- Обмен веществ

Содержание
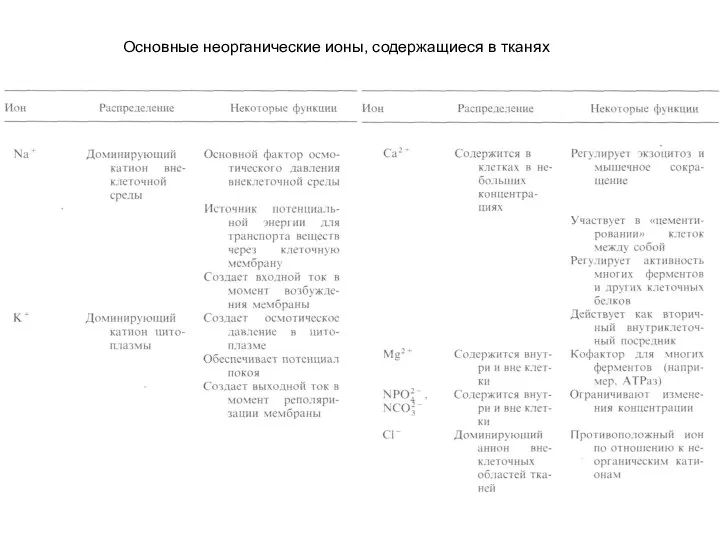
- 2. Основные неорганические ионы, содержащиеся в тканях
- 3. Состав внеклеточной жидкости у различных животных (концентрация дана в моль*д-1) часть 1 (Schmidt-Nielsen, Mackay, 1972; Prosser,
- 4. Состав внеклеточной жидкости у различных животных (концентрация дана в моль*д-1) часть 2
- 5. Состав электролитов в жидких средах человеческого организма Примечание. Некоторые ионы, содержащиеся внутри клетки, не распределены свободным
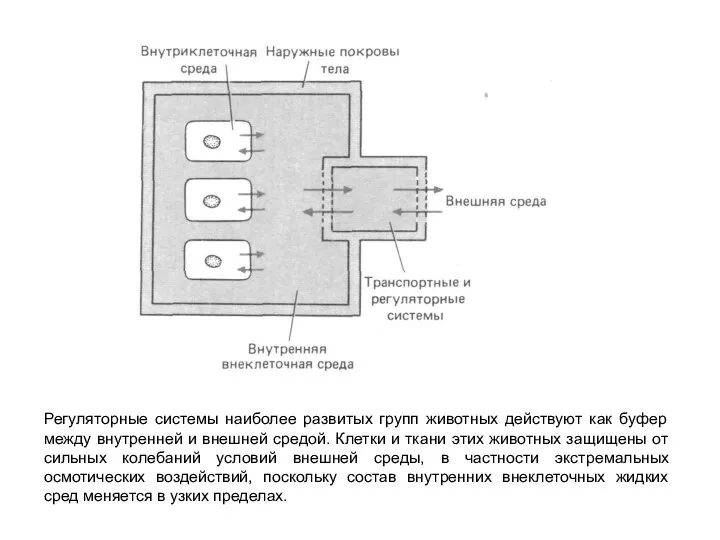
- 6. Регуляторные системы наиболее развитых групп животных действуют как буфер между внутренней и внешней средой. Клетки и
- 7. У животных - строгих осморегуляторов - концентрация соли и воды внутри организма поддерживается относительно постоянной несмотря
- 9. Контроль содержания глюкозы в плазме инсулином и глюкагоном
- 10. Вызванное глюкозой высвобождение инсулина из панкреатических В-клеток Инсулин стимулирует перемещение переносчиков глюкозы к мембране клеток-мишеней
- 11. Терморегуляция
