- Обмен липидов. Анаболизм липидов

Содержание
- 2. СИНТЕЗ ЖИРНЫХ КИСЛОТ
- 3. СИНТЕЗ ЖИРНЫХ КИСЛОТ ЛОКАЛИЗАЦИЯ В КЛЕТКЕ: -ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16) -МИТОХОНДРИИ (элонгаза синтезирует
- 4. СИНТЕЗ ЖИРНЫХ КИСЛОТ Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при окислении глюкозы в
- 5. СИНТЕЗ ЖИРНЫХ КИСЛОТ Синтез жирных кислот требует: источник углеродного скелета: ацетил-КоА дополнительный источник углерода: СО2 (для
- 6. Выделяют три этапа образования пальмитиновой кислоты: транспортировка ацетил-КоА из митохондрий в цитоплазму; образование малонил-КоА из ацетил-КоА;
- 7. СИНТЕЗ ЖИРНЫХ КИСЛОТ Транспорт ацетил-КоА в цитоплазму
- 8. СИНТЕЗ ЖИРНЫХ КИСЛОТ Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных кислот Карбоксилирование ацетил-КоА
- 9. СИНТЕЗ ЖИРНЫХ КИСЛОТ Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ Пальмитоилсинтаза - димер из двух полипептидных цепей. Каждая субъединица
- 10. СИНТЕЗ ЖИРНЫХ КИСЛОТ 1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
- 11. СИНТЕЗ ЖИРНЫХ КИСЛОТ 2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего белка
- 12. 3. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2 СИНТЕЗ ЖИРНЫХ КИСЛОТ
- 13. СИНТЕЗ ЖИРНЫХ КИСЛОТ 4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
- 14. СИНТЕЗ ЖИРНЫХ КИСЛОТ 5. Дегидратация β-гидроксиацильного остатка
- 15. СИНТЕЗ ЖИРНЫХ КИСЛОТ 6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
- 16. СИНТЕЗ ЖИРНЫХ КИСЛОТ 7. Завершается первый этап синтеза перемещением радикала бутирила на свободную SH-группу цистеина Затем
- 17. СИНТЕЗ ЖИРНЫХ КИСЛОТ Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в свободную пальмитиновую кислоту
- 18. ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2- в качестве восстановителя.
- 19. ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ Основные жирные кислоты, образующиеся в организме человека в результате десатурации – пальмитоолеиновая (С16:1Δ9)
- 20. РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ Регулируется пищевым режимом: - тормозит голодание - активирует богатая углеводами пища 2.
- 21. СИНТЕЗ ХОЛЕСТЕРИНА Субстрат для синтеза холестерола- ацетил-КоА Активация синтеза холестерина происходит при поступлении в организм пищи,
- 22. СИНТЕЗ ХОЛЕСТЕРИНА ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА: • синтез мевалоната (С6) • синтез сквалена из мевалоната (С30)
