- 基因表达调控

Содержание
- 2. 第 一 节 基本概念与原理 Basic Conceptions and Principle
- 3. 一、基因表达的概念 * 基因组(genome) 一个细胞或病毒所携带的全部遗传信息或整套基因。 基因经过转录、翻译,产生具有特异生物学功能的蛋白质分子的过程。 * 基因表达(gene expression) 基因表达是受调控的
- 4. 二、基因表达的时间性及空间性 (一)时间特异性
- 5. (二)空间特异性 基因表达伴随时间顺序所表现出的这种分布差异,实际上是由细胞在器官的分布决定的,所以空间特异性又称细胞或组织特异性(cell or tissue specificity)。 在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,称之为基因表达的空间特异性(spatial specificity)。
- 6. 三、基因表达的方式 按对刺激的反应性,基因表达的方式分为: (一)组成性表达 某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因(housekeeping gene)。
- 7. 无论表达水平高低,管家基因较少受环境因素影响,而是在个体各个生长阶段的大多数或几乎全部组织中持续表达,或变化很小。区别于其他基因,这类基因表达被视为组成性基因表达(constitutive gene expression)。
- 8. (二)诱导和阻遏表达 在特定环境信号刺激下,相应的基因被激活,基因表达产物增加,这种基因称为可诱导基因。 可诱导基因在特定环境中表达增强的过程,称为诱导(induction)。 如果基因对环境信号应答是被抑制,这种基因是可阻遏基因。可阻遏基因表达产物水平降低的过程称为阻遏(repression)。
- 9. 在一定机制控制下,功能上相关的一组基因,无论其为何种表达方式,均需协调一致、共同表达,即为协调表达(coordinate expression),这种调节称为协调调节(coordinate regulation)。
- 10. 四、基因表达调控的生物学意义 (一)适应环境、维持生长和增殖 (二)维持个体发育与分化
- 11. 五、基因表达调控的基本原理 (一)基因表达的多级调控 基因激活 转录起始 转录后加工 mRNA降解 转录起始
- 12. (二)基因转录激活调节基本要素 基因表达的调节与基因的结构、性质,生物个体或细胞所处的内、外环境,以及细胞内所存在的转录调节蛋白有关。 1. 特异DNA序列和调节蛋白质
- 13. 原核生物 —— 操纵子(operon) 机制
- 15. 共有序列(consensus sequence) 决定启动序列的转录活性大小。 某些特异因子(蛋白质)决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力。
- 16. 2) 操纵序列 ——阻遏蛋白(repressor)的结合位点 当操纵序列结合有阻遏蛋白时,会阻碍RNA聚合酶与启动序列的结合,或是RNA聚合酶不能沿DNA向前移动 ,阻碍转录。
- 17. 3) 其他调节序列、调节蛋白 例如 激活蛋白(activator)可结合启动序列邻近的DNA序列,促进RNA聚合酶与启动序列的结合,增强RNA聚合酶活性。 有些基因在没有激活蛋白存在时,RNA聚合酶很少或完全不能结合启动序列。
- 18. 真核生物 不同真核生物的顺式作用元件中也会发现一些共有序列 ,如TATA盒、CAAT盒等,这些共有序列是RNA聚合酶或特异转录因子的结合位点。
- 19. 2) 真核基因的调节蛋白 还有蛋白质因子可特异识别、结合自身基因的调节序列,调节自身基因的表达,称顺式作用。 由某一基因表达产生的蛋白质因子,通过与另一基因的特异的顺式作用元件相互作用,调节其表达。 反式作用因子(trans-acting factor) 这种调节作用称为反式作用。
- 20. 反式调节 顺式调节
- 21. 指的是反式作用因子与顺式作用元件之间的特异识别及结合。通常是非共价结合,被识别的DNA结合位点通常呈对称、或不完全对称结构。 绝大多数调节蛋白质结合DNA前,需通过蛋白质-蛋白质相互作用,形成二聚体(dimer)或多聚体(polymer)。
- 22. 3. RNA聚合酶 ⑴ 原核启动序列/真核启动子与RNA聚合酶活性 RNA聚合酶与其的亲和力,影响转录。 ⑵ 调节蛋白与RNA聚合酶活性 一些特异调节蛋白在适当环境信号刺激下表达,然后通过DNA-蛋白质、蛋白质-蛋白质相互作用影响RNA聚合酶活性。
- 23. 第 二 节 原核基因转录调节 Regulation of Prokaryotic Gene Transcription
- 24. (一)σ因子决定RNA聚合酶识别特异性 (二)操纵子模型的普遍性 (三)阻遏蛋白与阻遏机制的普遍性
- 25. 二、乳糖操纵子调节机制 (一)乳糖操纵子(lac operon)的结构 调控区 CAP结合位点
- 26. 没有乳糖存在时 (二)阻遏蛋白的负性调节
- 27. 有乳糖存在时
- 28. 无葡萄糖,cAMP浓度高时 有葡萄糖,cAMP浓度低时 (三)CAP的正性调节
- 29. (四)协调调节 ※当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用; ※如无CAP存在,即使没有阻遏蛋白与操纵序列结合,操纵子仍无转录活性。 单纯乳糖存在时,细菌利用乳糖作碳源; 若有葡萄糖或葡萄糖/乳糖共同存在时,细菌首先利用葡萄糖。 葡萄糖对 lac 操纵子的阻遏作用称分解代谢阻遏(catabolic repression)。
- 30. 低半乳糖时 高半乳糖时 葡萄糖低 cAMP浓度高 葡萄糖高cAMP浓度低
- 31. Trp 高时 Trp 低时 mRNA O P trpR 调节区 结构基因 RNA聚合酶 RNA聚合酶 ? 三、其他转录调节机制 (一)转录衰减 色氨酸操纵子
- 32. 前导序列 第10、11密码子为trp密码子 14aa前导肽编码区: 包含序列1 形成发夹结构能力强弱: 序列1/2>序列2/3>序列3/4
- 33. UUUU 3’ 前导肽 前导mRNA 1.当色氨酸浓度高时 转录衰减机制 衰减子结构 就是终止子 可使转录 RNA聚合酶 终止
- 34. 前导肽 前导mRNA RNA聚合酶 2.当色氨酸浓度低时 Trp合成酶系相关 结构基因被转录 序列3、4不能形成衰减子结构
- 35. (二)基因重组 沙门菌鞭毛素基因的调节
- 36. (三)SOS反应 紫外线 与DNA 损伤修复有关的酶和蛋白质 Lex A阻遏蛋白 DNA
- 37. 第 三 节 真核基因转录调节 Regulation of Eukaryotic Gene Transcription
- 38. 一、真核基因组结构特点 (一)真核基因组结构庞大
- 39. (二)单顺反子 单顺反子(monocistron) 即一个编码基因转录生成一个mRNA分子,经翻译生成一条多肽链。 (三)重复序列 (四)基因不连续性
- 40. 二、真核基因表达调控特点 (一)RNA聚合酶 (二)活性染色体结构变化 1. 对核酸酶敏感 活化基因常有超敏位点,位于调节蛋白结合位点附近。
- 41. 2. DNA拓扑结构变化 天然双链DNA均以负性超螺旋构象存在; 基因活化后 3. DNA碱基修饰变化
- 42. 4. 组蛋白变化 ① 富含Lys组蛋白水平降低 ② H2A, H2B二聚体不稳定性增加 ③ 组蛋白修饰 ④ H3组蛋白巯基暴露 (三)正性调节占主导 (四)转录与翻译分隔进行 (五)转录后修饰、加工
- 43. 三、真核基因转录激活调节 (一)顺式作用元件 1. 启动子 真核基因启动子是RNA聚合酶结合位点周围的一组转录控制组件,至少包括一个转录起始点以及一个以上的功能组件。
- 46. (二)反式作用因子 1. 转录调节因子分类(按功能特性) * 基本转录因子(general transcription factors) 是RNA聚合酶结合启动子所必需的一组蛋白因子,决定三种RNA(mRNA、tRNA及rRNA)转录的类别。
- 47. * 特异转录因子(special transcription factors) 为个别基因转录所必需,决定该基因的时间、空间特异性表达。 转录激活因子 转录抑制因子
- 48. 2. 转录调节因子结构
- 49. 最常见的DNA结合域 1. 锌指(zinc finger) C —— Cys H —— His 常结合GC盒
- 50. Zn Cys His
- 51. 2. α-螺旋 常结合CAAT盒
- 52. (三)mRNA 转录激活及其调节 真核RNA聚合酶Ⅱ在转录因子帮助下,形成的转录起始复合物 TBP相关因子
- 54. Скачать презентацию