- 12. Различие в процессе нагревания и охлаждения у галапагосской морской игуаны. Греясь на суше, игуана поглощает солнечное
- 13. Различие в процессе нагревания и охлаждения у галапагосской морской игуаны. Б. Разница между температурой тела и
- 14. Регуляция температуры у неподвижной бабочки-бражника Manduca sexta во время 25-минутного подведения внешнего тепла к груди (цветная
- 15. Схема кровоснабжения «холодной» рыбы (А) и типичной «теплой» рыбы-голубого тунца Tunnus thynnus (Б). У эктотермной рыбы
- 16. Респираторный калориметр Этуотера-Розы Животное помещено в изолированную камеру, в которой поддерживается пос- тоянная температура. Теплопродукцию животного
- 17. Каротидная сеть, обнаруженная у некоторых млекопитающих (обозначена в цвете). Система мелких артерий действует как теплообменник на
- 18. Кaротидная сеть и противоточное охлаждение крови из сонной артерии у овцы. Холодная венозная кровь, возвращаясь из
- 19. Изменения напряжения кислорода и углекислого газа в процессе газообмена между воздухом (А) или водой (Б) и
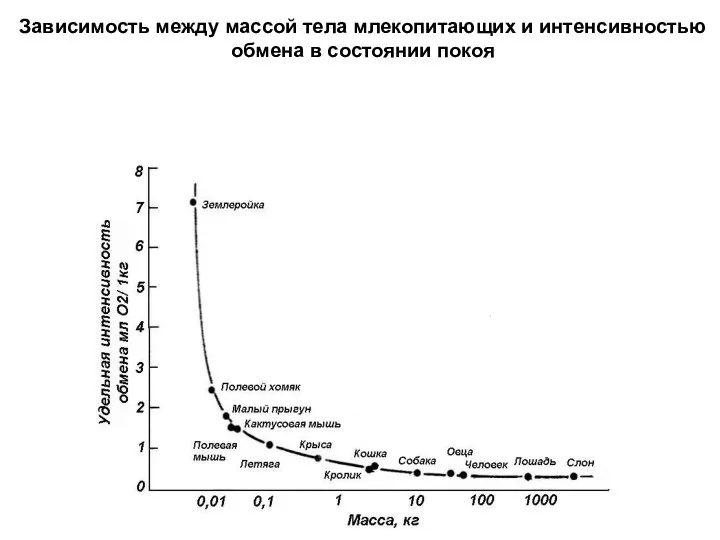
- 21. Зависимость между массой тела млекопитающих и интенсивностью обмена в состоянии покоя
