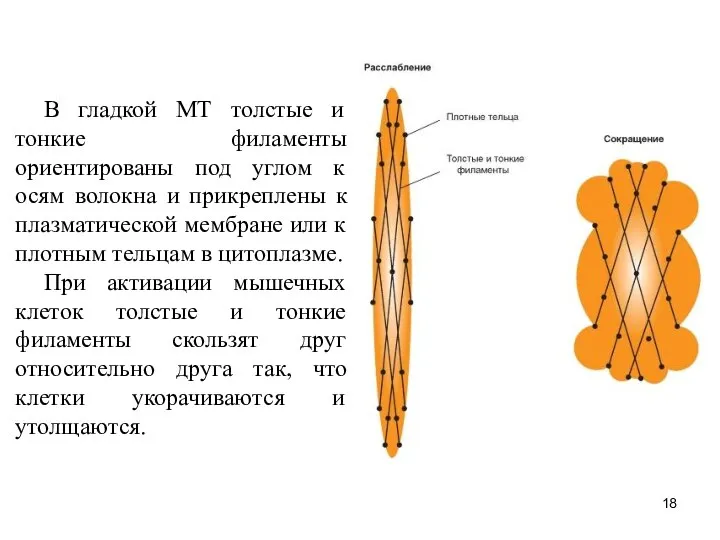
Слайд 18В гладкой МТ толстые и тонкие филаменты ориентированы под углом к осям
В гладкой МТ толстые и тонкие филаменты ориентированы под углом к осям
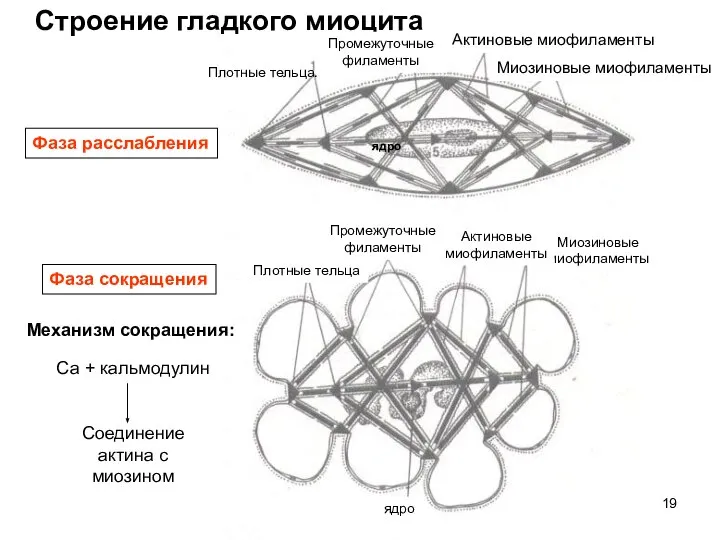
Слайд 19Строение гладкого миоцита
Фаза расслабления
Фаза сокращения
Плотные тельца:
Плотные тельца
Промежуточные
филаменты
Промежуточные
филаменты
ядро
ядро
Актиновые миофиламенты
Миозиновые
миофиламенты
Актиновые
миофиламенты
Миозиновые миофиламенты
Са +
Строение гладкого миоцита
Фаза расслабления
Фаза сокращения
Плотные тельца:
Плотные тельца
Промежуточные
филаменты
Промежуточные
филаменты
ядро
ядро
Актиновые миофиламенты
Миозиновые
миофиламенты
Актиновые
миофиламенты
Миозиновые миофиламенты
Са +
Слайд 20 Ядро в центре клетки, а периферическая часть клетки занята сократительным аппаратом: много
Ядро в центре клетки, а периферическая часть клетки занята сократительным аппаратом: много

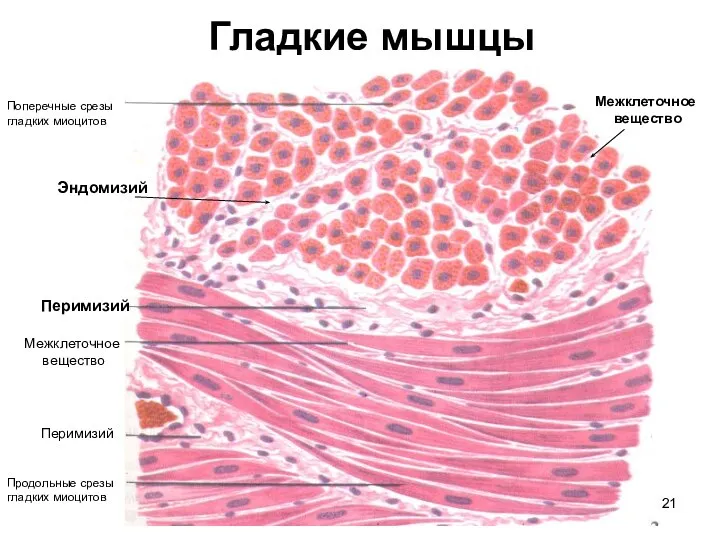
Слайд 21Гладкие мышцы
Поперечные срезы
гладких миоцитов
Продольные срезы
гладких миоцитов
Перимизий
Перимизий
Эндомизий
Межклеточное
вещество
Межклеточное
вещество
Гладкие мышцы
Поперечные срезы
гладких миоцитов
Продольные срезы
гладких миоцитов
Перимизий
Перимизий
Эндомизий
Межклеточное
вещество
Межклеточное
вещество
Слайд 22Гладкие мышечные ткани.
Нейральные – Миоциты расположены пучками, Эндомизий и Перимизий тонкие, содержат
Гладкие мышечные ткани.
Нейральные – Миоциты расположены пучками, Эндомизий и Перимизий тонкие, содержат

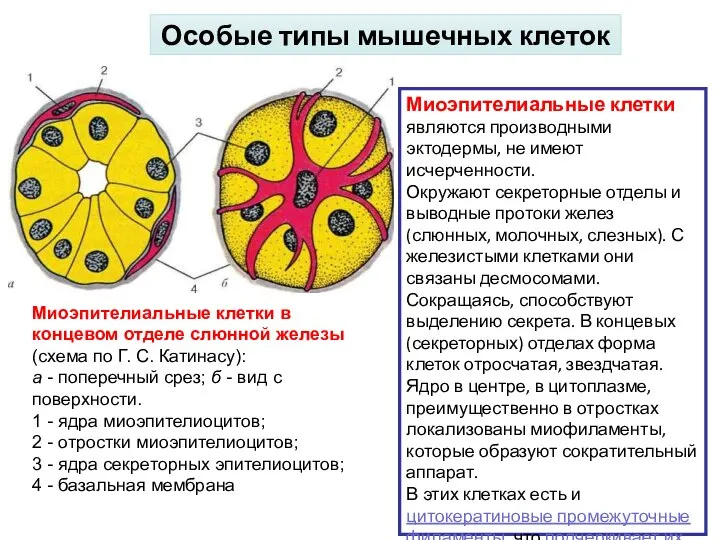
Слайд 23Особые типы мышечных клеток
Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности.
Окружают
Особые типы мышечных клеток
Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности.
Окружают
Слайд 24 Мионейральные клетки
Мионейральная ткань входит в состав мышц радужной оболочки глаза –
Мионейральные клетки
Мионейральная ткань входит в состав мышц радужной оболочки глаза –

