- Обонятельная рецепция

Содержание
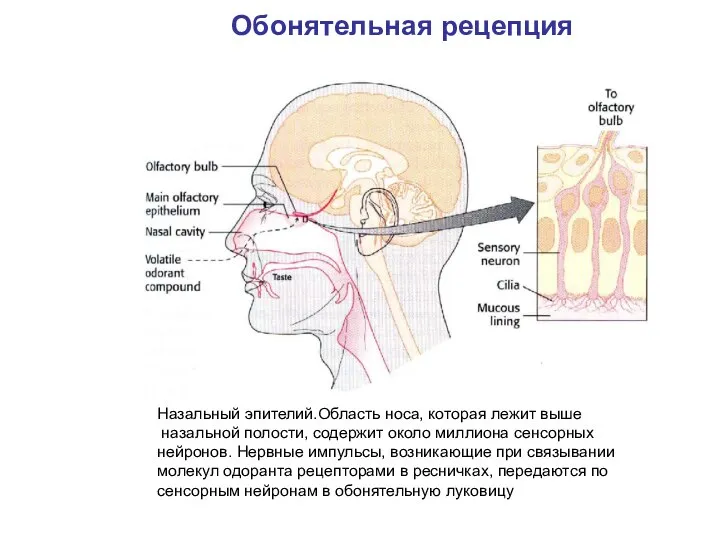
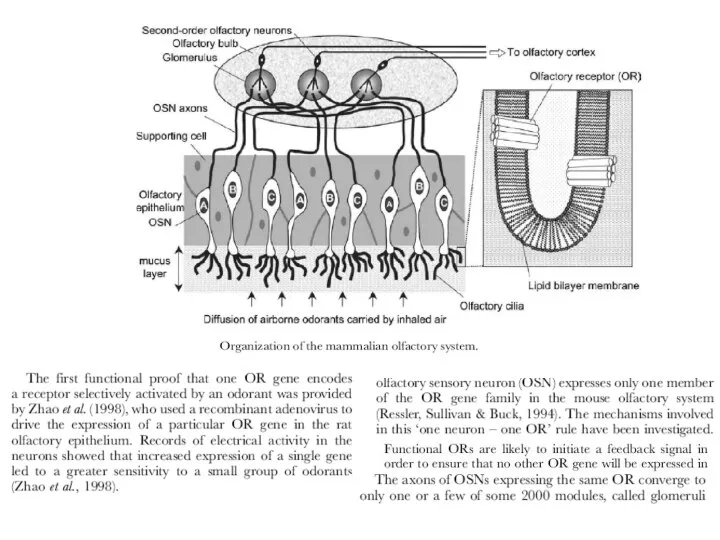
- 2. Назальный эпителий.Область носа, которая лежит выше назальной полости, содержит около миллиона сенсорных нейронов. Нервные импульсы, возникающие
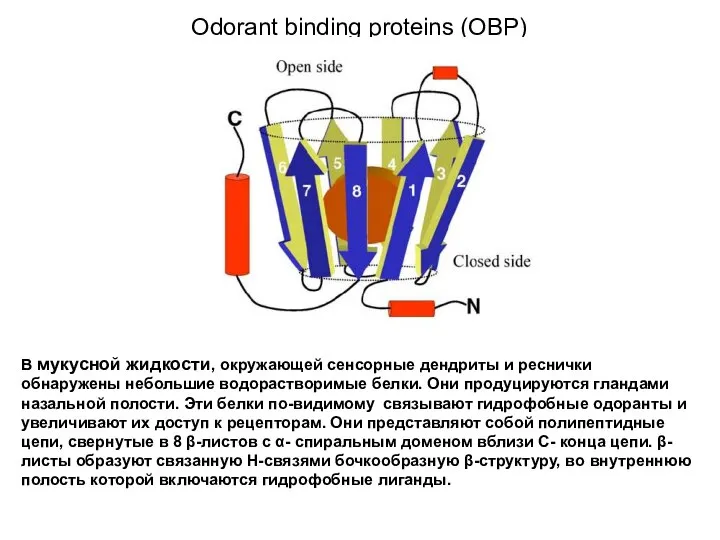
- 3. Odorant binding proteins (OBP) В мукусной жидкости, окружающей сенсорные дендриты и реснички обнаружены небольшие водорастворимые белки.
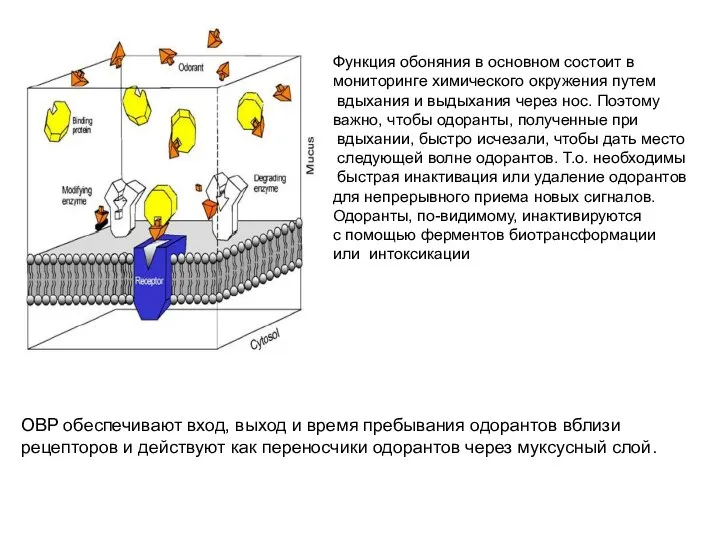
- 4. OBP обеспечивают вход, выход и время пребывания одорантов вблизи рецепторов и действуют как переносчики одорантов через
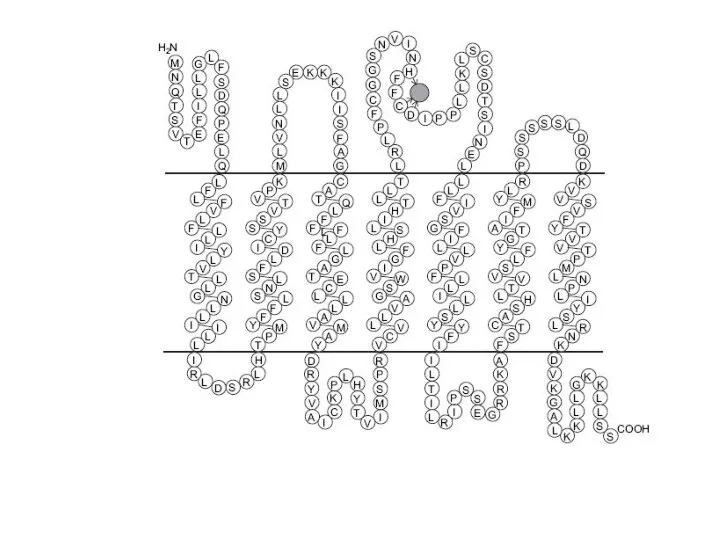
- 5. Обоняние обеспечивается за счет громадного семейства рецепторов, которые представляют собой трансмембранные белки, состоящие из семи α-спиралей,
- 7. Связывание одоранта с ОР на нейрональной поверхности инициирует каскад трансдукции сигнала, который приводит к созданию потенциала
- 8. Ощущение запаха: молекулярные основы узнавания одоранта Действие химических функциональных групп Давно замечено, что присутствие определенных химических