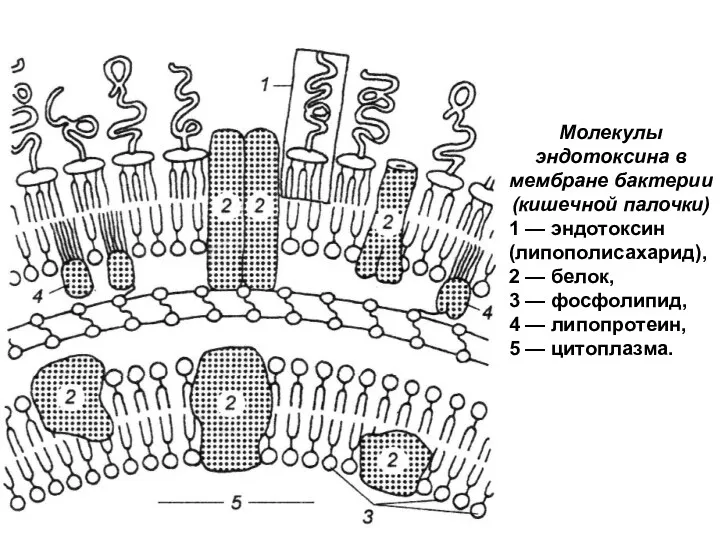
- 23. Молекулы эндотоксина в мембране бактерии (кишечной палочки) 1 — эндотоксин (липополисахарид), 2 — белок, 3 —
- 25. Скачать презентацию
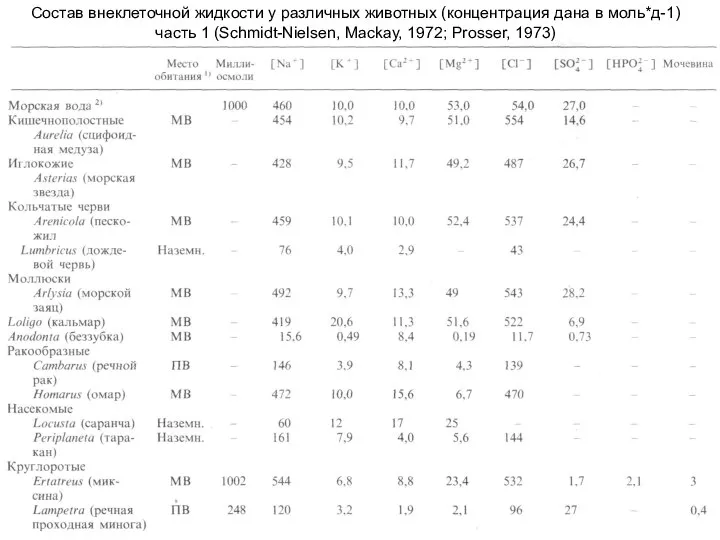
Слайд 3Состав внеклеточной жидкости у различных животных (концентрация дана в моль*д-1)
часть 1 (Schmidt-Nielsen,
Состав внеклеточной жидкости у различных животных (концентрация дана в моль*д-1)
часть 1 (Schmidt-Nielsen,
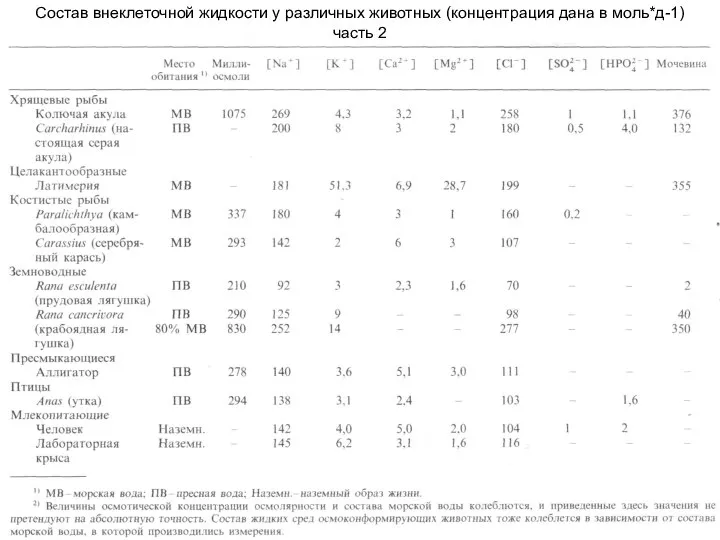
Слайд 4Состав внеклеточной жидкости у различных животных (концентрация дана в моль*д-1)
часть 2
Состав внеклеточной жидкости у различных животных (концентрация дана в моль*д-1)
часть 2
Слайд 5Состав электролитов в жидких средах человеческого организма
Примечание. Некоторые ионы, содержащиеся внутри клетки,
Состав электролитов в жидких средах человеческого организма
Примечание. Некоторые ионы, содержащиеся внутри клетки,

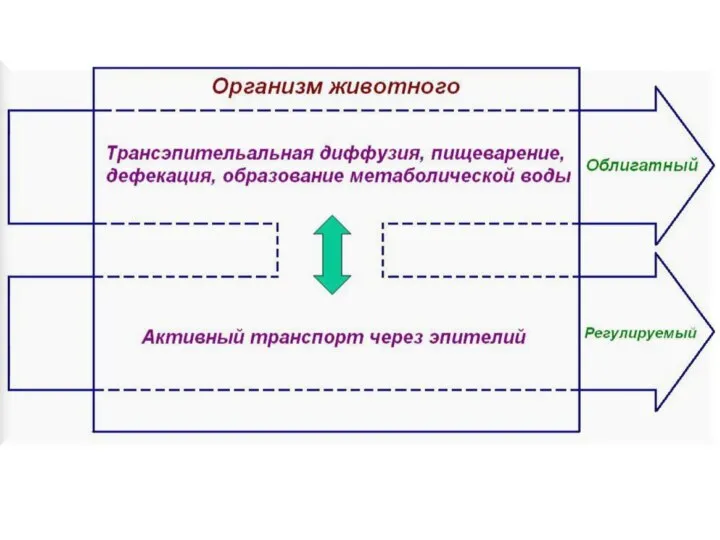
Слайд 6Регуляторные системы наиболее развитых групп животных действуют как буфер между внутренней и