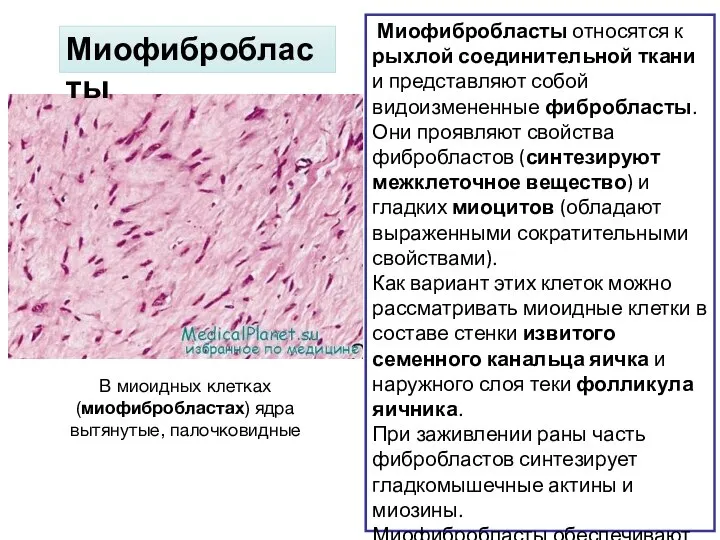
Слайд 25В миоидных клетках (миофибробластах) ядра вытянутые, палочковидные
Миофибробласты относятся к рыхлой соединительной
В миоидных клетках (миофибробластах) ядра вытянутые, палочковидные
Миофибробласты относятся к рыхлой соединительной
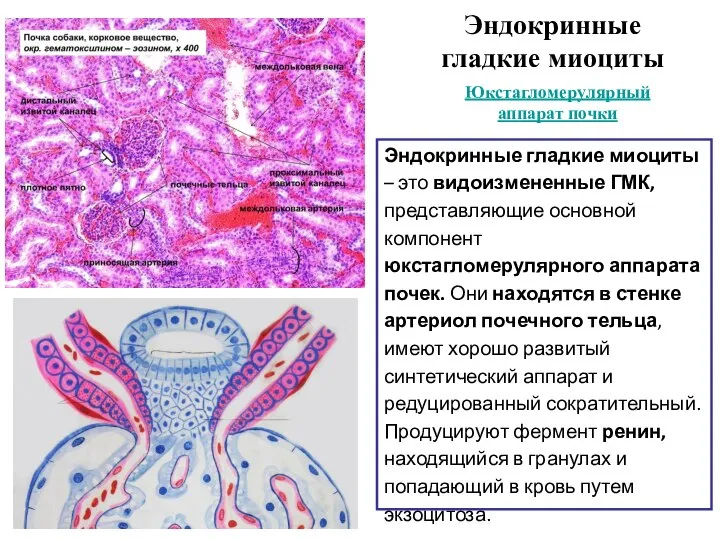
Слайд 26Эндокринные гладкие миоциты
Эндокринные гладкие миоциты – это видоизмененные ГМК, представляющие основной
Эндокринные гладкие миоциты
Эндокринные гладкие миоциты – это видоизмененные ГМК, представляющие основной
Слайд 27Регенерация гладкой мышечной ткани осуществляется несколькими способами:
посредством внутриклеточной регенерации гипертрофии при усилении
Регенерация гладкой мышечной ткани осуществляется несколькими способами:
посредством внутриклеточной регенерации гипертрофии при усилении

Слайд 283. Строение, функции, регенерация поперечно-полосатой
мышечной ткани (соматической)
3. Строение, функции, регенерация поперечно-полосатой
мышечной ткани (соматической)

Слайд 29Гистогенез
Источник происхождения в эмбриогенезе – это сомиты мезодермы (дорсо-медиальные участки, которые называются
Гистогенез
Источник происхождения в эмбриогенезе – это сомиты мезодермы (дорсо-медиальные участки, которые называются
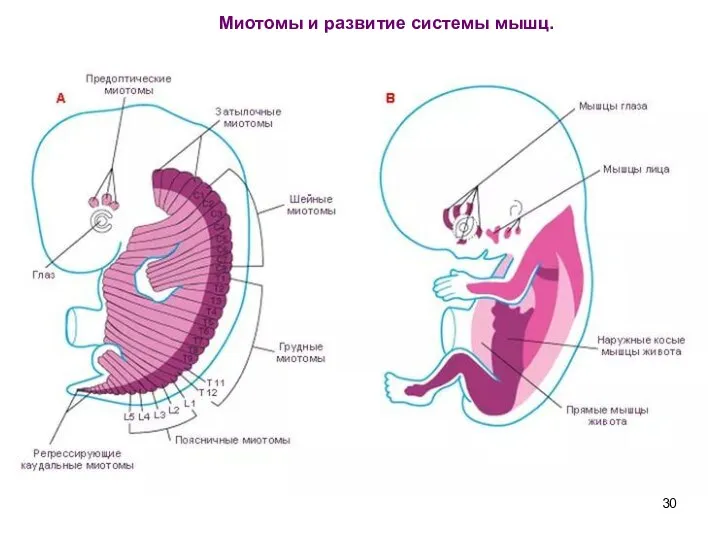
Слайд 30 Миотомы и развитие системы мышц.
Миотомы и развитие системы мышц.
Слайд 322 тип: Митотический миобласт переходит в состояние G1 – готовы всегда к
2 тип: Митотический миобласт переходит в состояние G1 – готовы всегда к

Слайд 33Скелетная мышечная ткань (соматического типа).
Основные гистологические элементы: скелетные мышечные волокна (функция сокращения)
Скелетная мышечная ткань (соматического типа).
Основные гистологические элементы: скелетные мышечные волокна (функция сокращения)

Слайд 34Строение скелетной мышечной ткани
Рассмотрим строение мышечной ткани на нескольких уровнях организации живого:
Строение скелетной мышечной ткани
Рассмотрим строение мышечной ткани на нескольких уровнях организации живого:

Слайд 351) Органный уровень: строение мышцы как органа.
Скелетная мышца состоит из пучков мышечных
1) Органный уровень: строение мышцы как органа.
Скелетная мышца состоит из пучков мышечных
Слайд 36СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Анатомия скелетной мускулатуры
сухожилие
мышца
эпимизиум
пучок мышечных
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Анатомия скелетной мускулатуры
сухожилие
мышца
эпимизиум
пучок мышечных
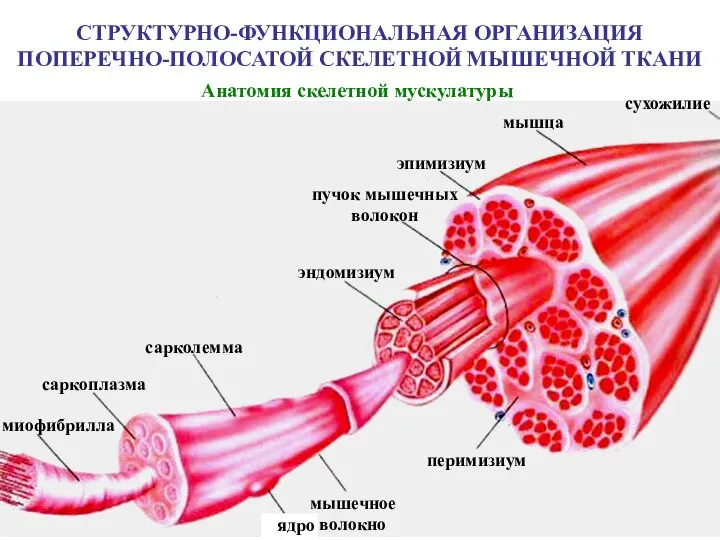
Слайд 37Структурно-функциональной единицей скелетной поперечнополосатой (исчерченной) мышечной ткани является мышечное волокно (симпласт) –
Структурно-функциональной единицей скелетной поперечнополосатой (исчерченной) мышечной ткани является мышечное волокно (симпласт) –

Слайд 38Клеточный уровень - строение мышечного волокна (миосимпласта).
Термин «мышечное волокно» подразумевает «миосимпласт», поскольку
Клеточный уровень - строение мышечного волокна (миосимпласта).
Термин «мышечное волокно» подразумевает «миосимпласт», поскольку
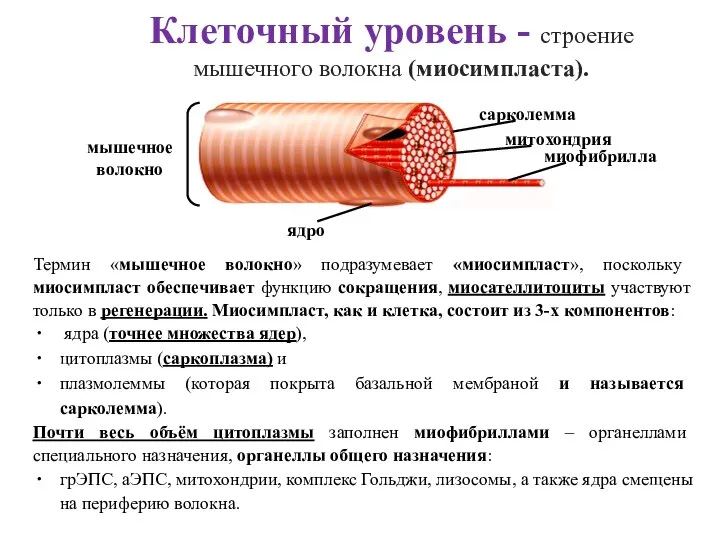
Слайд 39Субклеточный уровень: строение миофибриллы
I-диск содержит только тонкие нити.
В середине I-диска проходит
Субклеточный уровень: строение миофибриллы
I-диск содержит только тонкие нити.
В середине I-диска проходит

Слайд 40Субклеточный уровень: строение миофибриллы.
Саркомер содержит один А-диск (тёмный) и две половины I-диска
Субклеточный уровень: строение миофибриллы.
Саркомер содержит один А-диск (тёмный) и две половины I-диска
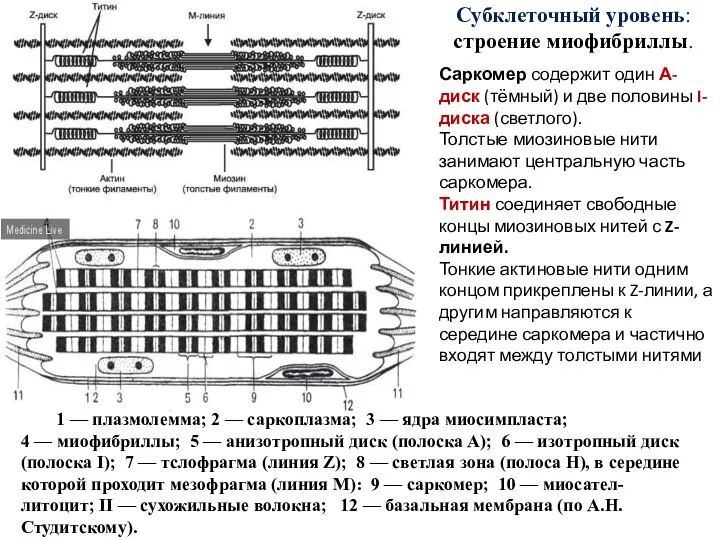
Слайд 41В мышечном волокне (миосимпласте) различают функциональные аппараты
1. Мембранный
2. Фибриллярный (сократительный) и
3. Трофический.
Трофический
В мышечном волокне (миосимпласте) различают функциональные аппараты
1. Мембранный
2. Фибриллярный (сократительный) и
3. Трофический.
Трофический

Слайд 42Каждая миофибрилла окружена регулярно повторяющимися элементами сaркоплазматического ретикулума - анастомозирующими мембранными трубочками
Каждая миофибрилла окружена регулярно повторяющимися элементами сaркоплазматического ретикулума - анастомозирующими мембранными трубочками
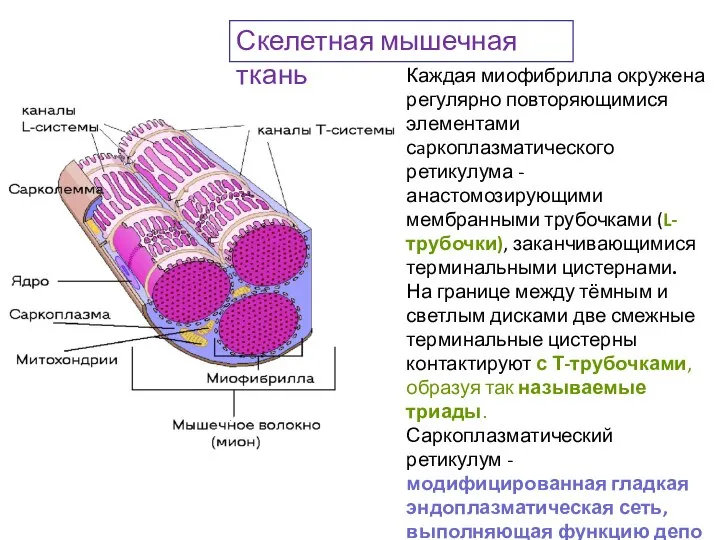
Слайд 43Молекулярный уровень: строение актиновых и миозиновых филаментов.
Под электронным микроскопом миофибриллы представляют агрегаты
Молекулярный уровень: строение актиновых и миозиновых филаментов.
Под электронным микроскопом миофибриллы представляют агрегаты