Слайд 3一、基因表达的概念
* 基因组(genome)
一个细胞或病毒所携带的全部遗传信息或整套基因。
基因经过转录、翻译,产生具有特异生物学功能的蛋白质分子的过程。
* 基因表达(gene expression)
基因表达是受调控的
一、基因表达的概念
* 基因组(genome)
一个细胞或病毒所携带的全部遗传信息或整套基因。
基因经过转录、翻译,产生具有特异生物学功能的蛋白质分子的过程。
* 基因表达(gene expression)
基因表达是受调控的

Слайд 4二、基因表达的时间性及空间性
(一)时间特异性
二、基因表达的时间性及空间性
(一)时间特异性

Слайд 5(二)空间特异性
基因表达伴随时间顺序所表现出的这种分布差异,实际上是由细胞在器官的分布决定的,所以空间特异性又称细胞或组织特异性(cell or tissue specificity)。
在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,称之为基因表达的空间特异性(spatial specificity)。
(二)空间特异性
基因表达伴随时间顺序所表现出的这种分布差异,实际上是由细胞在器官的分布决定的,所以空间特异性又称细胞或组织特异性(cell or tissue specificity)。
在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,称之为基因表达的空间特异性(spatial specificity)。

Слайд 6三、基因表达的方式
按对刺激的反应性,基因表达的方式分为:
(一)组成性表达
某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因(housekeeping gene)。
三、基因表达的方式
按对刺激的反应性,基因表达的方式分为:
(一)组成性表达
某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因(housekeeping gene)。

Слайд 7无论表达水平高低,管家基因较少受环境因素影响,而是在个体各个生长阶段的大多数或几乎全部组织中持续表达,或变化很小。区别于其他基因,这类基因表达被视为组成性基因表达(constitutive gene expression)。
无论表达水平高低,管家基因较少受环境因素影响,而是在个体各个生长阶段的大多数或几乎全部组织中持续表达,或变化很小。区别于其他基因,这类基因表达被视为组成性基因表达(constitutive gene expression)。

Слайд 8(二)诱导和阻遏表达
在特定环境信号刺激下,相应的基因被激活,基因表达产物增加,这种基因称为可诱导基因。
可诱导基因在特定环境中表达增强的过程,称为诱导(induction)。
如果基因对环境信号应答是被抑制,这种基因是可阻遏基因。可阻遏基因表达产物水平降低的过程称为阻遏(repression)。
(二)诱导和阻遏表达
在特定环境信号刺激下,相应的基因被激活,基因表达产物增加,这种基因称为可诱导基因。
可诱导基因在特定环境中表达增强的过程,称为诱导(induction)。
如果基因对环境信号应答是被抑制,这种基因是可阻遏基因。可阻遏基因表达产物水平降低的过程称为阻遏(repression)。

Слайд 9在一定机制控制下,功能上相关的一组基因,无论其为何种表达方式,均需协调一致、共同表达,即为协调表达(coordinate expression),这种调节称为协调调节(coordinate regulation)。
在一定机制控制下,功能上相关的一组基因,无论其为何种表达方式,均需协调一致、共同表达,即为协调表达(coordinate expression),这种调节称为协调调节(coordinate regulation)。

Слайд 10四、基因表达调控的生物学意义
(一)适应环境、维持生长和增殖
(二)维持个体发育与分化
四、基因表达调控的生物学意义
(一)适应环境、维持生长和增殖
(二)维持个体发育与分化

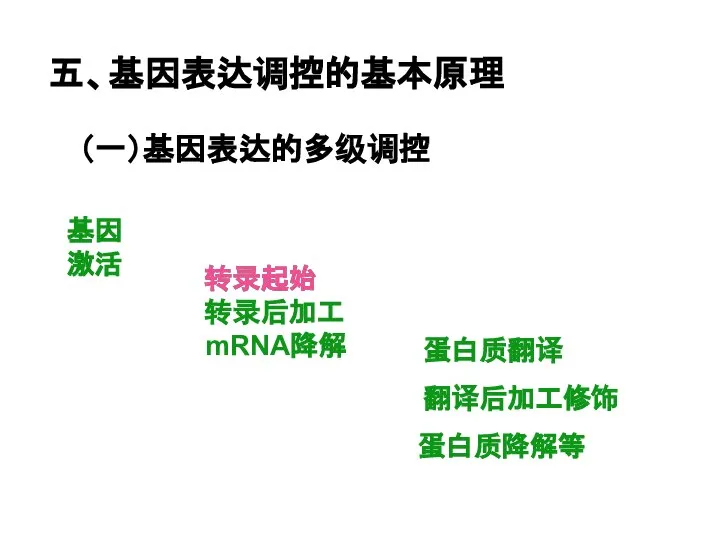
Слайд 11五、基因表达调控的基本原理
(一)基因表达的多级调控
基因激活
转录起始
转录后加工
mRNA降解
转录起始
五、基因表达调控的基本原理
(一)基因表达的多级调控
基因激活
转录起始
转录后加工
mRNA降解
转录起始
Слайд 12(二)基因转录激活调节基本要素
基因表达的调节与基因的结构、性质,生物个体或细胞所处的内、外环境,以及细胞内所存在的转录调节蛋白有关。
1. 特异DNA序列和调节蛋白质
(二)基因转录激活调节基本要素
基因表达的调节与基因的结构、性质,生物个体或细胞所处的内、外环境,以及细胞内所存在的转录调节蛋白有关。
1. 特异DNA序列和调节蛋白质