Регуляторные системы наиболее развитых групп животных действуют как буфер между внутренней и
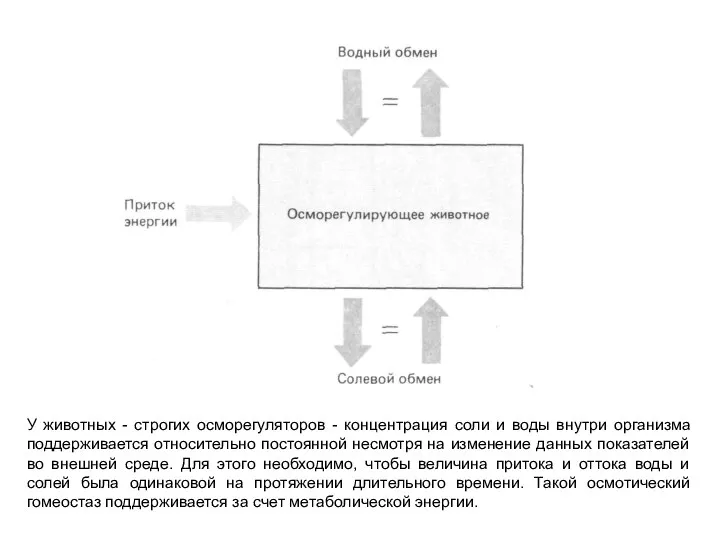
Слайд 7У животных - строгих осморегуляторов - концентрация соли и воды внутри организма
У животных - строгих осморегуляторов - концентрация соли и воды внутри организма
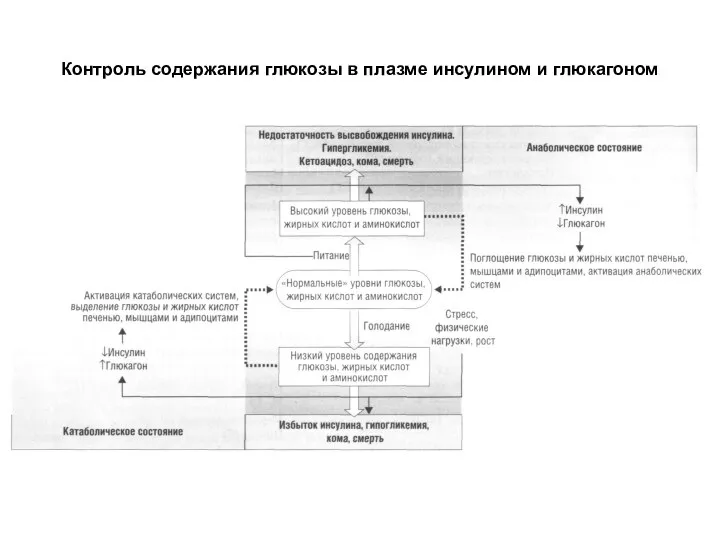
Слайд 9Контроль содержания глюкозы в плазме инсулином и глюкагоном
Контроль содержания глюкозы в плазме инсулином и глюкагоном
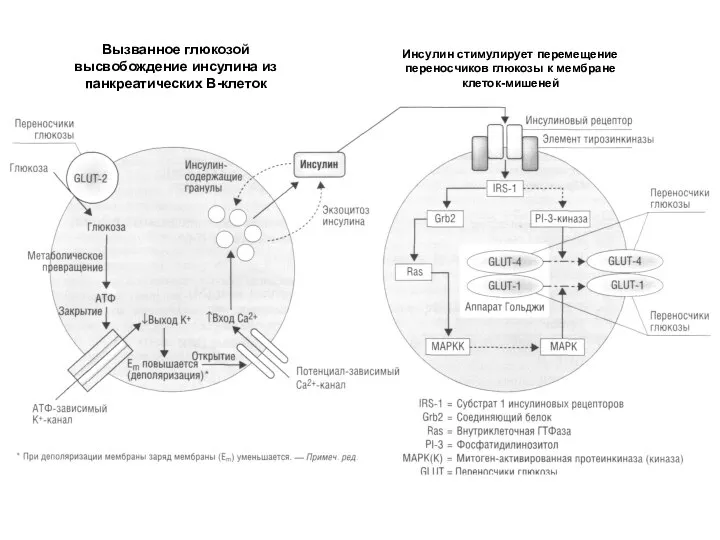
Слайд 10Вызванное глюкозой высвобождение инсулина из панкреатических В-клеток
Инсулин стимулирует перемещение переносчиков глюкозы к
Вызванное глюкозой высвобождение инсулина из панкреатических В-клеток
Инсулин стимулирует перемещение переносчиков глюкозы к
Слайд 11Терморегуляция
Терморегуляция

Слайд 12Различие в процессе нагревания и охлаждения у галапагосской морской игуаны.
Греясь на суше,
Различие в процессе нагревания и охлаждения у галапагосской морской игуаны.
Греясь на суше,
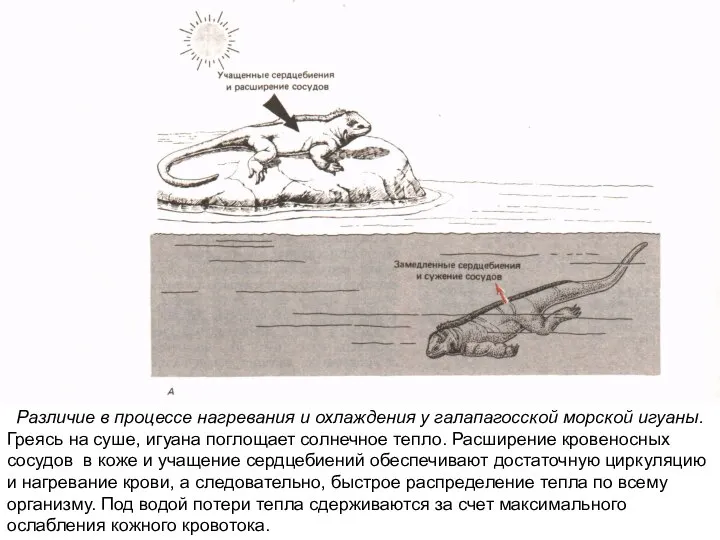
Слайд 13Различие в процессе нагревания и охлаждения у галапагосской морской игуаны.