- 23. СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА Регуляторная реакция синтеза холестерина
- 24. СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
- 25. СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
- 26. РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА 1. Регулируется пищевым режимом: • тормозит голодание • активирует богатая углеводами пища 2.
- 27. СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
- 28. СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ 1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани) 2. Ресинтез триацилглицеролов в
- 29. СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
- 31. Скачать презентацию

Слайд 3СИНТЕЗ ЖИРНЫХ КИСЛОТ
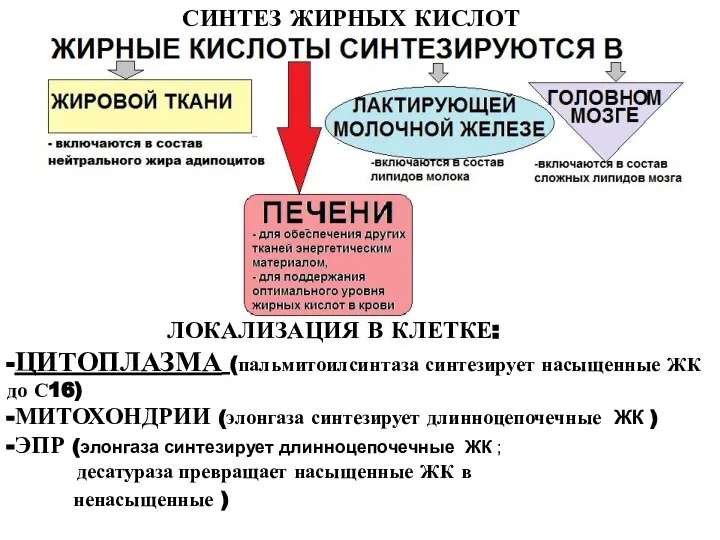
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16)
-МИТОХОНДРИИ
СИНТЕЗ ЖИРНЫХ КИСЛОТ
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ:
-ЦИТОПЛАЗМА (пальмитоилсинтаза синтезирует насыщенные ЖК до С16)
-МИТОХОНДРИИ
Слайд 4СИНТЕЗ ЖИРНЫХ КИСЛОТ
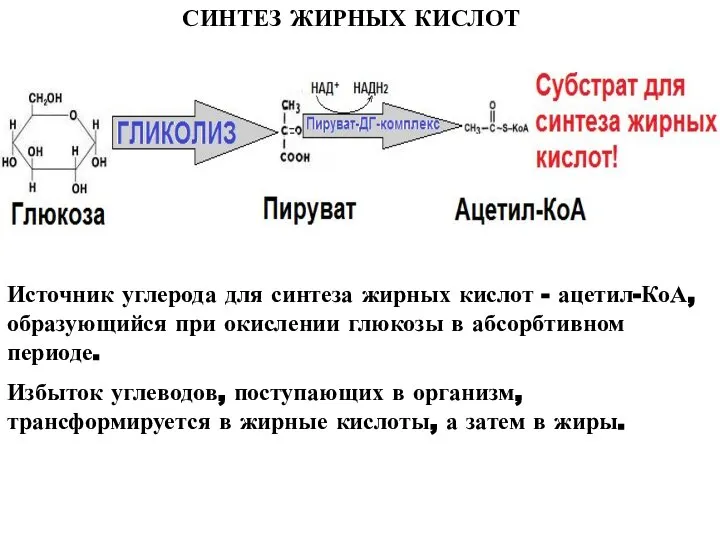
Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Источник углерода для синтеза жирных кислот - ацетил-КоА, образующийся при
Слайд 5СИНТЕЗ ЖИРНЫХ КИСЛОТ
Синтез жирных кислот требует:
источник углеродного скелета: ацетил-КоА
дополнительный источник углерода:
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Синтез жирных кислот требует:
источник углеродного скелета: ацетил-КоА
дополнительный источник углерода:

Слайд 6 Выделяют три этапа образования пальмитиновой кислоты:
транспортировка ацетил-КоА из митохондрий в цитоплазму;
образование
Выделяют три этапа образования пальмитиновой кислоты:
транспортировка ацетил-КоА из митохондрий в цитоплазму;
образование

Слайд 7СИНТЕЗ ЖИРНЫХ КИСЛОТ
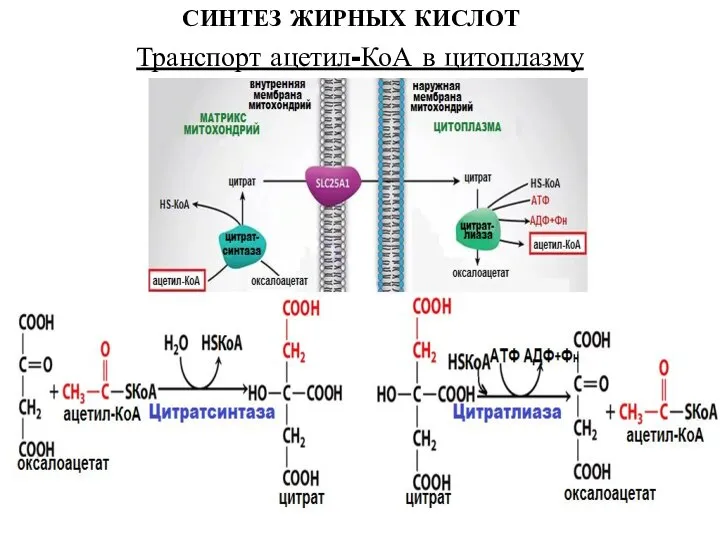
Транспорт ацетил-КоА в цитоплазму
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Транспорт ацетил-КоА в цитоплазму
Слайд 8СИНТЕЗ ЖИРНЫХ КИСЛОТ
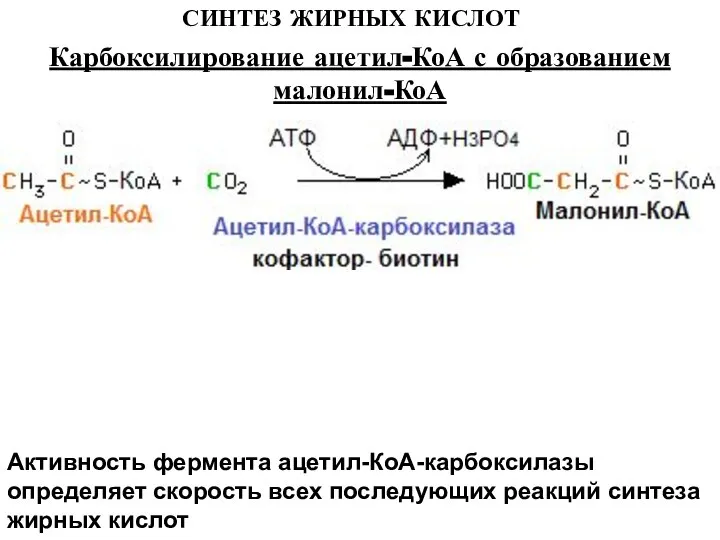
Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Активность фермента ацетил-КоА-карбоксилазы определяет скорость всех последующих реакций синтеза жирных
Слайд 9СИНТЕЗ ЖИРНЫХ КИСЛОТ
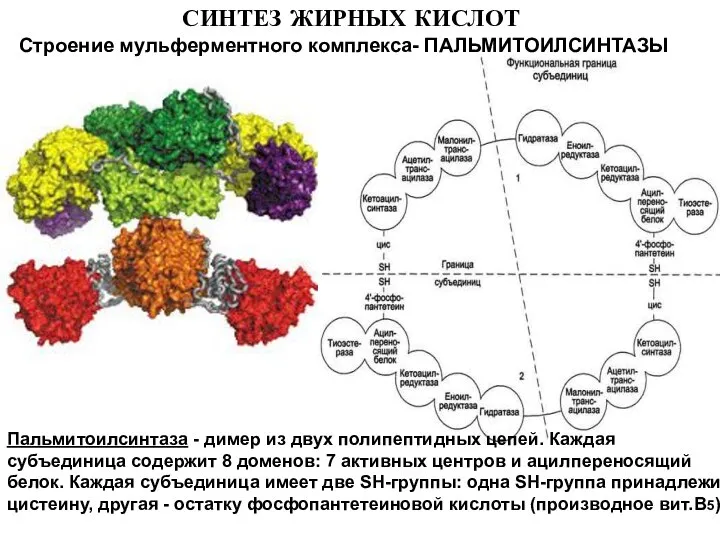
Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей.