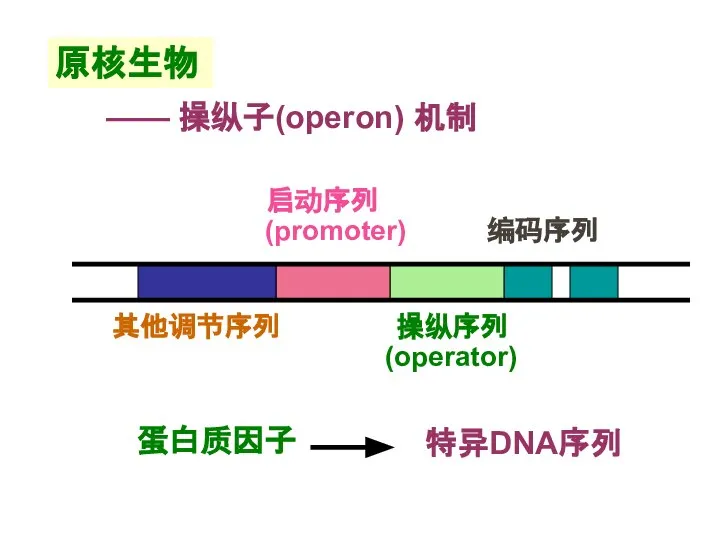
Слайд 13原核生物
—— 操纵子(operon) 机制
原核生物
—— 操纵子(operon) 机制
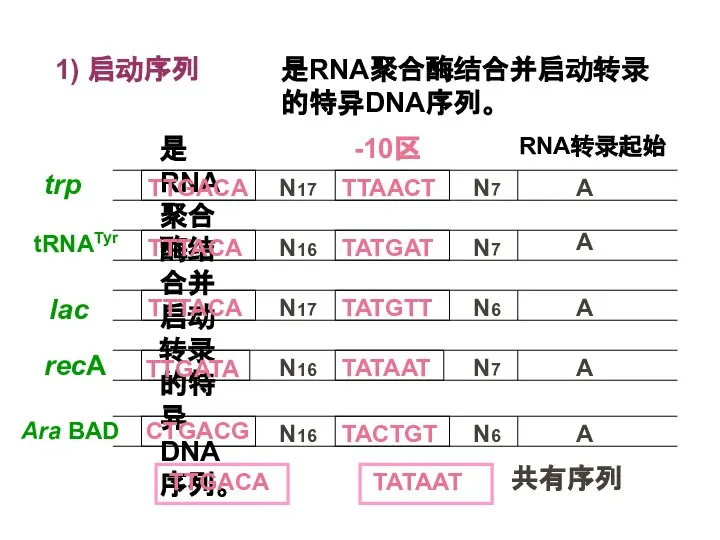
Слайд 15共有序列(consensus sequence) 决定启动序列的转录活性大小。
某些特异因子(蛋白质)决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力。
共有序列(consensus sequence) 决定启动序列的转录活性大小。
某些特异因子(蛋白质)决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力。

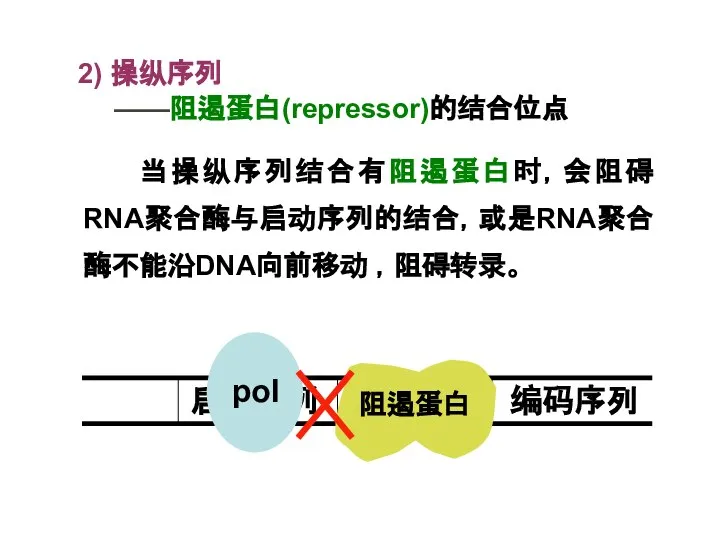
Слайд 162) 操纵序列
——阻遏蛋白(repressor)的结合位点
当操纵序列结合有阻遏蛋白时,会阻碍RNA聚合酶与启动序列的结合,或是RNA聚合酶不能沿DNA向前移动 ,阻碍转录。
2) 操纵序列
——阻遏蛋白(repressor)的结合位点
当操纵序列结合有阻遏蛋白时,会阻碍RNA聚合酶与启动序列的结合,或是RNA聚合酶不能沿DNA向前移动 ,阻碍转录。
Слайд 173) 其他调节序列、调节蛋白
例如
激活蛋白(activator)可结合启动序列邻近的DNA序列,促进RNA聚合酶与启动序列的结合,增强RNA聚合酶活性。
有些基因在没有激活蛋白存在时,RNA聚合酶很少或完全不能结合启动序列。
3) 其他调节序列、调节蛋白
例如
激活蛋白(activator)可结合启动序列邻近的DNA序列,促进RNA聚合酶与启动序列的结合,增强RNA聚合酶活性。
有些基因在没有激活蛋白存在时,RNA聚合酶很少或完全不能结合启动序列。

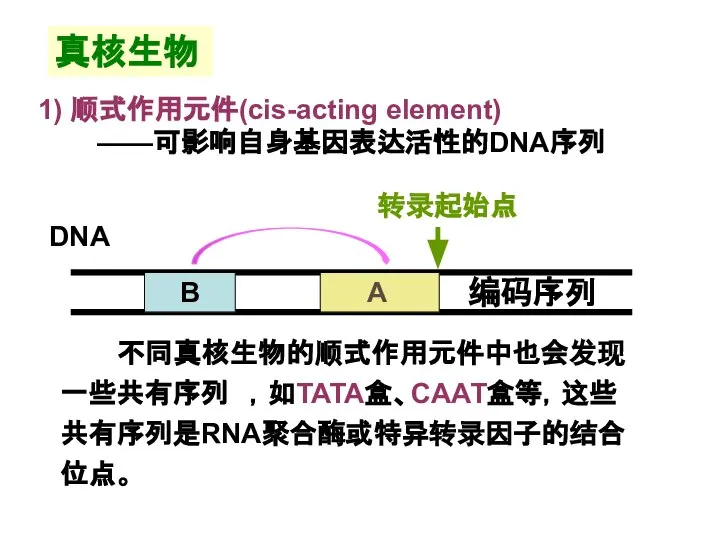
Слайд 18真核生物
不同真核生物的顺式作用元件中也会发现一些共有序列 ,如TATA盒、CAAT盒等,这些共有序列是RNA聚合酶或特异转录因子的结合位点。
真核生物
不同真核生物的顺式作用元件中也会发现一些共有序列 ,如TATA盒、CAAT盒等,这些共有序列是RNA聚合酶或特异转录因子的结合位点。
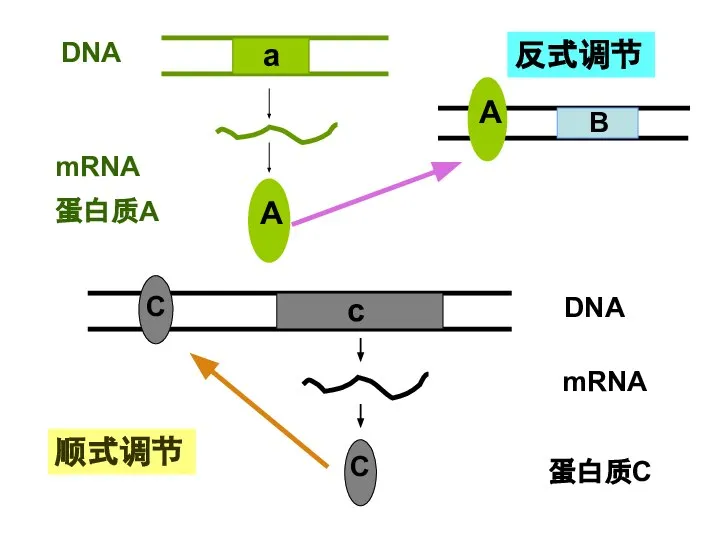
Слайд 192) 真核基因的调节蛋白
还有蛋白质因子可特异识别、结合自身基因的调节序列,调节自身基因的表达,称顺式作用。
由某一基因表达产生的蛋白质因子,通过与另一基因的特异的顺式作用元件相互作用,调节其表达。
反式作用因子(trans-acting factor)
这种调节作用称为反式作用。
2) 真核基因的调节蛋白
还有蛋白质因子可特异识别、结合自身基因的调节序列,调节自身基因的表达,称顺式作用。
由某一基因表达产生的蛋白质因子,通过与另一基因的特异的顺式作用元件相互作用,调节其表达。
反式作用因子(trans-acting factor)
这种调节作用称为反式作用。