Б. Разница
Различие в процессе нагревания и охлаждения у галапагосской морской игуаны.
Б. Разница
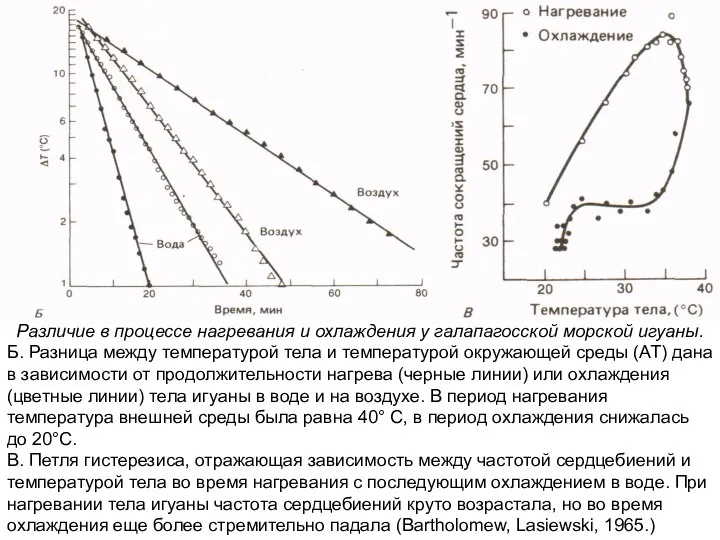
Слайд 14Регуляция температуры у неподвижной бабочки-бражника Manduca sexta во время 25-минутного подведения внешнего
Регуляция температуры у неподвижной бабочки-бражника Manduca sexta во время 25-минутного подведения внешнего
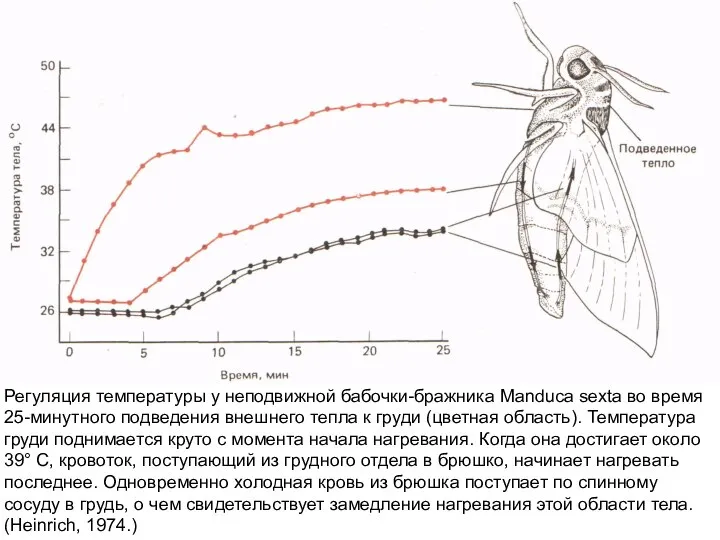
Слайд 15Схема кровоснабжения «холодной» рыбы (А) и типичной «теплой» рыбы-голубого тунца Tunnus thynnus
Схема кровоснабжения «холодной» рыбы (А) и типичной «теплой» рыбы-голубого тунца Tunnus thynnus
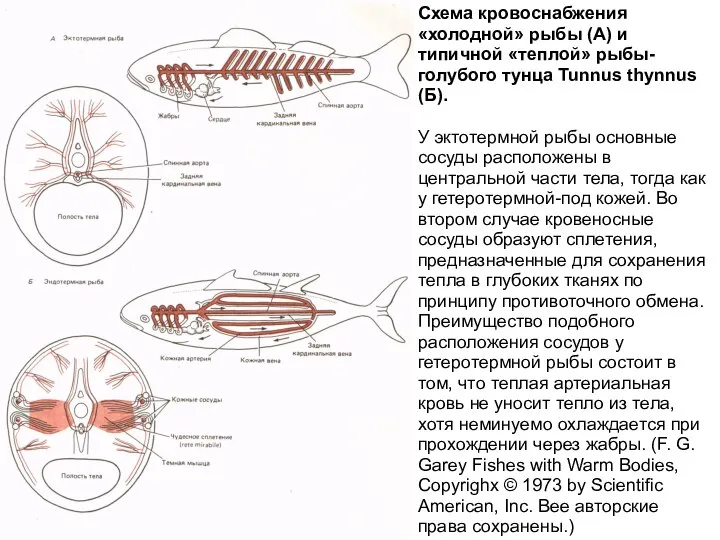
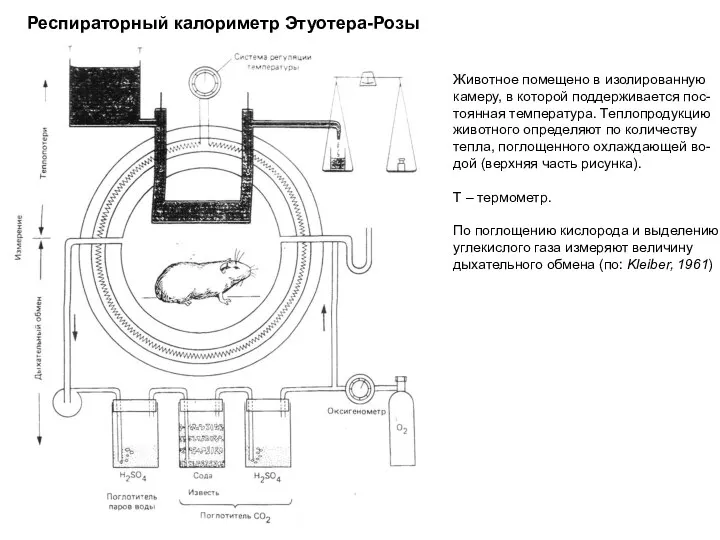
Слайд 16Респираторный калориметр Этуотера-Розы
Животное помещено в изолированную
камеру, в которой поддерживается пос-
тоянная температура. Теплопродукцию
животного
Респираторный калориметр Этуотера-Розы
Животное помещено в изолированную
камеру, в которой поддерживается пос-
тоянная температура. Теплопродукцию
животного