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Строение мульферментного комплекса- ПАЛЬМИТОИЛСИНТАЗЫ
Пальмитоилсинтаза - димер из двух полипептидных цепей.
Слайд 10СИНТЕЗ ЖИРНЫХ КИСЛОТ
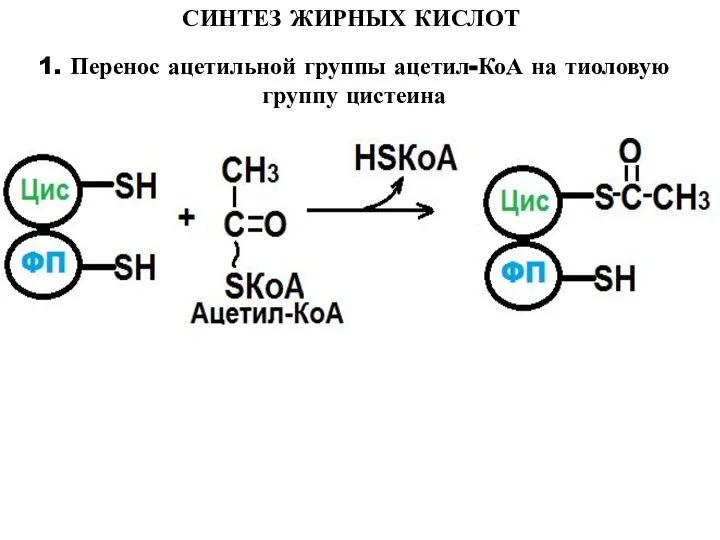
1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
СИНТЕЗ ЖИРНЫХ КИСЛОТ
1. Перенос ацетильной группы ацетил-КоА на тиоловую группу цистеина
Слайд 11СИНТЕЗ ЖИРНЫХ КИСЛОТ
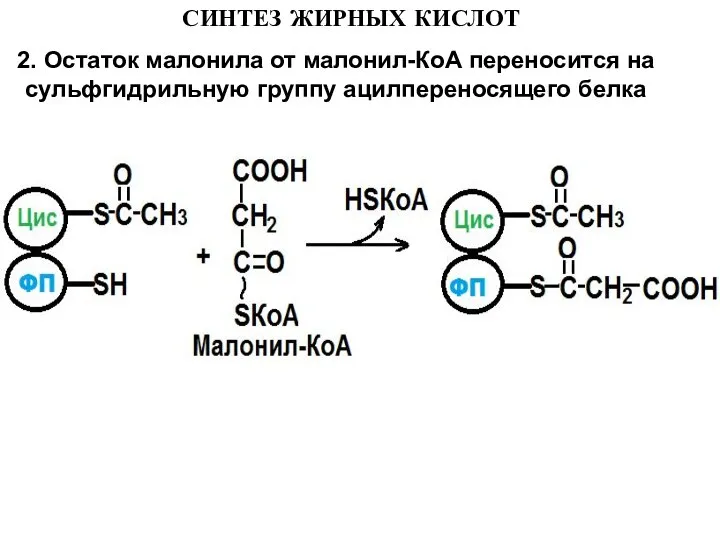
2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего
СИНТЕЗ ЖИРНЫХ КИСЛОТ
2. Остаток малонила от малонил-КоА переносится на сульфгидрильную группу ацилпереносящего
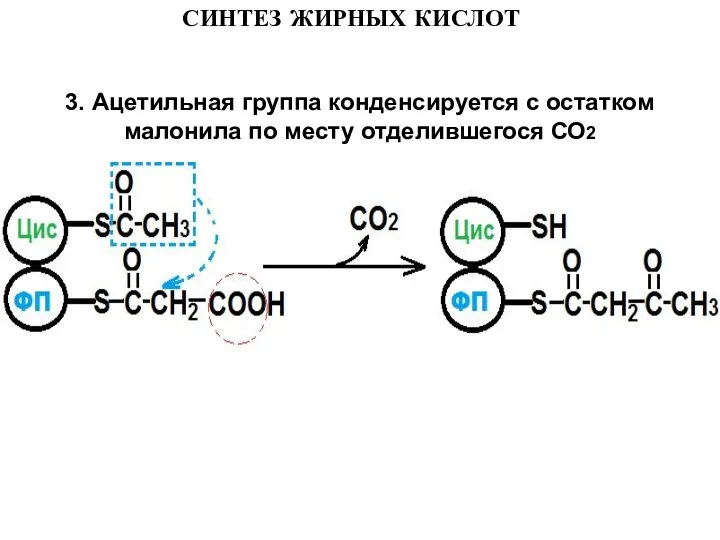
Слайд 123. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2
СИНТЕЗ
3. Ацетильная группа конденсируется с остатком малонила по месту отделившегося СО2
СИНТЕЗ
Слайд 13СИНТЕЗ ЖИРНЫХ КИСЛОТ
4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
СИНТЕЗ ЖИРНЫХ КИСЛОТ
4. Кетоацильный остаток восстанавливается, донор атомов водорода - НАДФН2
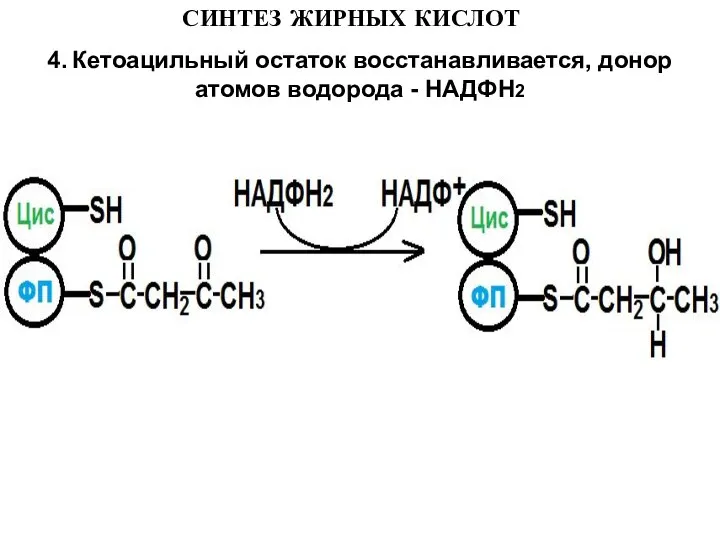
Слайд 14СИНТЕЗ ЖИРНЫХ КИСЛОТ
5. Дегидратация β-гидроксиацильного остатка
СИНТЕЗ ЖИРНЫХ КИСЛОТ
5. Дегидратация β-гидроксиацильного остатка