- 9. 2.1. Теория обоняния за счет profile-functional groups. Химики давно заметили, что присутствие в молекуле определенных химических
- 10. 2.2. Вибрационная теория обоняния. Спектроскопические исследования Дайсона в 1930 г. обнаружили корреляцию между определенными запахами и
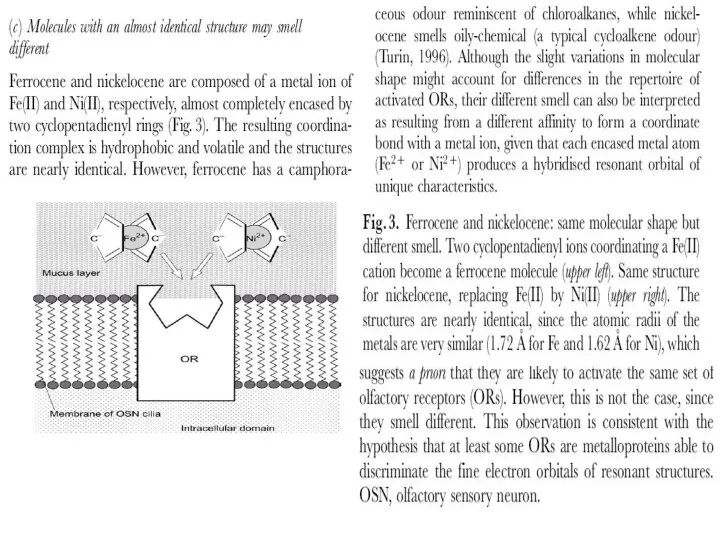
- 12. 2.3. Стерическая теория обоняния. После открытия фермент-субстратных взаимодействий, основанных на молекулярном узнавании между белковым рецептором и
- 26. Isotope effects
- 27. Резкая граница по размерам молекул при их детектировании Не все молекулы пахнут. Во первых, требуется определенная
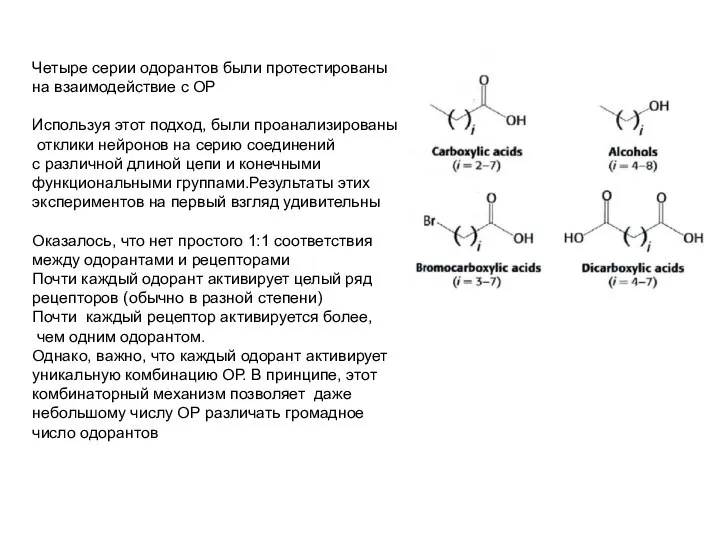
- 28. Одоранты декодируются по комбинаторному механизму Проблема обонятельной рецепции - как сопоставить каждый обонятельный рецептор (ОР) из
- 29. Четыре серии одорантов были протестированы на взаимодействие с ОР Используя этот подход, были проанализированы отклики нейронов
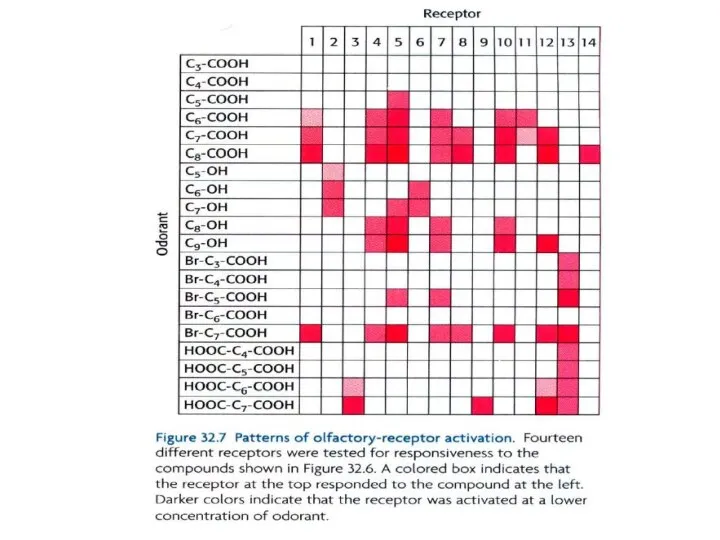
- 31. Как информация о том, какой ОР активировался данным одорантом, передается в мозг? Нужно вспомнить, что каждый
- 32. Может ли такой комбинаторный механизм реально различать много различных одорантов? Электронный нос, который функционирует по тому
- 35. CONCLUSIONS Обоняние – это химическое восприятие, которое производит ощущение и дискриминацию летучих химических веществ. Оно играет
- 36. ОР принадлежат к семейству рецепторов, действующих через G-белки, которое охватывает большое число рецепторов, реагирующих на разнообразные
- 38. Скачать презентацию
Слайд 2Назальный эпителий.Область носа, которая лежит выше
назальной полости, содержит около миллиона сенсорных
нейронов.
Назальный эпителий.Область носа, которая лежит выше
назальной полости, содержит около миллиона сенсорных
нейронов.
Слайд 3Odorant binding proteins (OBP)
В мукусной жидкости, окружающей сенсорные дендриты и реснички обнаружены
Odorant binding proteins (OBP)
В мукусной жидкости, окружающей сенсорные дендриты и реснички обнаружены
Слайд 4OBP обеспечивают вход, выход и время пребывания одорантов вблизи рецепторов и действуют
Слайд 5Обоняние обеспечивается за счет громадного семейства рецепторов, которые
представляют собой трансмембранные белки,
Обоняние обеспечивается за счет громадного семейства рецепторов, которые
представляют собой трансмембранные белки,

