- Общая концепция организации движений. Двигательная система

Содержание
- 2. Роль спинного мозга в процессах регуляции деятельности опорно-двигательного аппарата и вегетативных функций организма. Мышечные веретена, эфферентная
- 3. Целевые функции двигательной системы
- 4. Функции опорно-двигательного аппарата и системы его регуляции: поддержание равновесия (позная/статическая деятельность мышц) движения в процессе деятельности
- 5. Представление о пирамидной и экстрапирамидной системах
- 6. Структуры, отвечающие за регуляцию позы и движений, находятся в разных отделах ЦНС — от спинного мозга
- 7. Самый низший уровень в организации движения связан со спинным мозгом чувствительные, вставочные и мотонейроны спинальные рефлексы
- 8. Проекционные (двигательные) зоны коры головного мозга – двигательная кора (кпереди от центр.извилины) их раздражение приводит к
- 9. Двигательные пути, идущие от головного мозга к спинному, делятся на две системы пирамидную начало в моторной
- 10. Экстрапирамидная система – совокупность структур головного мозга, участвующих в управлении движениями, поддержании мышечного тонуса и позы,
- 11. Экстрапирамидная система эволюционно более древняя система моторного контроля значение в построении и контроле движений, не требующих
- 12. Пирамидная система (пирамидный путь ) одно из поздних приобретений эволюции нет у низших позвоночных появляется только
- 13. Кора полушарий головного мозга, V слой - гигантские пирамидные клетки (Беца)* нервные волокна – в спинной
- 14. Различают следующие типы волокон в пирамидной системе: Наибольшее количество пирамидных клеток (клеток Беца) иннервирует мелкие мышцы,
- 15. Повреждения пирамидной системы проявляются параличами, парезами, патологическими рефлексами. Поражение пирамидной системы может быть вызвано воспалением (энцефалит),
- 16. Общие принципы регуляции двигательной функции
- 17. Произвольные динамические движения мышц – целенаправленное действие: побуждение к действию (драйв, мотивация)→замысел действия→реализация замысла→регуляция позы при
- 18. Двигательная система - нейронные механизмы, участвующие в реализации фазной и позной мышечной активности Кора головного мозга
- 20. Общий план организации двигательной системы (ДС): 3 этажа (по Н.А. Агаджаняну, 2006) Спинной мозг – первый
- 21. Стволовые структуры мозга (вестибулярные ядра, красное ядро, ретикулярная формация, покрышка четверохолмия) – второй этаж управления ДС
- 22. Кора головного мозга – третий этаж управления ДС в ассоциативной коре – «замысел» движения → двигательная
- 23. Все звенья ДС используют сенсорную информацию от рецепторов мышц мышечные веретена, сухожильные рецепторы Гольджи рецепторы суставов
- 24. октябрь 2008 № 10 "В МИРЕ НАУКИ" Биология мозга Нейрофизиология танца Стивен Браун и Лоренс Парсонс
- 26. Контроль позы и движений
- 27. Схема контроля произвольных движений
- 28. Общий конечный путь к скелетным мышцам от мотонейронов спинного мозга и нейронов двигательных ядер черепных нервов
- 29. Мотонейроны спинного мозга и нейроны двигательных ядер черепных нервов — общий конечный путь к скелетным мышцам.