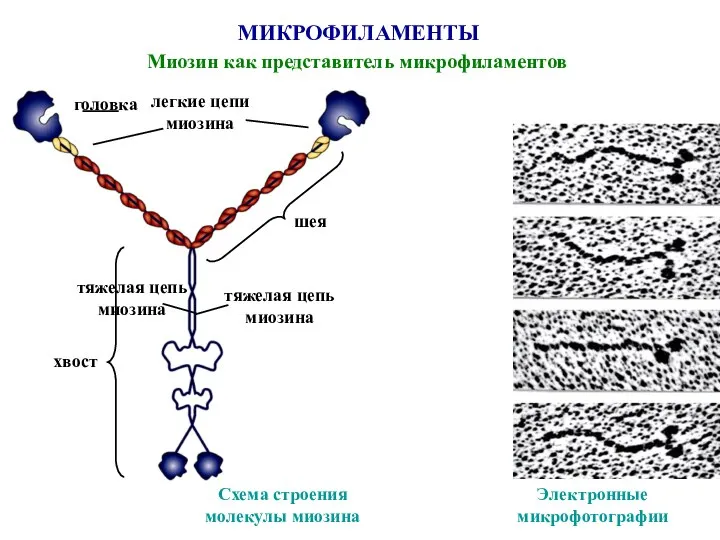
Слайд 44Тяжелые цепи состоят из шести полипептидных цепей, две из них тяжелые и
Тяжелые цепи состоят из шести полипептидных цепей, две из них тяжелые и

Слайд 45МИКРОФИЛАМЕНТЫ
Миозин как представитель микрофиламентов
тяжелая цепь
миозина
Схема строения
молекулы миозина
тяжелая цепь
миозина
легкие цепи
миозина
хвост
шея
головка
Электронные
микрофотографии
МИКРОФИЛАМЕНТЫ
Миозин как представитель микрофиламентов
тяжелая цепь
миозина
Схема строения
молекулы миозина
тяжелая цепь
миозина
легкие цепи
миозина
хвост
шея
головка
Электронные
микрофотографии
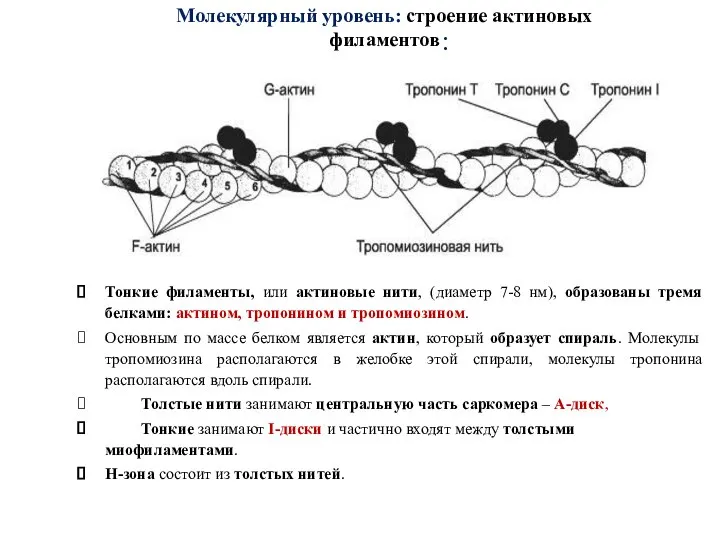
Слайд 46:
Тонкие филаменты, или актиновые нити, (диаметр 7-8 нм), образованы тремя белками: актином,
:
Тонкие филаменты, или актиновые нити, (диаметр 7-8 нм), образованы тремя белками: актином,
Слайд 47Молекулярный уровень: строение актиновых филаментов
В покое взаимодействие тонких и толстых нитей (миофиламентов)
Молекулярный уровень: строение актиновых филаментов
В покое взаимодействие тонких и толстых нитей (миофиламентов)