Слайд 20反式调节
顺式调节
反式调节
顺式调节
Слайд 21指的是反式作用因子与顺式作用元件之间的特异识别及结合。通常是非共价结合,被识别的DNA结合位点通常呈对称、或不完全对称结构。
绝大多数调节蛋白质结合DNA前,需通过蛋白质-蛋白质相互作用,形成二聚体(dimer)或多聚体(polymer)。
指的是反式作用因子与顺式作用元件之间的特异识别及结合。通常是非共价结合,被识别的DNA结合位点通常呈对称、或不完全对称结构。
绝大多数调节蛋白质结合DNA前,需通过蛋白质-蛋白质相互作用,形成二聚体(dimer)或多聚体(polymer)。

Слайд 223. RNA聚合酶
⑴ 原核启动序列/真核启动子与RNA聚合酶活性
RNA聚合酶与其的亲和力,影响转录。
⑵ 调节蛋白与RNA聚合酶活性
一些特异调节蛋白在适当环境信号刺激下表达,然后通过DNA-蛋白质、蛋白质-蛋白质相互作用影响RNA聚合酶活性。
3. RNA聚合酶
⑴ 原核启动序列/真核启动子与RNA聚合酶活性
RNA聚合酶与其的亲和力,影响转录。
⑵ 调节蛋白与RNA聚合酶活性
一些特异调节蛋白在适当环境信号刺激下表达,然后通过DNA-蛋白质、蛋白质-蛋白质相互作用影响RNA聚合酶活性。

Слайд 23第 二 节
原核基因转录调节
Regulation of Prokaryotic
Gene Transcription
第 二 节
原核基因转录调节
Regulation of Prokaryotic
Gene Transcription

Слайд 24(一)σ因子决定RNA聚合酶识别特异性
(二)操纵子模型的普遍性
(三)阻遏蛋白与阻遏机制的普遍性
(一)σ因子决定RNA聚合酶识别特异性
(二)操纵子模型的普遍性
(三)阻遏蛋白与阻遏机制的普遍性

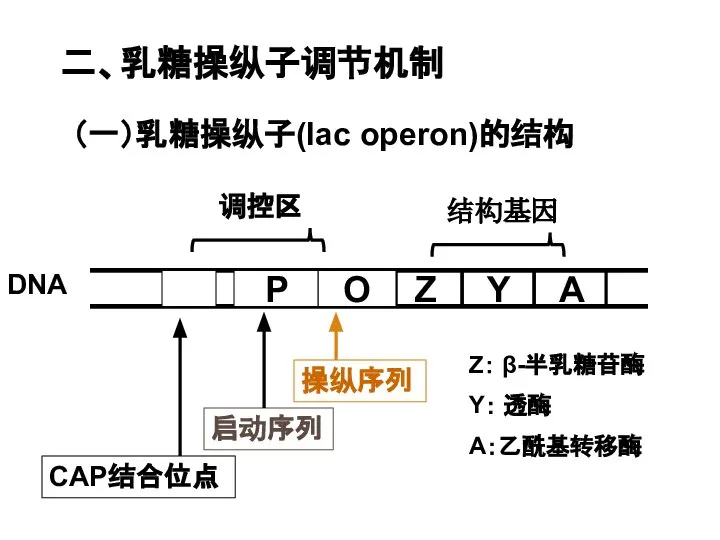
Слайд 25二、乳糖操纵子调节机制
(一)乳糖操纵子(lac operon)的结构
调控区
CAP结合位点
二、乳糖操纵子调节机制
(一)乳糖操纵子(lac operon)的结构
调控区
CAP结合位点
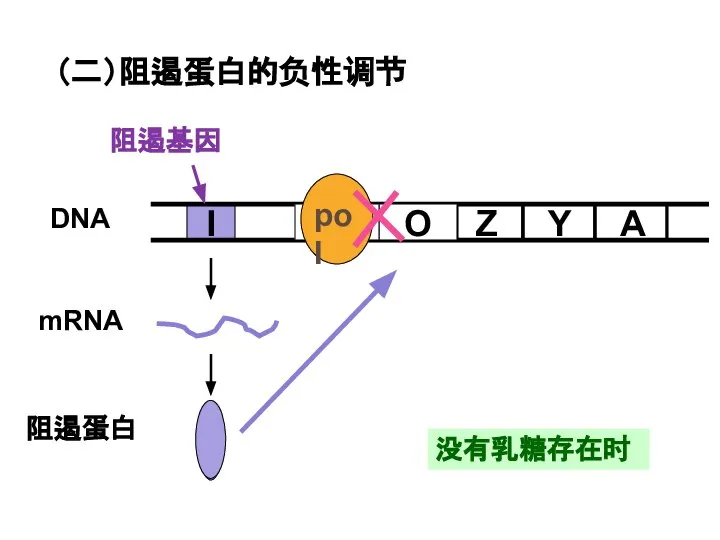
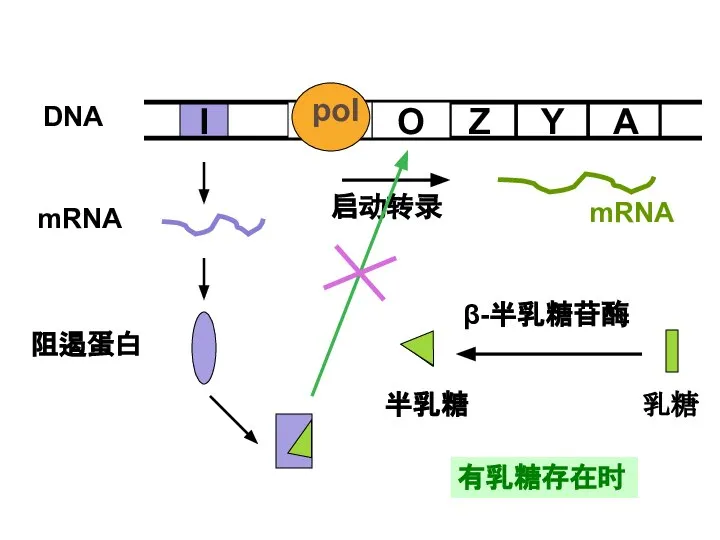
Слайд 26没有乳糖存在时
(二)阻遏蛋白的负性调节
没有乳糖存在时
(二)阻遏蛋白的负性调节
Слайд 27有乳糖存在时
有乳糖存在时
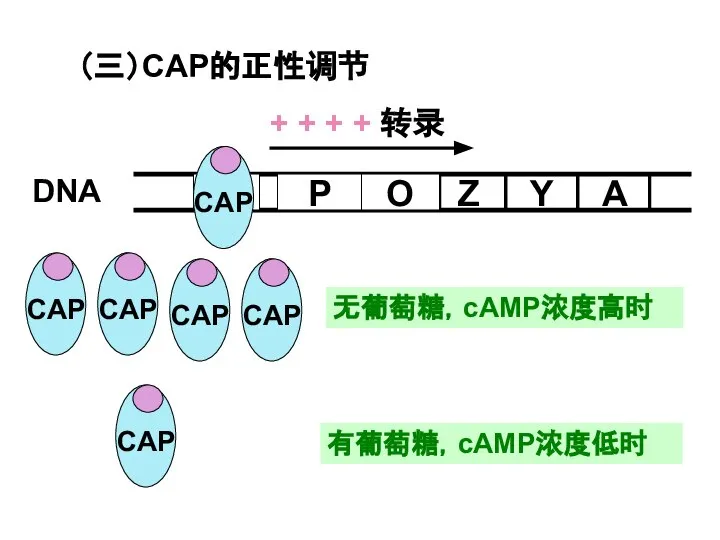
Слайд 28无葡萄糖,cAMP浓度高时
有葡萄糖,cAMP浓度低时
(三)CAP的正性调节
无葡萄糖,cAMP浓度高时
有葡萄糖,cAMP浓度低时
(三)CAP的正性调节
Слайд 29(四)协调调节
※当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用;
※如无CAP存在,即使没有阻遏蛋白与操纵序列结合,操纵子仍无转录活性。
单纯乳糖存在时,细菌利用乳糖作碳源;
若有葡萄糖或葡萄糖/乳糖共同存在时,细菌首先利用葡萄糖。
葡萄糖对 lac 操纵子的阻遏作用称分解代谢阻遏(catabolic repression)。
(四)协调调节
※当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用;
※如无CAP存在,即使没有阻遏蛋白与操纵序列结合,操纵子仍无转录活性。
单纯乳糖存在时,细菌利用乳糖作碳源;
若有葡萄糖或葡萄糖/乳糖共同存在时,细菌首先利用葡萄糖。
葡萄糖对 lac 操纵子的阻遏作用称分解代谢阻遏(catabolic repression)。