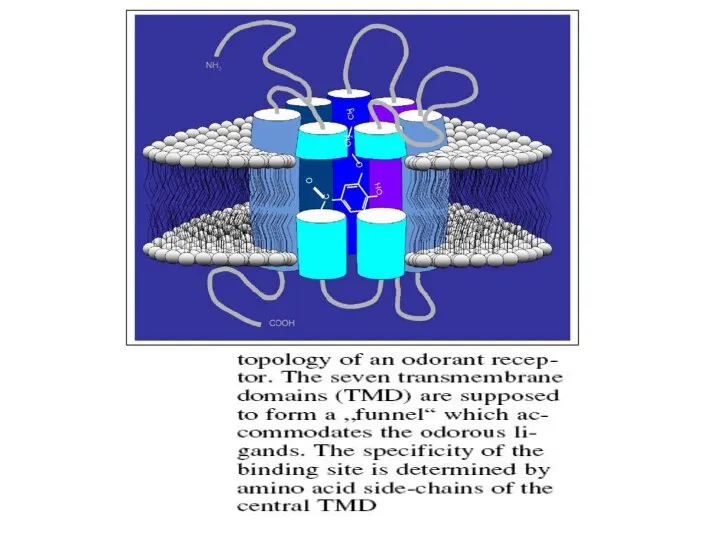
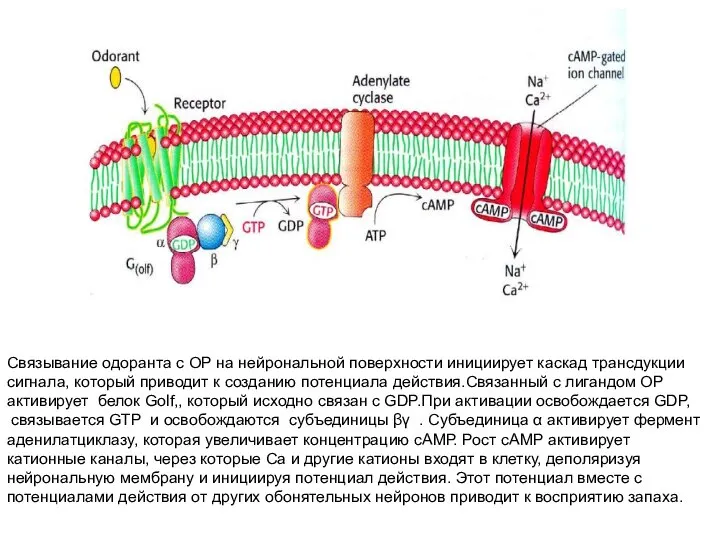
Слайд 7Связывание одоранта с ОР на нейрональной поверхности инициирует каскад трансдукции
сигнала, который
Связывание одоранта с ОР на нейрональной поверхности инициирует каскад трансдукции
сигнала, который
Слайд 8Ощущение запаха: молекулярные основы узнавания одоранта
Действие химических
функциональных групп
Давно замечено, что присутствие
Ощущение запаха: молекулярные основы узнавания одоранта
Действие химических
функциональных групп
Давно замечено, что присутствие