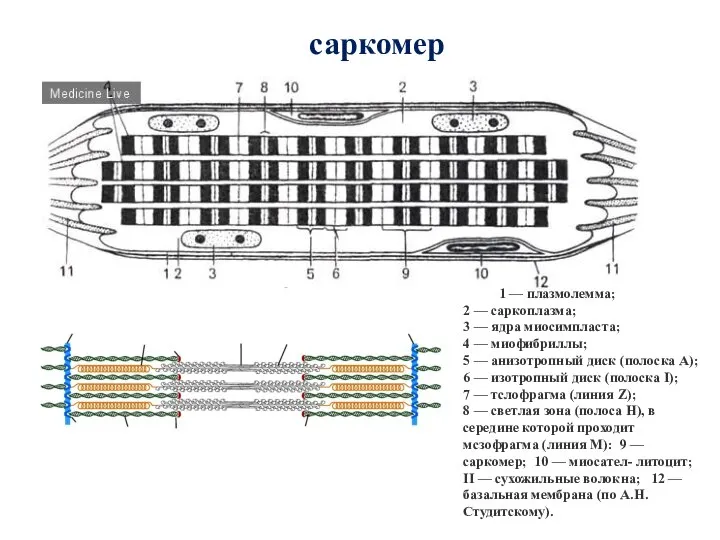
Слайд 48саркомер
1 — плазмолемма;
2 — саркоплазма;
3 — ядра миосимпласта;
4 — миофибриллы;
5 — анизотропный
саркомер
1 — плазмолемма; 2 — саркоплазма; 3 — ядра миосимпласта; 4 — миофибриллы; 5 — анизотропный
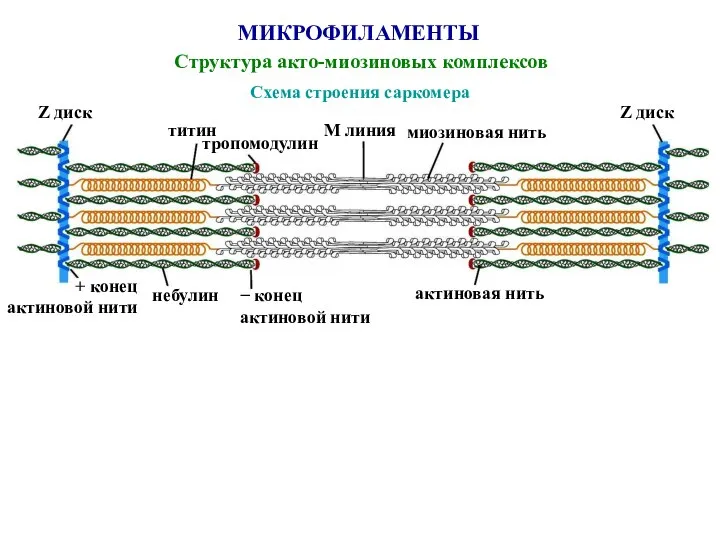
Слайд 49МИКРОФИЛАМЕНТЫ
Структура акто-миозиновых комплексов
Схема строения саркомера
Z диск
титин
тропомодулин
М линия
миозиновая нить
Z диск
+ конец
актиновой нити
небулин
− конец
актиновой
МИКРОФИЛАМЕНТЫ
Структура акто-миозиновых комплексов
Схема строения саркомера
Z диск
титин
тропомодулин
М линия
миозиновая нить
Z диск
+ конец
актиновой нити
небулин
− конец
актиновой
Слайд 50САРКОМЕР
Линия М
Миозин
Актин
Телофрагма
(линия Z)
Титин
Миозиновые
мостики
САРКОМЕР
Линия М
Миозин
Актин
Телофрагма
(линия Z)
Титин
Миозиновые
мостики
Слайд 51Молекулярный механизм мышечного сокращения
1. Прохождение нервного импульса через нервно-мышечный синапс и деполяризация
Молекулярный механизм мышечного сокращения
1. Прохождение нервного импульса через нервно-мышечный синапс и деполяризация
Слайд 52Сокращение мышцы
При сокращении мышечные волокна укорачиваются, но длина актиновых и миозиновых
Сокращение мышцы
При сокращении мышечные волокна укорачиваются, но длина актиновых и миозиновых
Слайд 53Структура мышечного волокна
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
потенциал действия
сарколемма
саркоплазматический ретикулум
Т-трубочка
Т-трубочка
саркомер
миофибриллы
Структура мышечного волокна
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
потенциал действия
сарколемма
саркоплазматический ретикулум
Т-трубочка
Т-трубочка
саркомер
миофибриллы
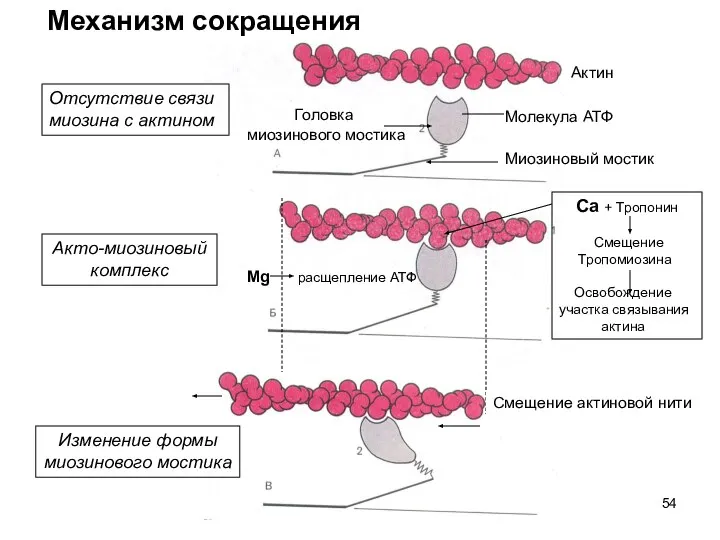
Слайд 54Актин
Механизм сокращения
Головка
миозинового мостика
Миозиновый мостик
Отсутствие связи
миозина с актином
Акто-миозиновый
комплекс
Изменение формы
миозинового мостика
Молекула
Актин
Механизм сокращения
Головка
миозинового мостика
Миозиновый мостик
Отсутствие связи
миозина с актином
Акто-миозиновый
комплекс
Изменение формы
миозинового мостика
Молекула
Слайд 55Структурные элементы саркомера (S)
в расслабленном состоянии можно выразить формулой:
S = Z1 +1/2I1
Структурные элементы саркомера (S)
в расслабленном состоянии можно выразить формулой:
S = Z1 +1/2I1

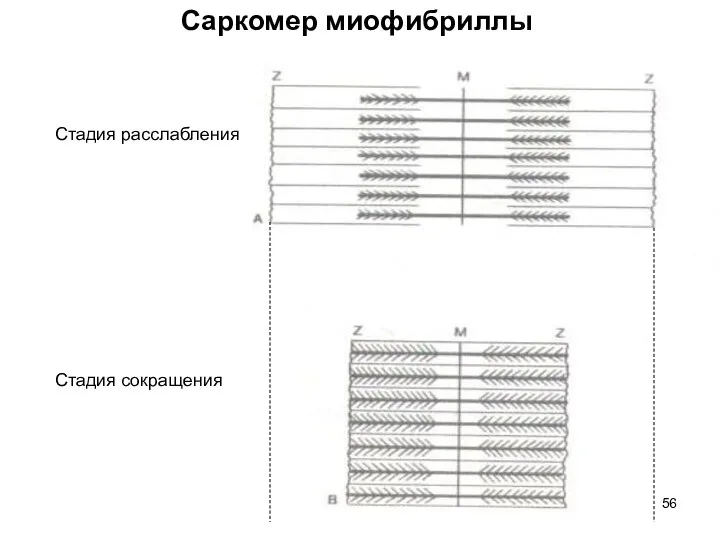
Слайд 56Стадия расслабления
Стадия сокращения
Саркомер миофибриллы
Стадия расслабления
Стадия сокращения
Саркомер миофибриллы
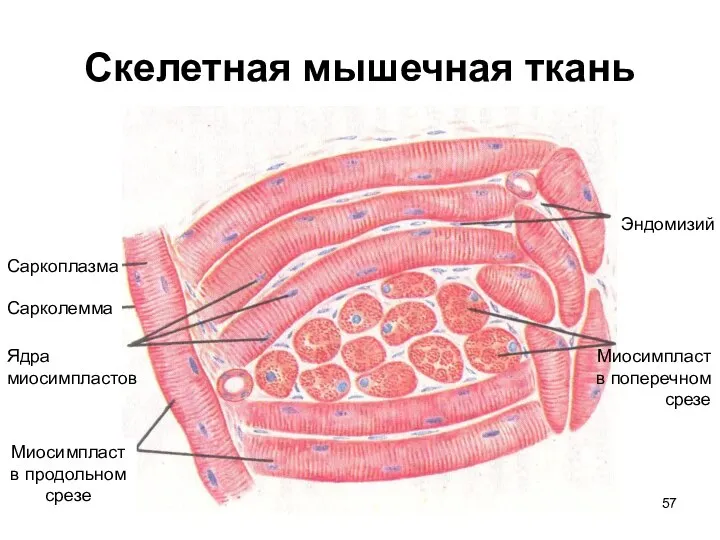
Слайд 57Скелетная мышечная ткань
Эндомизий
Миосимпласт
в поперечном
срезе
Миосимпласт
в продольном
срезе
Саркоплазма
Сарколемма
Ядра
миосимпластов
Скелетная мышечная ткань
Эндомизий
Миосимпласт
в поперечном
срезе
Миосимпласт
в продольном
срезе
Саркоплазма
Сарколемма
Ядра
миосимпластов
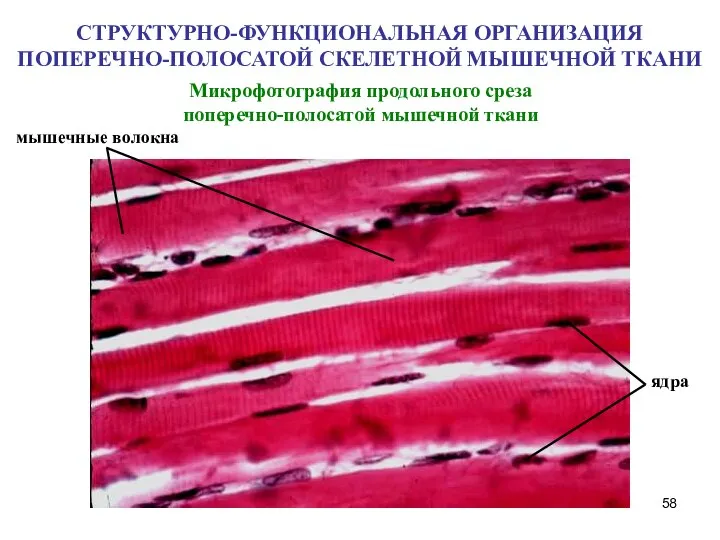
Слайд 58Микрофотография продольного среза поперечно-полосатой мышечной ткани
мышечные волокна
ядра
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Микрофотография продольного среза поперечно-полосатой мышечной ткани
мышечные волокна
ядра
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Слайд 59Скелетная мышечная ткань
Скелетная мышечная ткань