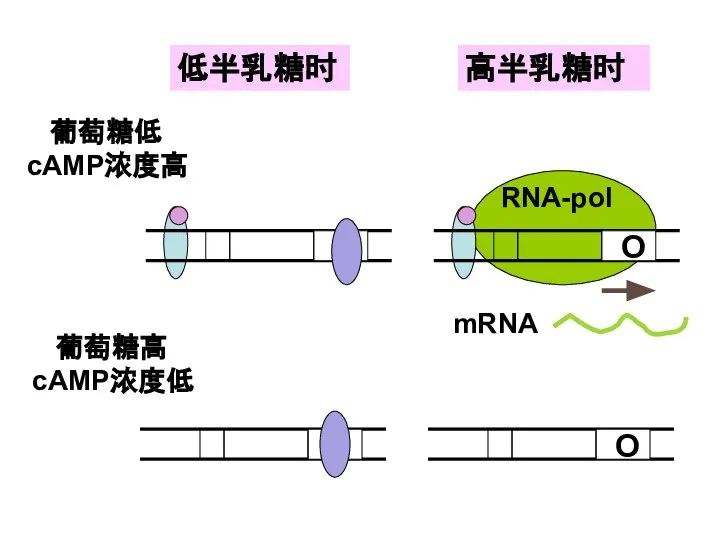
Слайд 30低半乳糖时
高半乳糖时
葡萄糖低 cAMP浓度高
葡萄糖高cAMP浓度低
低半乳糖时
高半乳糖时
葡萄糖低 cAMP浓度高
葡萄糖高cAMP浓度低
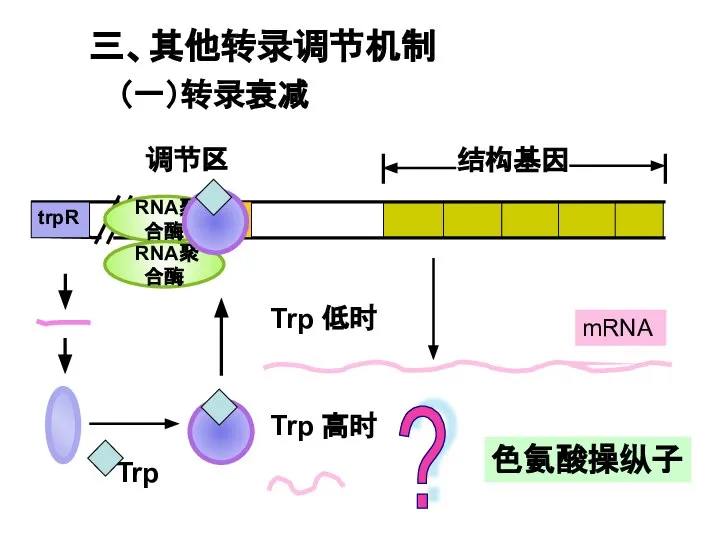
Слайд 31Trp 高时
Trp 低时
mRNA
O
P
trpR
调节区
结构基因
RNA聚合酶
RNA聚合酶
?
三、其他转录调节机制
(一)转录衰减
色氨酸操纵子
Trp 高时
Trp 低时
mRNA
O
P
trpR
调节区
结构基因
RNA聚合酶
RNA聚合酶
?
三、其他转录调节机制
(一)转录衰减
色氨酸操纵子
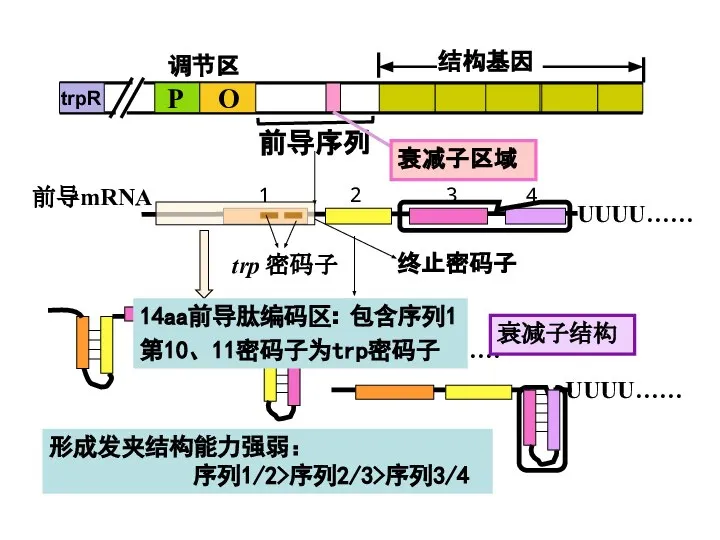
Слайд 32前导序列
第10、11密码子为trp密码子
14aa前导肽编码区:
包含序列1
形成发夹结构能力强弱:
序列1/2>序列2/3>序列3/4
前导序列
第10、11密码子为trp密码子
14aa前导肽编码区:
包含序列1
形成发夹结构能力强弱:
序列1/2>序列2/3>序列3/4
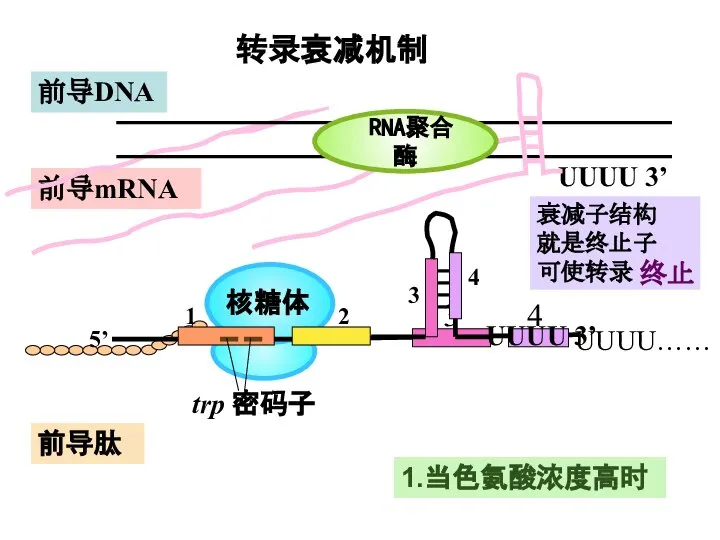
Слайд 33UUUU 3’
前导肽
前导mRNA
1.当色氨酸浓度高时
转录衰减机制
衰减子结构
就是终止子
可使转录
RNA聚合酶
终止
UUUU 3’
前导肽
前导mRNA
1.当色氨酸浓度高时
转录衰减机制
衰减子结构
就是终止子
可使转录
RNA聚合酶
终止
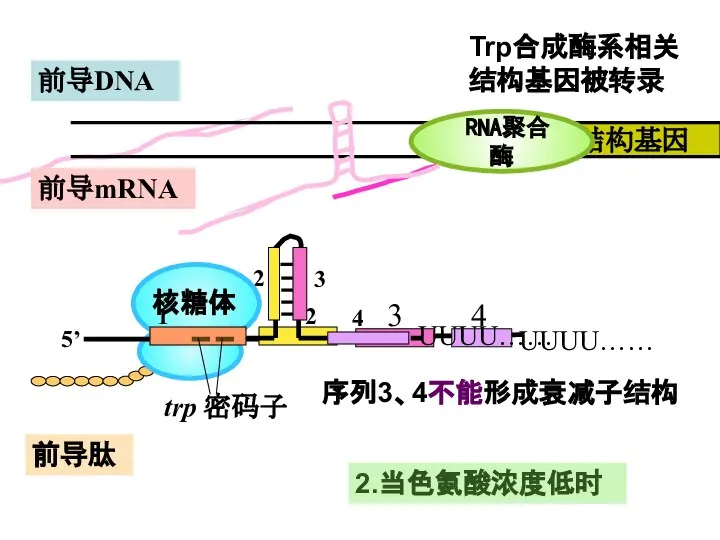
Слайд 34前导肽
前导mRNA
RNA聚合酶
2.当色氨酸浓度低时
Trp合成酶系相关
结构基因被转录
序列3、4不能形成衰减子结构
前导肽
前导mRNA
RNA聚合酶
2.当色氨酸浓度低时
Trp合成酶系相关
结构基因被转录
序列3、4不能形成衰减子结构
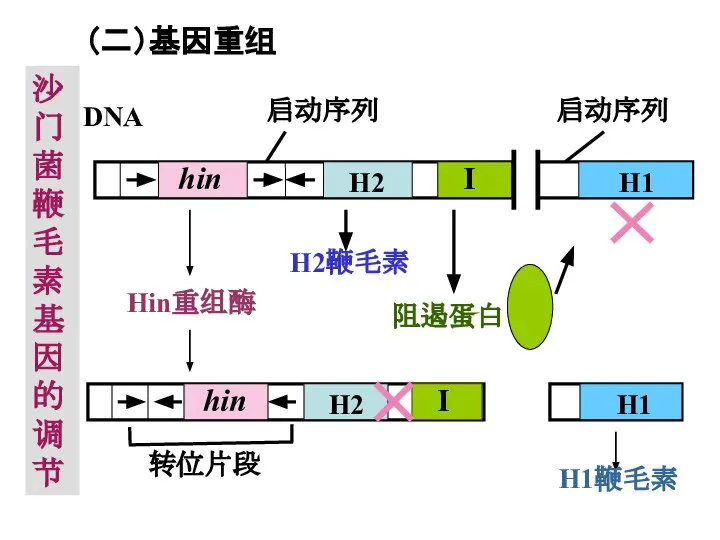
Слайд 35(二)基因重组
沙门菌鞭毛素基因的调节
(二)基因重组
沙门菌鞭毛素基因的调节
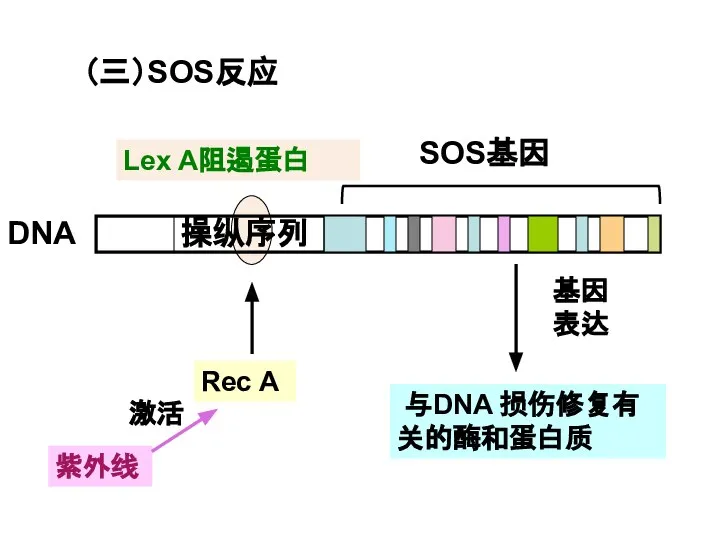
Слайд 36(三)SOS反应
紫外线
与DNA 损伤修复有关的酶和蛋白质
Lex A阻遏蛋白
DNA
(三)SOS反应
紫外线
与DNA 损伤修复有关的酶和蛋白质
Lex A阻遏蛋白
DNA
Слайд 37第 三 节
真核基因转录调节
Regulation of Eukaryotic
Gene Transcription
第 三 节
真核基因转录调节
Regulation of Eukaryotic
Gene Transcription