Слайд 92.1. Теория обоняния за счет profile-functional groups.
Химики давно заметили, что присутствие в
2.1. Теория обоняния за счет profile-functional groups.
Химики давно заметили, что присутствие в

Слайд 102.2. Вибрационная теория обоняния.
Спектроскопические исследования Дайсона в 1930 г. обнаружили корреляцию
между
2.2. Вибрационная теория обоняния.
Спектроскопические исследования Дайсона в 1930 г. обнаружили корреляцию
между


Слайд 122.3. Стерическая теория обоняния.
После открытия фермент-субстратных взаимодействий, основанных на молекулярном
узнавании между белковым
2.3. Стерическая теория обоняния.
После открытия фермент-субстратных взаимодействий, основанных на молекулярном
узнавании между белковым











Слайд 26Isotope effects
Isotope effects

Слайд 27Резкая граница по размерам молекул при их детектировании
Не все молекулы пахнут. Во
Резкая граница по размерам молекул при их детектировании
Не все молекулы пахнут. Во

Слайд 28Одоранты декодируются по комбинаторному механизму
Проблема обонятельной рецепции - как сопоставить каждый обонятельный
Одоранты декодируются по комбинаторному механизму
Проблема обонятельной рецепции - как сопоставить каждый обонятельный

Слайд 29Четыре серии одорантов были протестированы
на взаимодействие с ОР
Используя этот подход, были проанализированы
Четыре серии одорантов были протестированы
на взаимодействие с ОР
Используя этот подход, были проанализированы
Слайд 31Как информация о том, какой ОР активировался данным одорантом, передается
в мозг? Нужно
Как информация о том, какой ОР активировался данным одорантом, передается
в мозг? Нужно

Слайд 32Может ли такой комбинаторный механизм реально различать много различных
одорантов? Электронный нос,
Может ли такой комбинаторный механизм реально различать много различных
одорантов? Электронный нос,


Слайд 35CONCLUSIONS
Обоняние – это химическое восприятие, которое производит ощущение и
дискриминацию летучих химических
CONCLUSIONS
Обоняние – это химическое восприятие, которое производит ощущение и
дискриминацию летучих химических

Слайд 36ОР принадлежат к семейству рецепторов, действующих через G-белки, которое
охватывает большое число
ОР принадлежат к семейству рецепторов, действующих через G-белки, которое
охватывает большое число

 Семена. Сортовой контроль
Семена. Сортовой контроль Центральная нервная система
Центральная нервная система Строение головного мозга
Строение головного мозга Зимующие птицы
Зимующие птицы Сенсорные системы (анализаторы) в обеспечении защиты организма от опасных и вредных факторов окружающей среды
Сенсорные системы (анализаторы) в обеспечении защиты организма от опасных и вредных факторов окружающей среды Что такое удобрения
Что такое удобрения Трансляция генетической информации
Трансляция генетической информации Органы пищеварения
Органы пищеварения Кошки
Кошки Строение кости. Типы соединения костей
Строение кости. Типы соединения костей Биология в средневековье. (Лекция 3)
Биология в средневековье. (Лекция 3) Жизнь пчелиной семьи
Жизнь пчелиной семьи Путешествие по страницам Красной книги
Путешествие по страницам Красной книги Свойства аминокислот (3)
Свойства аминокислот (3) Презентация на тему ПОЧЕМУ ЛИСТЬЯ ЗЕЛЁНЫЕ ИЛИ РОЛЬ ЗЕЛЁНЫХ ЛИСТЬЕВ ДЛЯ РАСТЕНИЙ И ЧЕЛОВЕКА
Презентация на тему ПОЧЕМУ ЛИСТЬЯ ЗЕЛЁНЫЕ ИЛИ РОЛЬ ЗЕЛЁНЫХ ЛИСТЬЕВ ДЛЯ РАСТЕНИЙ И ЧЕЛОВЕКА  Ядра и капсулы конечного мозга
Ядра и капсулы конечного мозга Надкласс Рыбы. Класс Костные рыбы
Надкласс Рыбы. Класс Костные рыбы Сердечно-сосудистая система у домашних млекопитающих животных
Сердечно-сосудистая система у домашних млекопитающих животных Распространение плодов и семян
Распространение плодов и семян Игра - путешествие В лес по загадки
Игра - путешествие В лес по загадки Ткани растений
Ткани растений Презентация на тему Что такое кровь (4 класс)
Презентация на тему Что такое кровь (4 класс)  Сравнение процессов брожения и дыхания в клетках
Сравнение процессов брожения и дыхания в клетках Земноводные и пресмыкающиеся. Кто это?
Земноводные и пресмыкающиеся. Кто это? Методы регистрации клеточного цикла
Методы регистрации клеточного цикла Сердечно-сосудистая система
Сердечно-сосудистая система Виды плодов
Виды плодов Микрофлора тела здорового человека
Микрофлора тела здорового человека