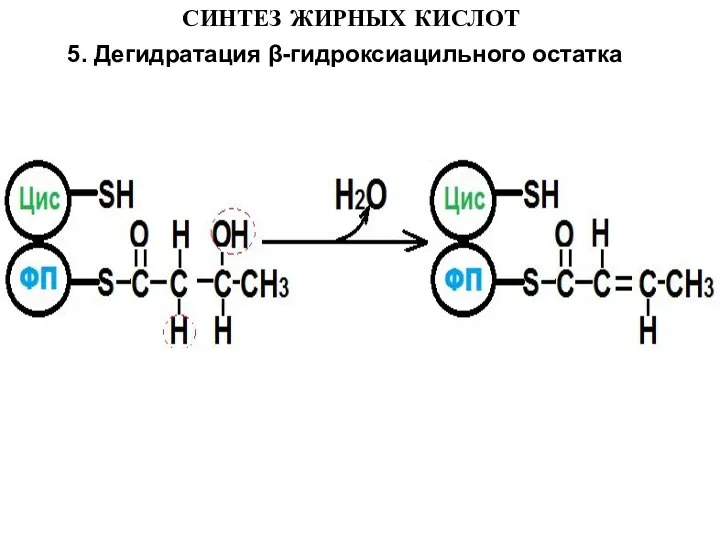
Слайд 15СИНТЕЗ ЖИРНЫХ КИСЛОТ
6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
СИНТЕЗ ЖИРНЫХ КИСЛОТ
6. Транс-еноильный остаток восстанавливается, донор атомов водорода - НАДФН2
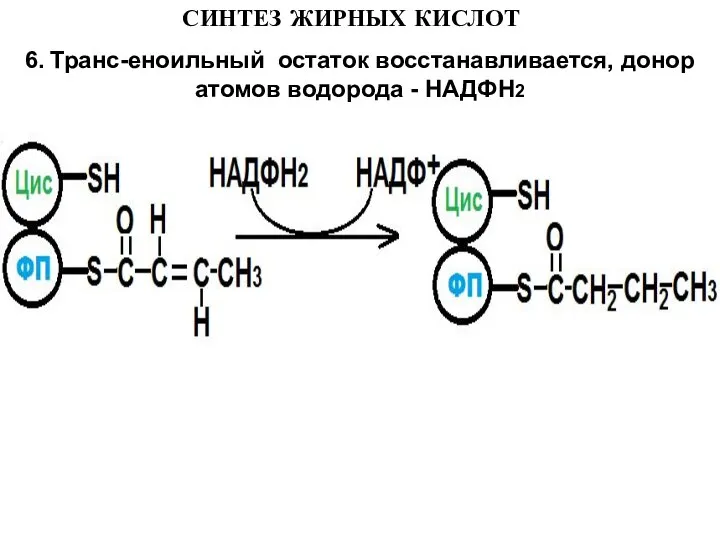
Слайд 16СИНТЕЗ ЖИРНЫХ КИСЛОТ
7. Завершается первый этап синтеза перемещением радикала бутирила на свободную
СИНТЕЗ ЖИРНЫХ КИСЛОТ
7. Завершается первый этап синтеза перемещением радикала бутирила на свободную
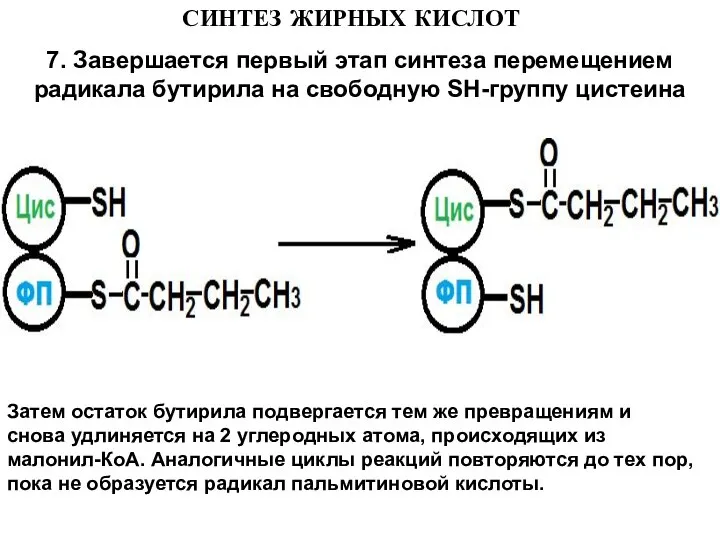
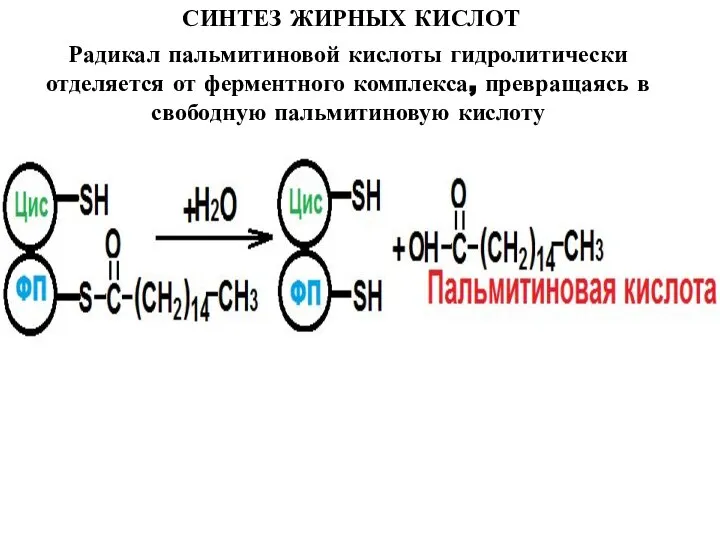
Слайд 17СИНТЕЗ ЖИРНЫХ КИСЛОТ
Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Радикал пальмитиновой кислоты гидролитически отделяется от ферментного комплекса, превращаясь в
Слайд 18ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ
Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2-
ЭЛОНГАЦИЯ ЖИРНЫХ КИСЛОТ
Элонгаза использует малонил-КоА в качестве донора углеродных атомов, а НАДФН2-

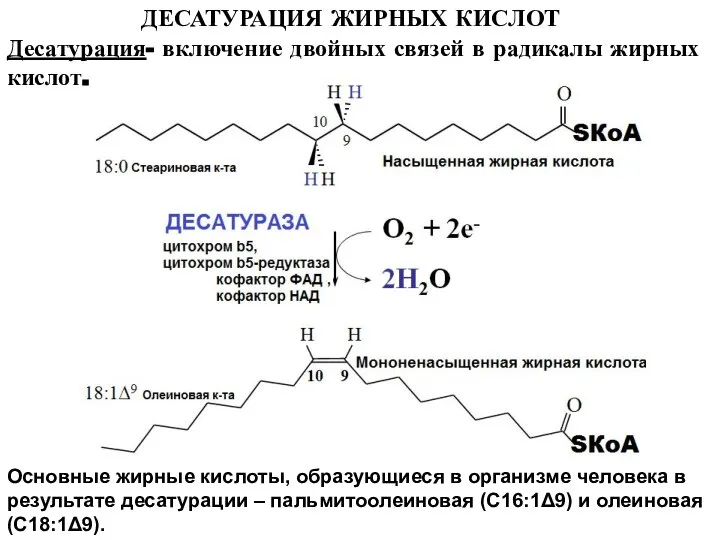
Слайд 19ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ
Основные жирные кислоты, образующиеся в организме человека в результате десатурации
ДЕСАТУРАЦИЯ ЖИРНЫХ КИСЛОТ