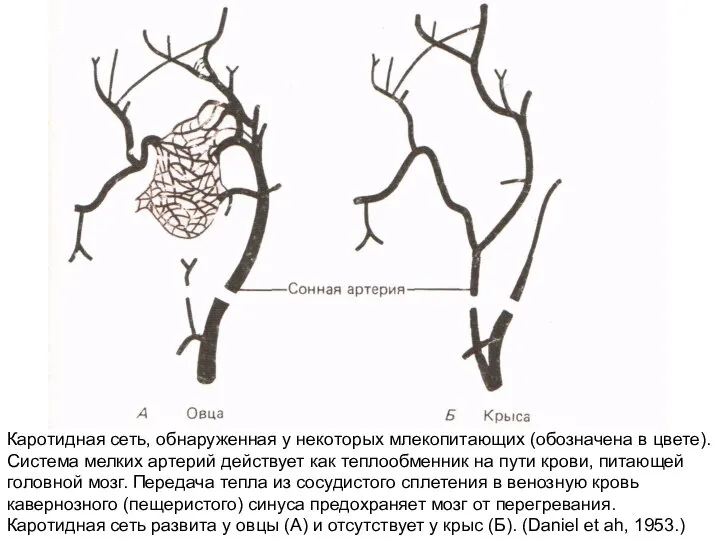
Слайд 17Каротидная сеть, обнаруженная у некоторых млекопитающих (обозначена в цвете). Система мелких артерий
Каротидная сеть, обнаруженная у некоторых млекопитающих (обозначена в цвете). Система мелких артерий
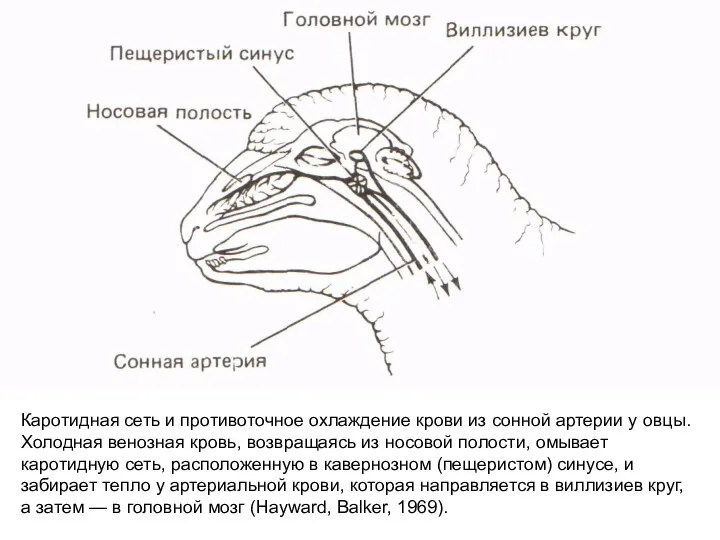
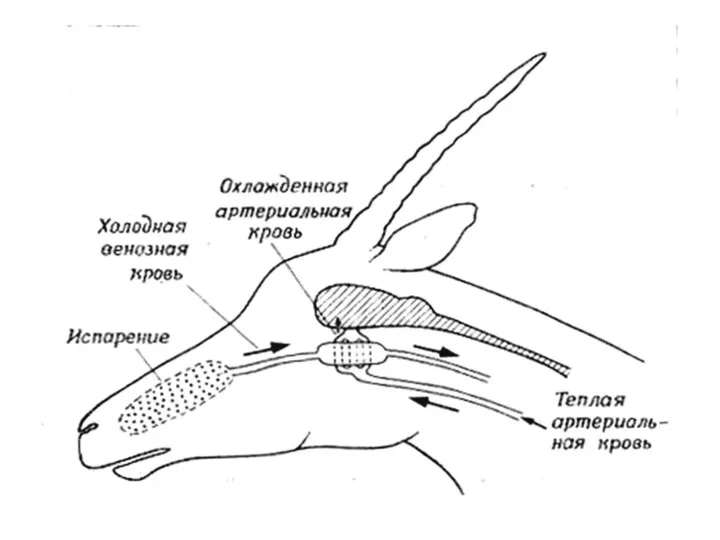
Слайд 18Кaротидная сеть и противоточное охлаждение крови из сонной артерии у овцы. Холодная
Кaротидная сеть и противоточное охлаждение крови из сонной артерии у овцы. Холодная
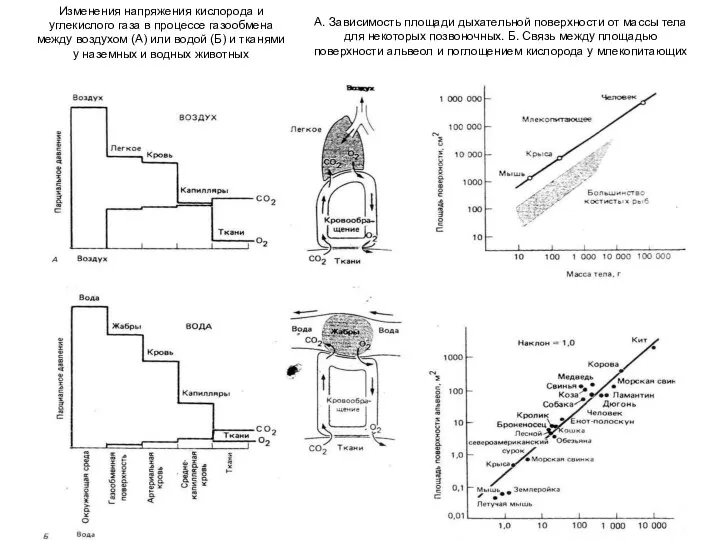
Слайд 19Изменения напряжения кислорода и
углекислого газа в процессе газообмена
между воздухом (А)
Изменения напряжения кислорода и
углекислого газа в процессе газообмена
между воздухом (А)
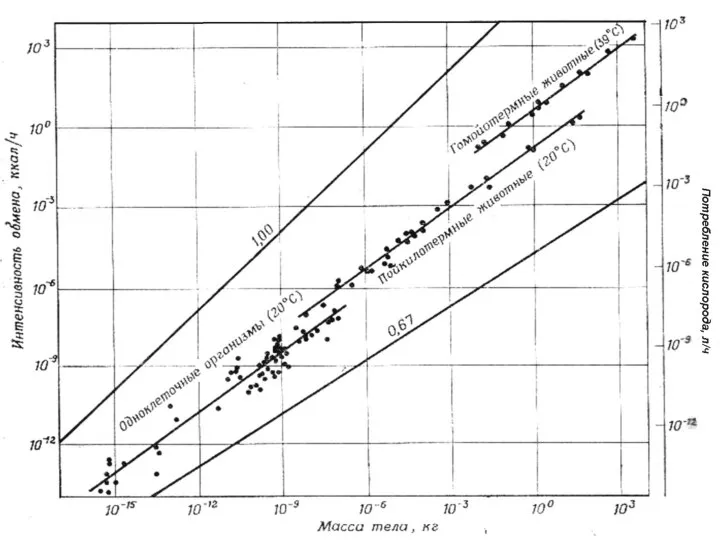
Слайд 21Зависимость между массой тела млекопитающих и интенсивностью
обмена в состоянии покоя
Зависимость между массой тела млекопитающих и интенсивностью
обмена в состоянии покоя
Слайд 23Молекулы эндотоксина в мембране бактерии (кишечной палочки)