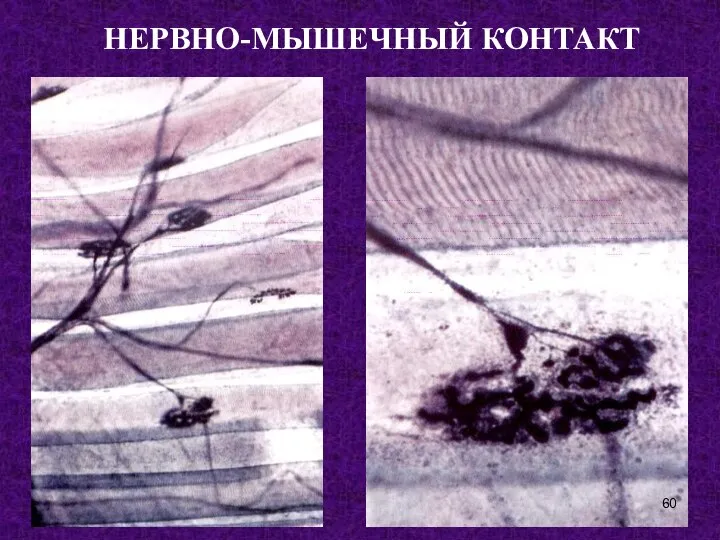
Слайд 60НЕРВНО-МЫШЕЧНЫЙ КОНТАКТ
НЕРВНО-МЫШЕЧНЫЙ КОНТАКТ
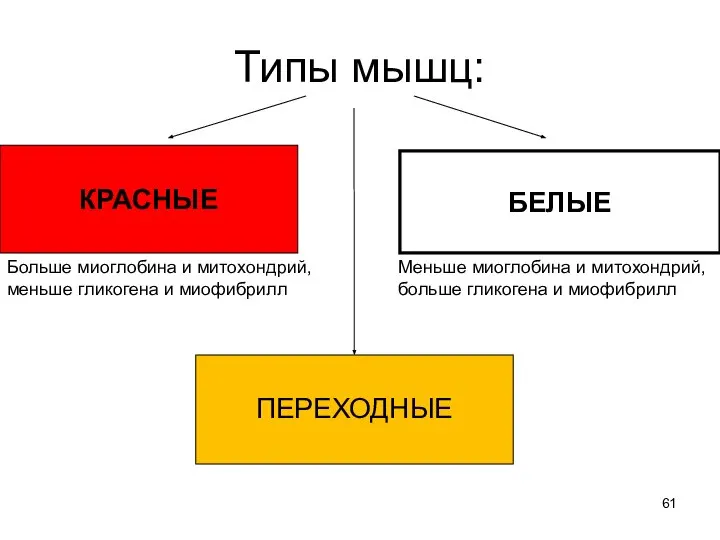
Слайд 61Типы мышц:
КРАСНЫЕ
БЕЛЫЕ
ПЕРЕХОДНЫЕ
Больше миоглобина и митохондрий,
меньше гликогена и миофибрилл
Меньше миоглобина и митохондрий,
больше гликогена
Типы мышц:
КРАСНЫЕ
БЕЛЫЕ
ПЕРЕХОДНЫЕ
Больше миоглобина и митохондрий,
меньше гликогена и миофибрилл
Меньше миоглобина и митохондрий,
больше гликогена
Слайд 62ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
Красные (окислительные) мышечные волокна небольшого диаметра, окружены массой капилляров, содержат
ТИПЫ МЫШЕЧНЫХ ВОЛОКОН
Красные (окислительные) мышечные волокна небольшого диаметра, окружены массой капилляров, содержат

Слайд 63Красные и белые мионы
Различия белых и красных мионов
Красные и белые мионы
Различия белых и красных мионов

Слайд 64ИННЕРВАЦИЯ:
Скелетные мышцы получают
двигательную,
чувствительную и
трофическую (вегетативную) иннервацию.
Двигательную (эфферентную)
ИННЕРВАЦИЯ:
Скелетные мышцы получают
двигательную,
чувствительную и
трофическую (вегетативную) иннервацию.
Двигательную (эфферентную)

Слайд 65КРОВОСНАБЖЕНИЕ
Скелетные мышцы богато снабжаются кровью. В рыхлой соединительной ткани перимизия в большом
КРОВОСНАБЖЕНИЕ
Скелетные мышцы богато снабжаются кровью. В рыхлой соединительной ткани перимизия в большом