Основные жирные кислоты, образующиеся в организме человека в результате десатурации
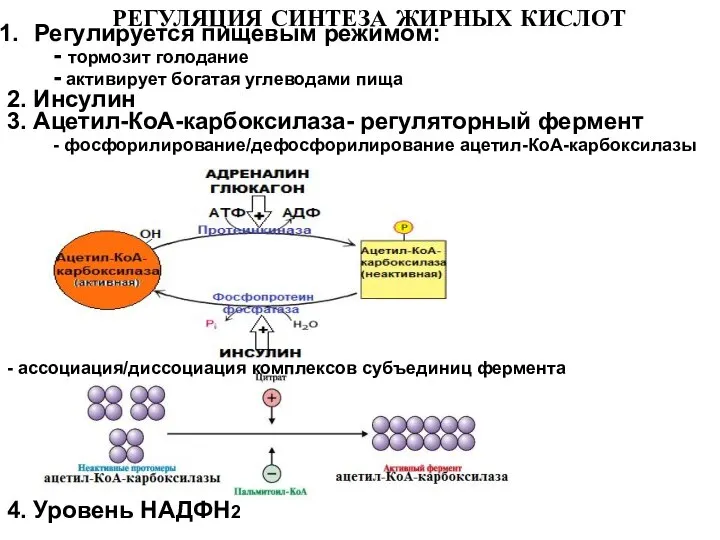
Слайд 20РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ
Регулируется пищевым режимом:
- тормозит голодание
- активирует
РЕГУЛЯЦИЯ СИНТЕЗА ЖИРНЫХ КИСЛОТ
Регулируется пищевым режимом:
- тормозит голодание
- активирует
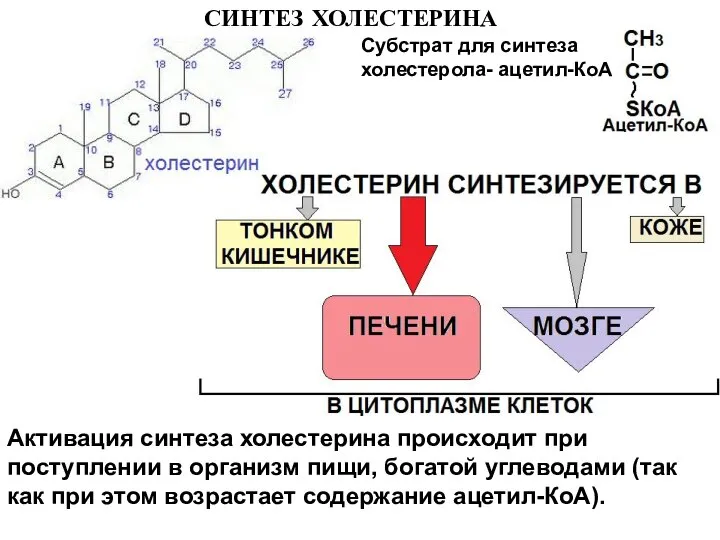
Слайд 21СИНТЕЗ ХОЛЕСТЕРИНА
Субстрат для синтеза холестерола- ацетил-КоА
Активация синтеза холестерина происходит при поступлении в
СИНТЕЗ ХОЛЕСТЕРИНА
Субстрат для синтеза холестерола- ацетил-КоА
Активация синтеза холестерина происходит при поступлении в
Слайд 22СИНТЕЗ ХОЛЕСТЕРИНА
ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната
СИНТЕЗ ХОЛЕСТЕРИНА
ОСНОВНЫЕ СТАДИИ СИНТЕЗА ХОЛЕСТЕРИНА:
• синтез мевалоната (С6)
• синтез сквалена из мевалоната

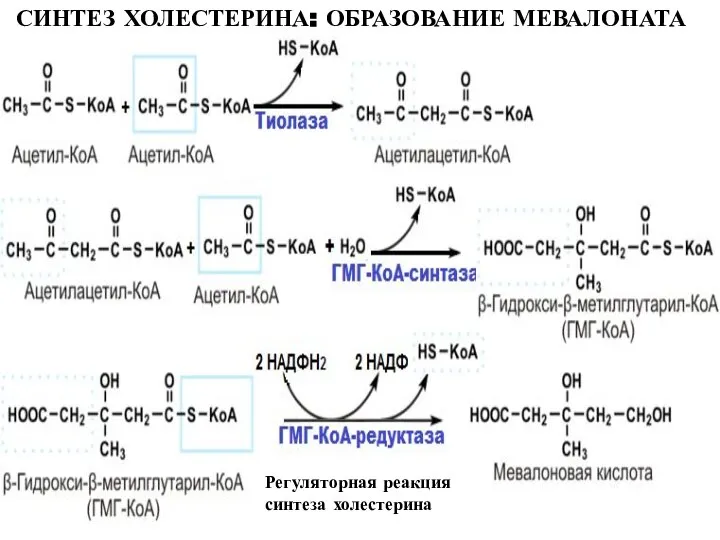
Слайд 23СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА
Регуляторная реакция синтеза холестерина
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ МЕВАЛОНАТА
Регуляторная реакция синтеза холестерина
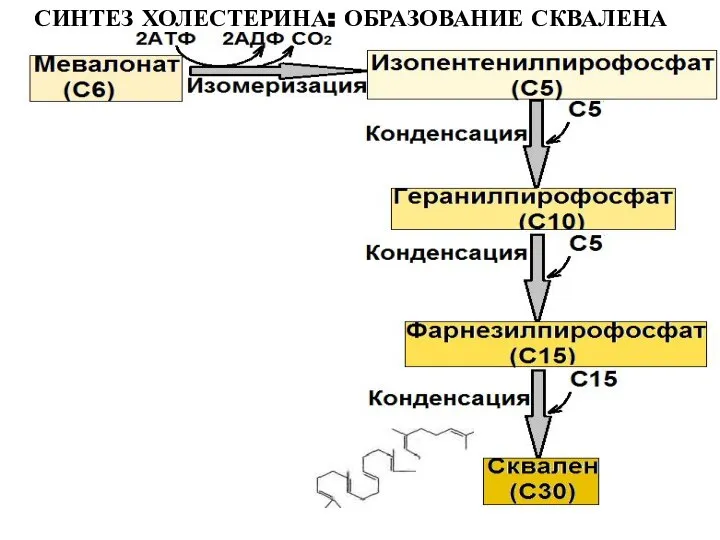