1 — эндотоксин (липополисахарид),
2 —
Молекулы эндотоксина в мембране бактерии (кишечной палочки)
1 — эндотоксин (липополисахарид),
2 —
 Насекомые вредители на станции юных натуралистов г. Тайшета
Насекомые вредители на станции юных натуралистов г. Тайшета Рыбы и насекомые пресных водоемов
Рыбы и насекомые пресных водоемов Грибы, цветы и ягоды леса
Грибы, цветы и ягоды леса Чей хвостик
Чей хвостик Семейство крестоцветных
Семейство крестоцветных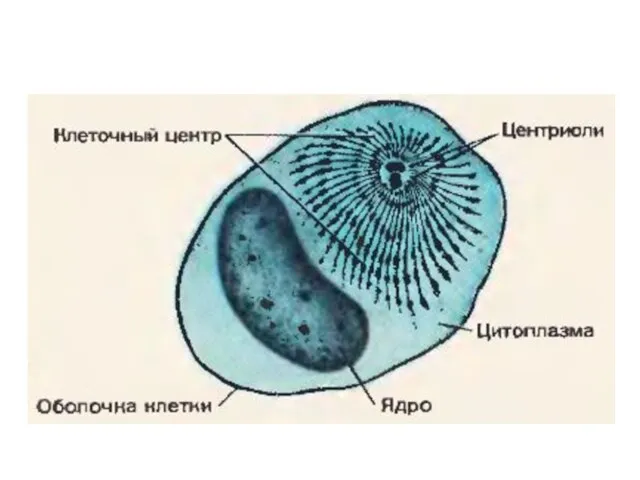 Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба
Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба Неклеточные формы жизни
Неклеточные формы жизни Раселение живых организмов
Раселение живых организмов Эпителий
Эпителий Презентация на тему Строение и химический состав клетки
Презентация на тему Строение и химический состав клетки  MOLEKUlyarnaya_biologia_LPZ_1
MOLEKUlyarnaya_biologia_LPZ_1 Фотодыхание
Фотодыхание Вирусология - наука о вирусах
Вирусология - наука о вирусах Кровь (физиология системы крови) - Презентация