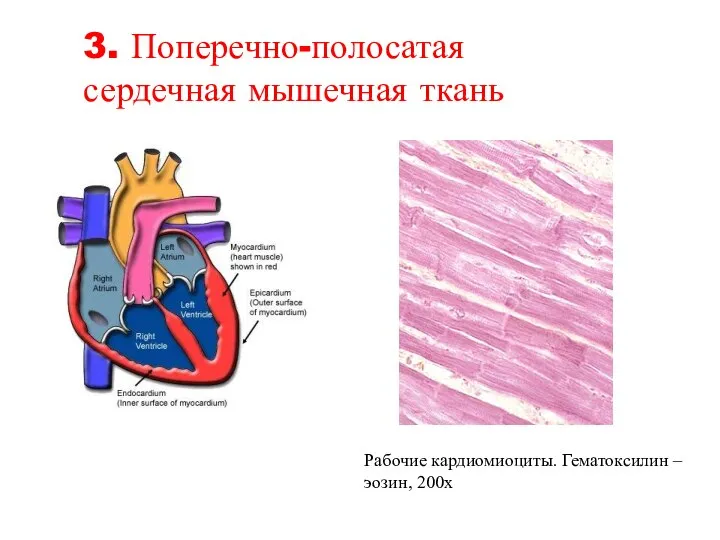
Слайд 66Рабочие кардиомиоциты. Гематоксилин – эозин, 200х
3. Поперечно-полосатая
сердечная мышечная ткань
Рабочие кардиомиоциты. Гематоксилин – эозин, 200х
3. Поперечно-полосатая
сердечная мышечная ткань
Слайд 67Поперечнополосатые мышечные ткани
сердечного типа
2. Сердечная мышечная ткань (целомического типа)
Целомический (сердечный)----Висцеральный листок спланхнотома
Поперечнополосатые мышечные ткани
сердечного типа
2. Сердечная мышечная ткань (целомического типа)
Целомический (сердечный)----Висцеральный листок спланхнотома

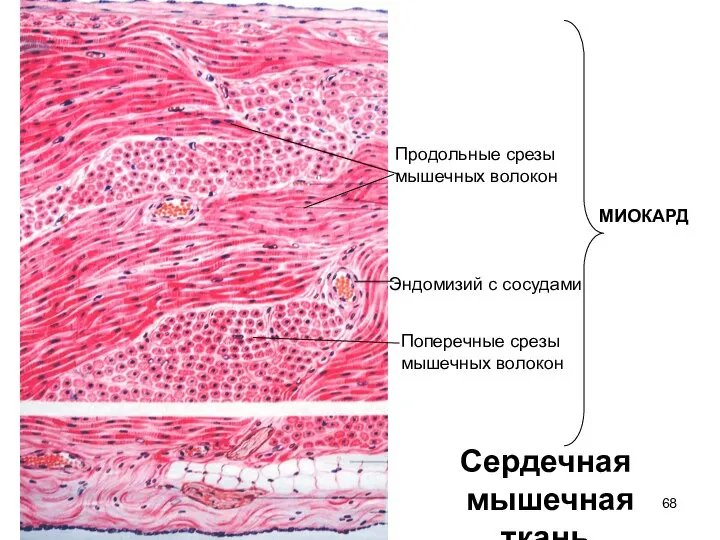
Слайд 68Сердечная
мышечная ткань
Продольные срезы
мышечных волокон
Поперечные срезы
мышечных волокон
Эндомизий с сосудами
МИОКАРД
Сердечная
мышечная ткань
Продольные срезы
мышечных волокон
Поперечные срезы
мышечных волокон
Эндомизий с сосудами
МИОКАРД
Слайд 69СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ.
Встречается в стенке сердца, проксимальной части аорты, в верхней полой
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ.
Встречается в стенке сердца, проксимальной части аорты, в верхней полой

Слайд 70КАРДИОМИОЦИТЫ
Клетки расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры
КАРДИОМИОЦИТЫ
Клетки расположены между элементами рыхлой волокнистой соединительной ткани, содержащей многочисленные кровеносные капилляры

Слайд 71Сердечная мышечная ткань
Электронная микрофотография
рабочего кардиомиоцита, 2500х.
1 – ядро, 2 –
Сердечная мышечная ткань
Электронная микрофотография
рабочего кардиомиоцита, 2500х.
1 – ядро, 2 –
Слайд 72СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Схема строения Микрофотография
ядро
вставочный диск
кардиомиоцит
анастамоз
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПОПЕРЕЧНО-ПОЛОСАТОЙ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Схема строения Микрофотография
ядро
вставочный диск
кардиомиоцит
анастамоз
Слайд 73Типичные кардиомиоциты (рабочие, сократительные)
Типичные (рабочие, сократительные) кардиомиоциты – клетки цилиндрической формы, длиной до
Типичные кардиомиоциты (рабочие, сократительные)
Типичные (рабочие, сократительные) кардиомиоциты – клетки цилиндрической формы, длиной до

Слайд 74Вставочный диск
базальная
мембрана
Десмосома
Нексус
митохондрии
миофибриллы
миофибриллы
саркоплазма
Диск И
Диск А
Интердигитации
Вставочный диск
базальная
мембрана
Десмосома
Нексус
митохондрии
миофибриллы
миофибриллы
саркоплазма
Диск И
Диск А
Интердигитации
Слайд 75Мембранный аппарат
Мембранный аппарат: каждая клетка покрыта оболочкой, состоящей из комплекса плазмолеммы и
Мембранный аппарат
Мембранный аппарат: каждая клетка покрыта оболочкой, состоящей из комплекса плазмолеммы и

Слайд 76Фибриллярный (сократительный) аппарат.
Фибриллярный (сократительный) аппарат.
Большую часть цитоплазмы кардиомиоцита занимают органеллы специального назначения
Фибриллярный (сократительный) аппарат.
Фибриллярный (сократительный) аппарат.
Большую часть цитоплазмы кардиомиоцита занимают органеллы специального назначения

Слайд 77Сократительные кардиомиоциты
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна,
Сократительные кардиомиоциты
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна,

Слайд 78Предсердные и желудочковые кардиомиоциты
Предсердные и желудочковые кардиомиоциты относятся к разным популяциям рабочих
Предсердные и желудочковые кардиомиоциты
Предсердные и желудочковые кардиомиоциты относятся к разным популяциям рабочих

Слайд 79Секреторные кардиомиоциты
Секреторные кардиомиоциты выполняют особую функцию. Все кардиомиоциты покрыты базальной мембраной. В
Секреторные кардиомиоциты
Секреторные кардиомиоциты выполняют особую функцию. Все кардиомиоциты покрыты базальной мембраной. В

Слайд 80Проводящие (атипичные) кардиомиоциты
Проводящие (атипичные) кардиомиоциты обладают способностью к генерации и быстрому проведению электрических
Проводящие (атипичные) кардиомиоциты
Проводящие (атипичные) кардиомиоциты обладают способностью к генерации и быстрому проведению электрических

Слайд 81Водители ритма (пейсмейкерные клетки)
ПРОВОДЯЩИЕ: - водители ритма (пейсмейкерные к-ки) – небольшого размера,
Водители ритма (пейсмейкерные клетки)
ПРОВОДЯЩИЕ: - водители ритма (пейсмейкерные к-ки) – небольшого размера,