Слайд 24СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
СИНТЕЗ ХОЛЕСТЕРИНА: ОБРАЗОВАНИЕ СКВАЛЕНА
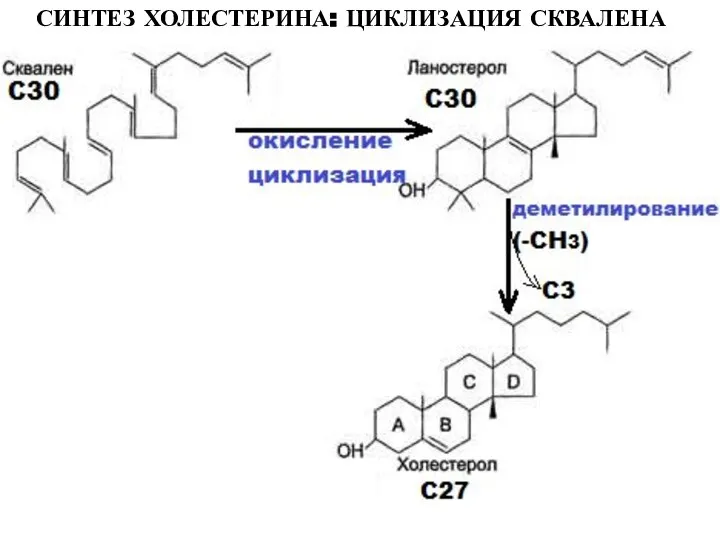
Слайд 25СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
СИНТЕЗ ХОЛЕСТЕРИНА: ЦИКЛИЗАЦИЯ СКВАЛЕНА
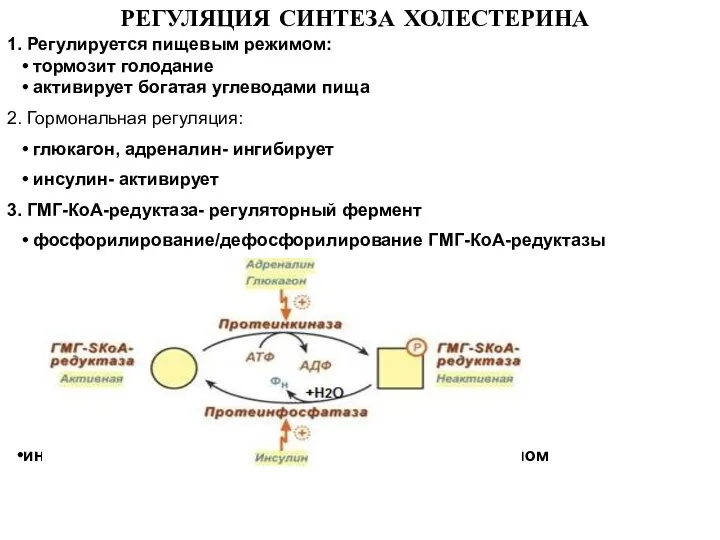
Слайд 26 РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА
1. Регулируется пищевым режимом:
• тормозит голодание
•
РЕГУЛЯЦИЯ СИНТЕЗА ХОЛЕСТЕРИНА
1. Регулируется пищевым режимом:
• тормозит голодание
•
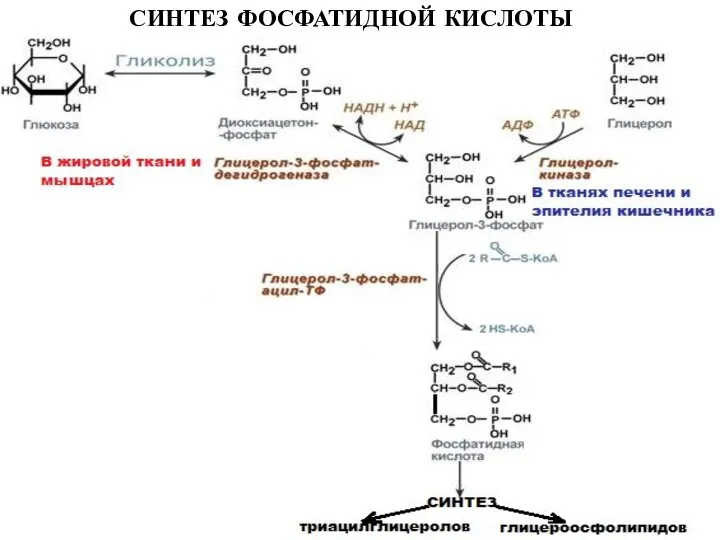
Слайд 27СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
СИНТЕЗ ФОСФАТИДНОЙ КИСЛОТЫ
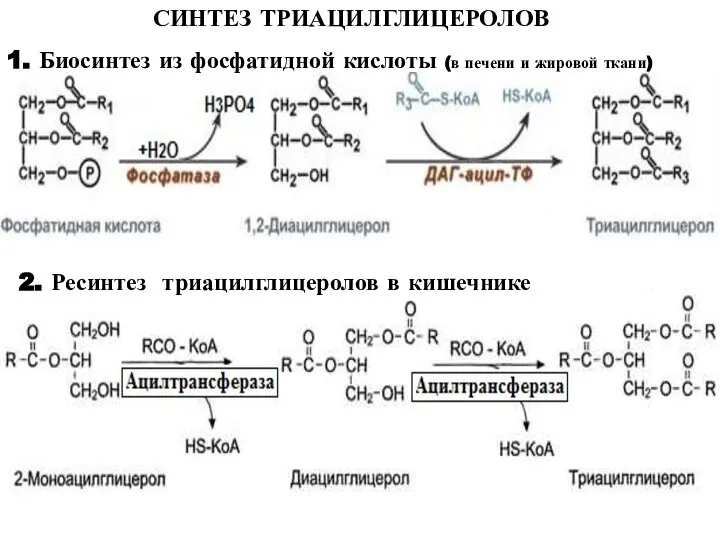
Слайд 28СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ
1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани)
2. Ресинтез
СИНТЕЗ ТРИАЦИЛГЛИЦЕРОЛОВ
1. Биосинтез из фосфатидной кислоты (в печени и жировой ткани)
2. Ресинтез
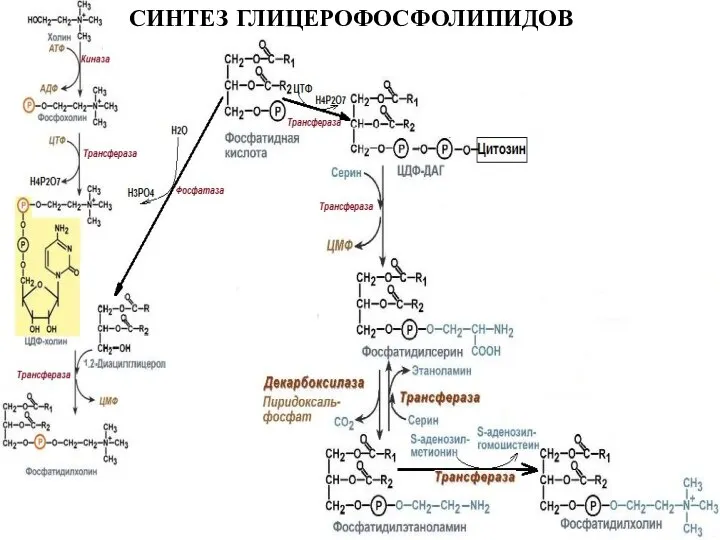
Слайд 29СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
СИНТЕЗ ГЛИЦЕРОФОСФОЛИПИДОВ
 Митоз
Митоз Человек и его здоровье
Человек и его здоровье Шестое чувство. Игра
Шестое чувство. Игра Нервная система человека