Слайд 38一、真核基因组结构特点
(一)真核基因组结构庞大
一、真核基因组结构特点
(一)真核基因组结构庞大

Слайд 39(二)单顺反子
单顺反子(monocistron)
即一个编码基因转录生成一个mRNA分子,经翻译生成一条多肽链。
(三)重复序列
(四)基因不连续性
(二)单顺反子
单顺反子(monocistron)
即一个编码基因转录生成一个mRNA分子,经翻译生成一条多肽链。
(三)重复序列
(四)基因不连续性

Слайд 40二、真核基因表达调控特点
(一)RNA聚合酶
(二)活性染色体结构变化
1. 对核酸酶敏感
活化基因常有超敏位点,位于调节蛋白结合位点附近。
二、真核基因表达调控特点
(一)RNA聚合酶
(二)活性染色体结构变化
1. 对核酸酶敏感
活化基因常有超敏位点,位于调节蛋白结合位点附近。

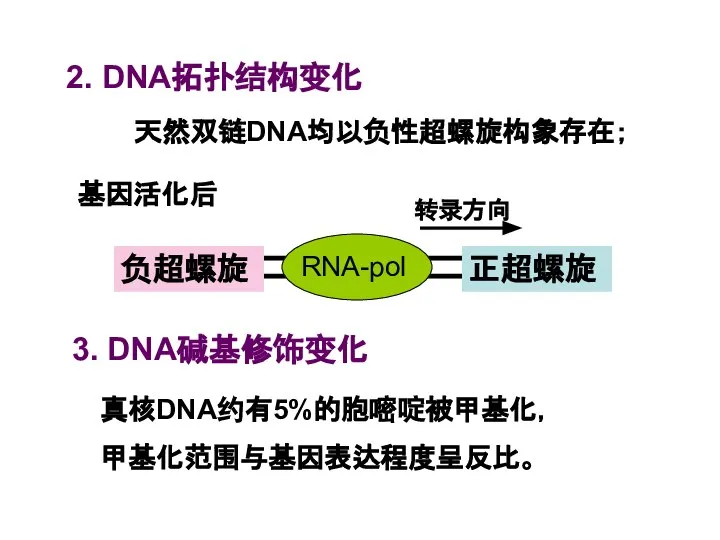
Слайд 412. DNA拓扑结构变化
天然双链DNA均以负性超螺旋构象存在;
基因活化后
3. DNA碱基修饰变化
2. DNA拓扑结构变化
天然双链DNA均以负性超螺旋构象存在;
基因活化后
3. DNA碱基修饰变化
Слайд 424. 组蛋白变化
① 富含Lys组蛋白水平降低
② H2A, H2B二聚体不稳定性增加
③ 组蛋白修饰
④ H3组蛋白巯基暴露
(三)正性调节占主导
(四)转录与翻译分隔进行
(五)转录后修饰、加工
4. 组蛋白变化
① 富含Lys组蛋白水平降低
② H2A, H2B二聚体不稳定性增加
③ 组蛋白修饰
④ H3组蛋白巯基暴露
(三)正性调节占主导
(四)转录与翻译分隔进行
(五)转录后修饰、加工