Кровь (физиология системы крови) - Презентация Чабрец (богородская трава, тимьян ползучий)
Чабрец (богородская трава, тимьян ползучий) Составление генетических паспортов и установление родственных связей
Составление генетических паспортов и установление родственных связей От клюва до хвоста
От клюва до хвоста Модификационная изменчивость
Модификационная изменчивость Строение и многообразие грибов
Строение и многообразие грибов Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности
Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности  Что было на Земле, когда она появилась?
Что было на Земле, когда она появилась? Биохимия. Разделы биохимии. Становление биохимии как науки
Биохимия. Разделы биохимии. Становление биохимии как науки История собаководства
История собаководства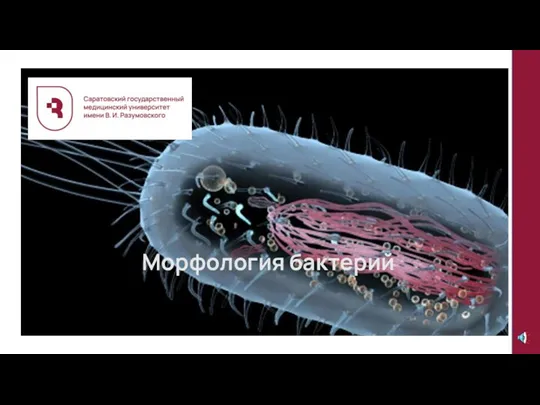 Морфология бактерий
Морфология бактерий Жизненная Ёмкость Лёгких
Жизненная Ёмкость Лёгких Ткани животных
Ткани животных 2. Урок - .ДНК
2. Урок - .ДНК Лишайники
Лишайники