Нервная система человека Морфофункциональная характеристика слуховой, вестибулярной, соматической, проприоцептивной сенсорных систем
Морфофункциональная характеристика слуховой, вестибулярной, соматической, проприоцептивной сенсорных систем Строение синапса
Строение синапса Бактериофаги и их применение в вирусологии и биотехнологии
Бактериофаги и их применение в вирусологии и биотехнологии Опорная система живых органихмов
Опорная система живых органихмов Плауновидные растения
Плауновидные растения Возникновение и развитие эволюционной биологии
Возникновение и развитие эволюционной биологии Круглые черви
Круглые черви Деятельность человека
Деятельность человека Перелетные птицы. Экологическая акция “Поможем птицам”
Перелетные птицы. Экологическая акция “Поможем птицам” Составление сравнительных характеристик. Выявление сходства и различия между организмами и их частями
Составление сравнительных характеристик. Выявление сходства и различия между организмами и их частями Генная инженерия как технология
Генная инженерия как технология В подводном царстве, в загадочном государстве. Всемирный день китов и дельфинов
В подводном царстве, в загадочном государстве. Всемирный день китов и дельфинов Микробиология как наука, предмет ее изучения, исторические этапы развития, задачи медицинской микробиологии
Микробиология как наука, предмет ее изучения, исторические этапы развития, задачи медицинской микробиологии Цветы фиалки
Цветы фиалки Наследование признаков у человека. Наследственные болезни человека, их причины и профилактика. Практическая работа №2
Наследование признаков у человека. Наследственные болезни человека, их причины и профилактика. Практическая работа №2 Презентация на тему Клонирование
Презентация на тему Клонирование  Закономерности наследственности
Закономерности наследственности Иллюстрированный словарь народных названий растений Урала
Иллюстрированный словарь народных названий растений Урала Презентация на тему Лекарственные растения Викторина
Презентация на тему Лекарственные растения Викторина  Анатомия спинного мозга
Анатомия спинного мозга Орган зрения
Орган зрения Биополимеры: липиды. Их роль в жизнедеятельности клетки
Биополимеры: липиды. Их роль в жизнедеятельности клетки Запуск аквариума. Инструкция для начинающих
Запуск аквариума. Инструкция для начинающих Мир животных в природе
Мир животных в природе