Слайд 43三、真核基因转录激活调节
(一)顺式作用元件
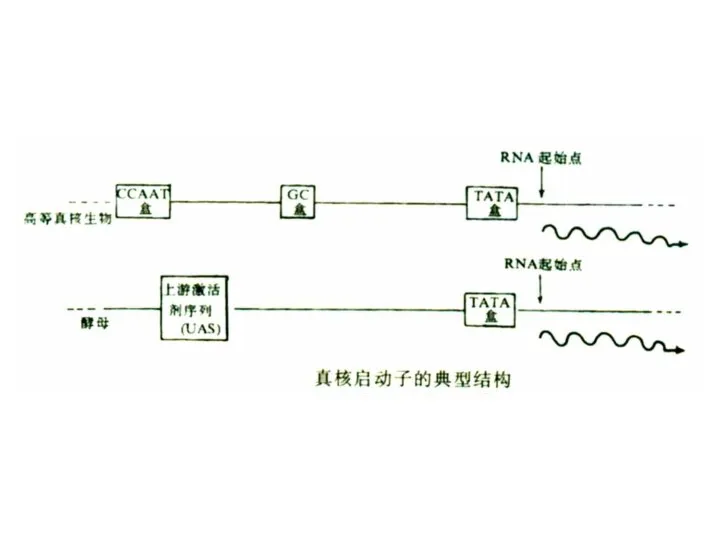
1. 启动子
真核基因启动子是RNA聚合酶结合位点周围的一组转录控制组件,至少包括一个转录起始点以及一个以上的功能组件。
三、真核基因转录激活调节
(一)顺式作用元件
1. 启动子
真核基因启动子是RNA聚合酶结合位点周围的一组转录控制组件,至少包括一个转录起始点以及一个以上的功能组件。


Слайд 46(二)反式作用因子
1. 转录调节因子分类(按功能特性)
* 基本转录因子(general transcription factors)
是RNA聚合酶结合启动子所必需的一组蛋白因子,决定三种RNA(mRNA、tRNA及rRNA)转录的类别。
(二)反式作用因子
1. 转录调节因子分类(按功能特性)
* 基本转录因子(general transcription factors)
是RNA聚合酶结合启动子所必需的一组蛋白因子,决定三种RNA(mRNA、tRNA及rRNA)转录的类别。

Слайд 47* 特异转录因子(special transcription factors)
为个别基因转录所必需,决定该基因的时间、空间特异性表达。
转录激活因子
转录抑制因子
* 特异转录因子(special transcription factors)
为个别基因转录所必需,决定该基因的时间、空间特异性表达。
转录激活因子
转录抑制因子