Слайд 82Атипичные кардиомиоциты
А - водитель ритма синусно-предсердного узла; Б - проводящий кардиомиоцит предсердно-желудочкового
Атипичные кардиомиоциты
А - водитель ритма синусно-предсердного узла; Б - проводящий кардиомиоцит предсердно-желудочкового
Слайд 83ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА
На деятельность сердца - сложной авторегуляторной и регулируемой системы -
ДВИГАТЕЛЬНАЯ ИННЕРВАЦИЯ СЕРДЦА
На деятельность сердца - сложной авторегуляторной и регулируемой системы -

Слайд 84Сердечная мышечная ткань
Поперечный срез через рабочие
и проводящие кардиомиоциты. Гематоксилин-эозин, 200х.
Секреторный кардиомиоцит,
Сердечная мышечная ткань
Поперечный срез через рабочие
и проводящие кардиомиоциты. Гематоксилин-эозин, 200х.
Секреторный кардиомиоцит,
Слайд 85Рабочий кардиомиоцит
Рабочий кардиомиоцит - удлинённой формы клетка. Ядро расположено центрально, вблизи ядра
Рабочий кардиомиоцит
Рабочий кардиомиоцит - удлинённой формы клетка. Ядро расположено центрально, вблизи ядра
Слайд 86Анастомозы мышечных волокон
Вставочные диски
Строение сердечного синцития
Кардиомиоциты
Мышечные волокна
Анастомозы мышечных волокон
Вставочные диски
Строение сердечного синцития
Кардиомиоциты
Мышечные волокна
Слайд 87ПРОВОДЯЩАЯ СИСТЕМА СЕРДЦА
кардиомиоциты
кардиомиоциты
волокна Пуркинье
рыхлая соединительная ткань
ядра
волокна Пуркинье
цитоплазма
миофибриллы
ПРОВОДЯЩАЯ СИСТЕМА СЕРДЦА
кардиомиоциты
кардиомиоциты
волокна Пуркинье
рыхлая соединительная ткань
ядра
волокна Пуркинье
цитоплазма
миофибриллы
 Соловей – великий маэстро
Соловей – великий маэстро Разнообразие внешнего строения бактериальных клеток
Разнообразие внешнего строения бактериальных клеток Выполнила Ученица 9 а класса Школы № 953 Васильева Маргарита Научный руководитель: Гусева Наталья Геннадьевна
Выполнила Ученица 9 а класса Школы № 953 Васильева Маргарита Научный руководитель: Гусева Наталья Геннадьевна Ядовитые животные и растения
Ядовитые животные и растения Видоизменения побегов. Лабораторная работа
Видоизменения побегов. Лабораторная работа Обґрунтування параметрів дозуючого пристрою безпілотного літального апарату для біологічного захисту рослин
Обґрунтування параметрів дозуючого пристрою безпілотного літального апарату для біологічного захисту рослин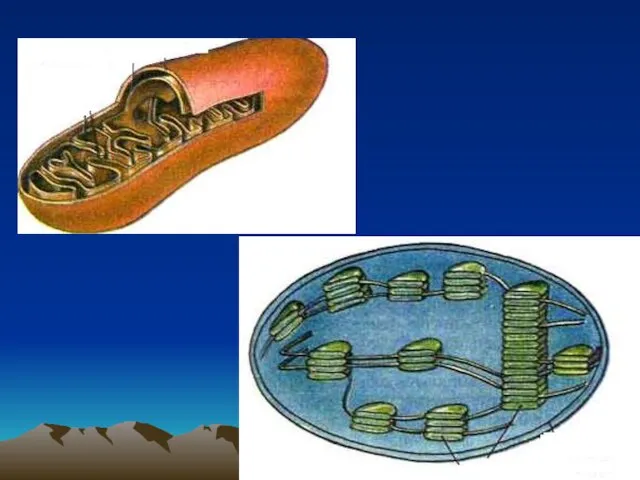 Строение прокариот. Метаболизм
Строение прокариот. Метаболизм Витамины и их значение для организма
Витамины и их значение для организма Дыхательная система
Дыхательная система Презентация на тему ОДНОКЛЕТОЧНЫЕ ОРГАНИЗМЫ
Презентация на тему ОДНОКЛЕТОЧНЫЕ ОРГАНИЗМЫ  Эволюция человека и обезьян шла параллельно друг другу, а не последовательно
Эволюция человека и обезьян шла параллельно друг другу, а не последовательно Про зрение
Про зрение Суставы. Практика
Суставы. Практика Органоиды, участвующие в энергообеспечении клетки
Органоиды, участвующие в энергообеспечении клетки chasti_rasteniy
chasti_rasteniy Яблоко. Самый полезный и любимый мой фрукт
Яблоко. Самый полезный и любимый мой фрукт Плесневые грибы и дрожжи
Плесневые грибы и дрожжи Грибы (фотографии)
Грибы (фотографии) Корнеплоды и клубнеплоды
Корнеплоды и клубнеплоды Кошки. Виды кошек
Кошки. Виды кошек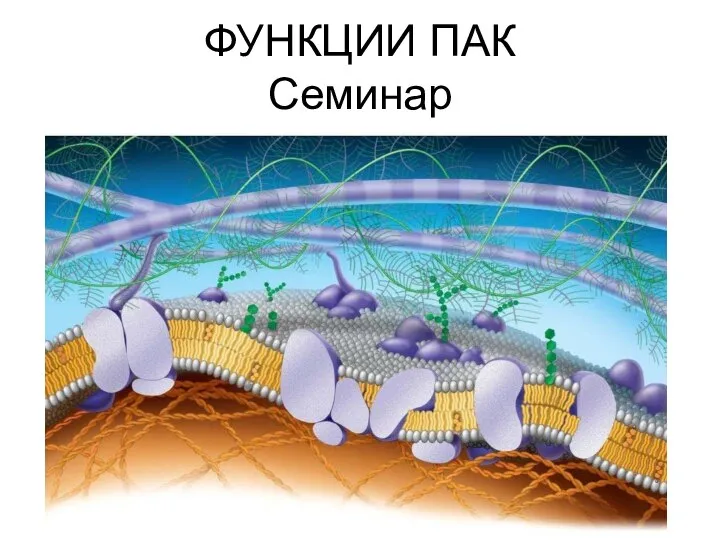 Функции_ПАК_контактн,_рецепт_и_др
Функции_ПАК_контактн,_рецепт_и_др Приручение животных
Приручение животных Силы в природе
Силы в природе Поток генов. Миграция
Поток генов. Миграция Оводы
Оводы Дикие и домашние животные
Дикие и домашние животные Классифиция живых организмов
Классифиция живых организмов Амурский тигр на грани исчезновения
Амурский тигр на грани исчезновения