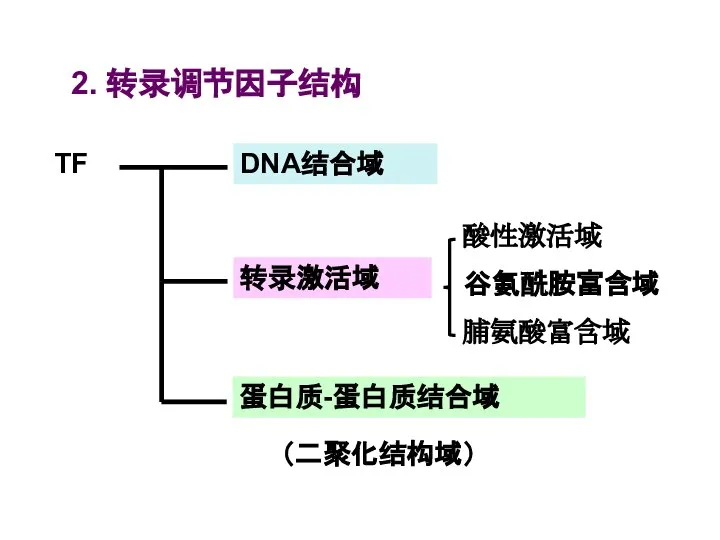
Слайд 482. 转录调节因子结构
2. 转录调节因子结构
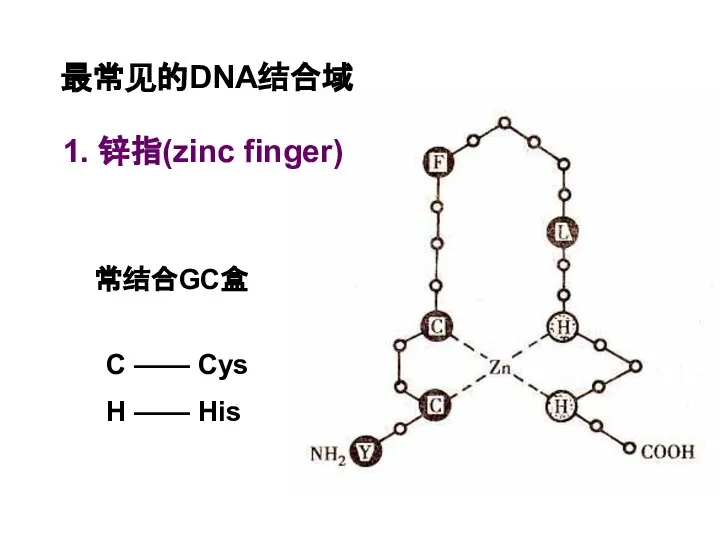
Слайд 49最常见的DNA结合域
1. 锌指(zinc finger)
C —— Cys
H —— His
常结合GC盒
最常见的DNA结合域
1. 锌指(zinc finger)
C —— Cys
H —— His
常结合GC盒
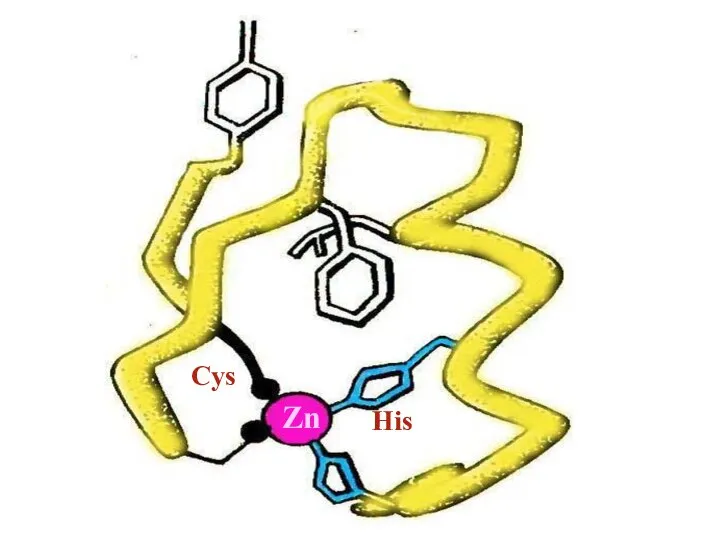
Слайд 50Zn
Cys
His
Zn
Cys
His
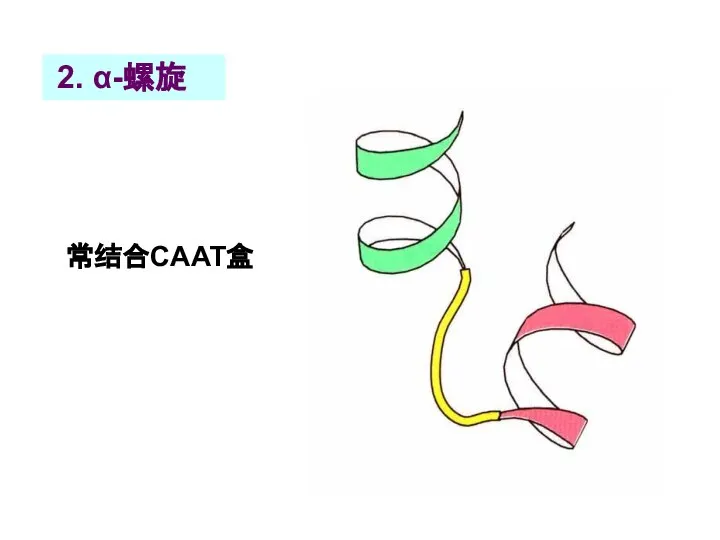
Слайд 51 2. α-螺旋
常结合CAAT盒
2. α-螺旋
常结合CAAT盒
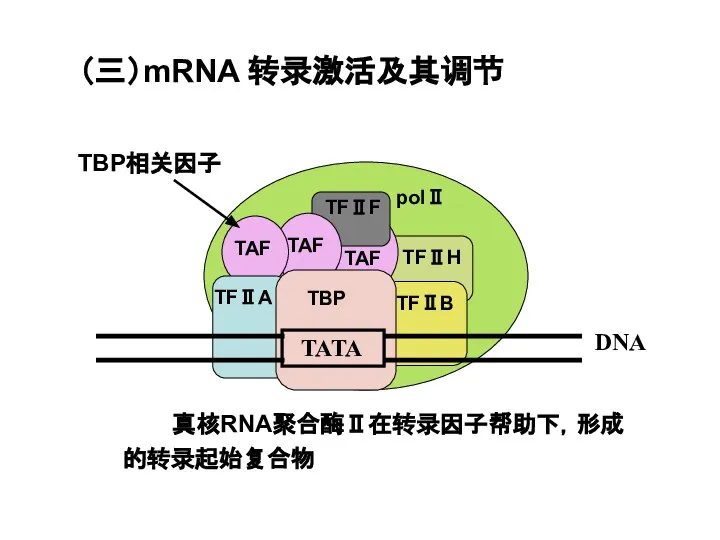
Слайд 52(三)mRNA 转录激活及其调节
真核RNA聚合酶Ⅱ在转录因子帮助下,形成的转录起始复合物
TBP相关因子
(三)mRNA 转录激活及其调节
真核RNA聚合酶Ⅱ在转录因子帮助下,形成的转录起始复合物
TBP相关因子
 лекция4-5 ЦНС
лекция4-5 ЦНС Как стареют лица
Как стареют лица Изучить признаки семейства Бобовые
Изучить признаки семейства Бобовые Подорожник
Подорожник Состав, строение и свойства белков
Состав, строение и свойства белков Строение клетки
Строение клетки Популяции. Изменчивость популяции
Популяции. Изменчивость популяции Свойства биогеоценозов
Свойства биогеоценозов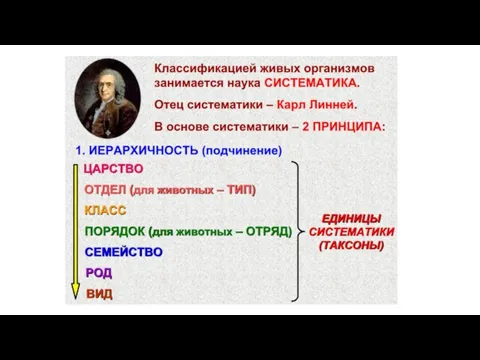 Многообразие бактерий
Многообразие бактерий Fiziologia_analizatorov
Fiziologia_analizatorov Грибы рода Aspergillus
Грибы рода Aspergillus Презентация на тему Эволюция кровеносной системы у животных
Презентация на тему Эволюция кровеносной системы у животных  Сенсорная система
Сенсорная система Транспорт веществ через мембрану
Транспорт веществ через мембрану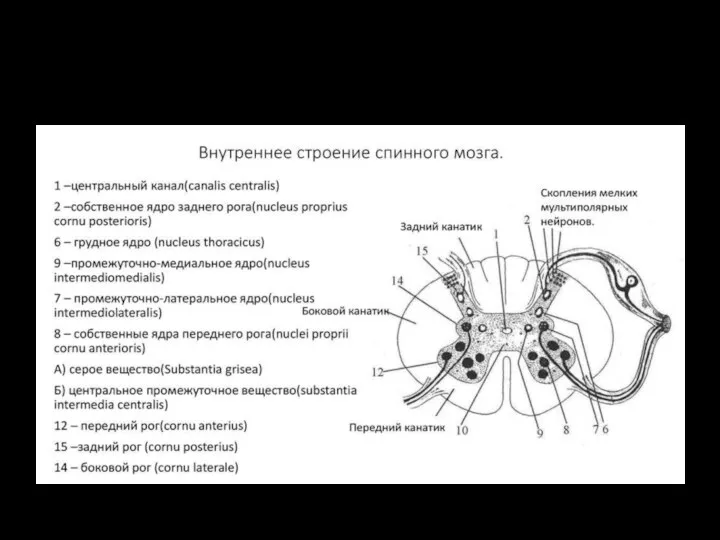 Спинной мозг. Внутреннее строение
Спинной мозг. Внутреннее строение Презентация на тему Увеличительные приборы : микроскоп и лупа
Презентация на тему Увеличительные приборы : микроскоп и лупа  _Как работают в лаборатории
_Как работают в лаборатории Китова акула
Китова акула Нетрадиционные методы лечения. Биолокация
Нетрадиционные методы лечения. Биолокация 3._Экспрессия_генов_как_функционирование_генома__и_ее_регуляция
3._Экспрессия_генов_как_функционирование_генома__и_ее_регуляция Животные Кубани
Животные Кубани ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ Хавортия
Хавортия «Размножение и индивидуальное развитие организмов». Подготовка к ЕГЭ. Учитель высшей квалификационной категории МОУ «СОШ № 7» Б
«Размножение и индивидуальное развитие организмов». Подготовка к ЕГЭ. Учитель высшей квалификационной категории МОУ «СОШ № 7» Б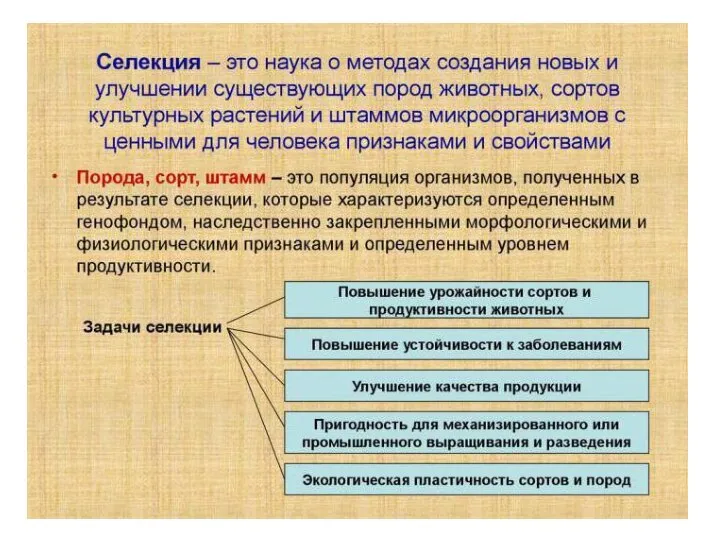 Селекция растений
Селекция растений Происхождение жизни на Земле
Происхождение жизни на Земле Цитология и генетика
Цитология и генетика Роль жиров в организме человека
Роль жиров в организме человека