- 30. Роль спинного мозга в регуляций двигательной активности Примитивные процессы регуляции скелетных мышц фазные движения типа сгибания/разгибания
- 31. Роль ствола мозга в регуляции движений Продолговатый мозг, мост, средний мозг двигательные ядра черепно-мозговых нервов вестибулярные
- 32. «Базальные ядра» хвостатое ядро, скорлупа, субталамическое (подбугорное) ядро, бледный шар и чёрная субстанция Координация двигательных функций:
- 33. (Fredrik Ullén, доцент Королевского Каролинского института и Стокгольмского института мозга, 2010). сложные движения (одевание, игра на
- 34. Патология базальных ядер Гипер/гипокинетические состояния Хорея (от греч. choreia — хоровод, пляска) Атетоз (от греч. athetos
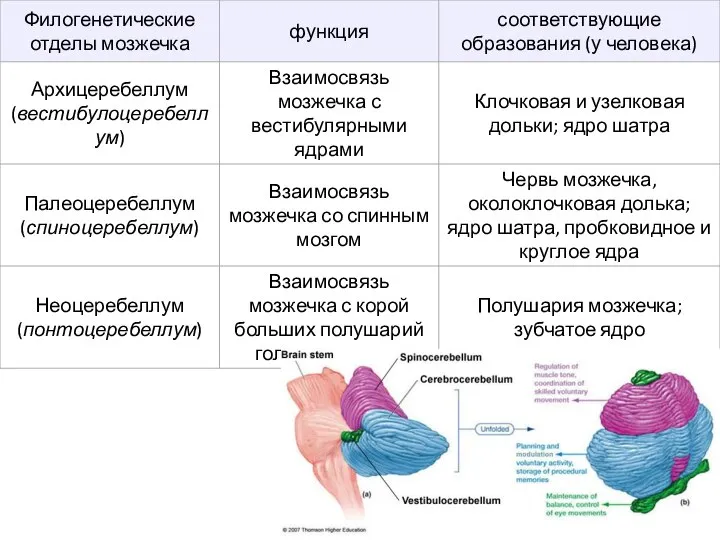
- 35. Мозжечок в системе двигательного контроля Три уровня участия в системе контроля и координации движений: Архиоцеребеллум (древний
- 37. Мозжечковые симптомы определяются местом повреждения Дисметрия — неспособность правильной оценки расстояния → неспособность сразу взять необходимый
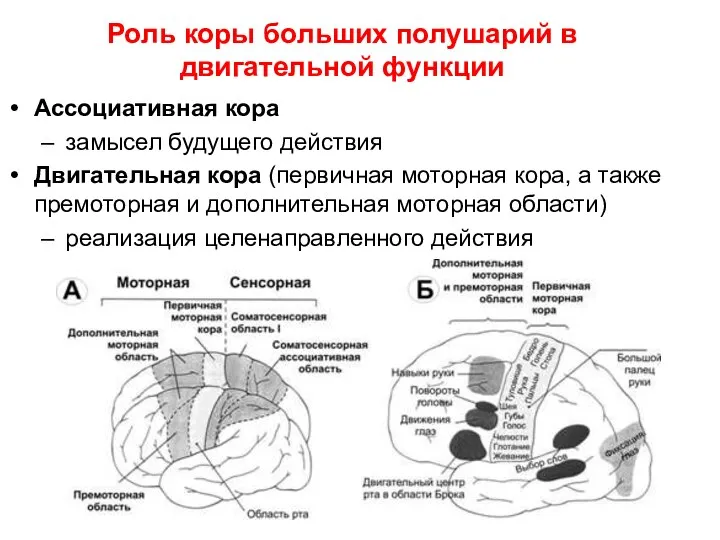
- 38. Роль коры больших полушарий в двигательной функции Ассоциативная кора замысел будущего действия Двигательная кора (первичная моторная
- 39. Первичная моторная кора картированы участки, ответственные за сокращения отдельных мышц, но чаще возбуждаются мышечные группы, далее
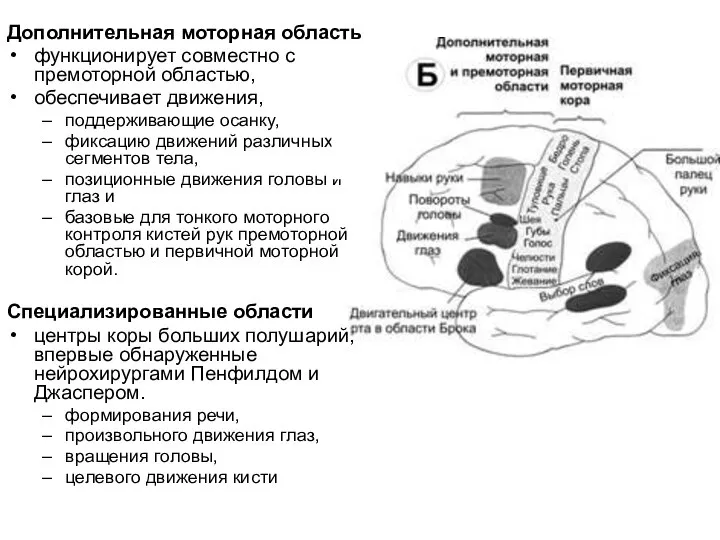
- 40. Дополнительная моторная область функционирует совместно с премоторной областью, обеспечивает движения, поддерживающие осанку, фиксацию движений различных сегментов
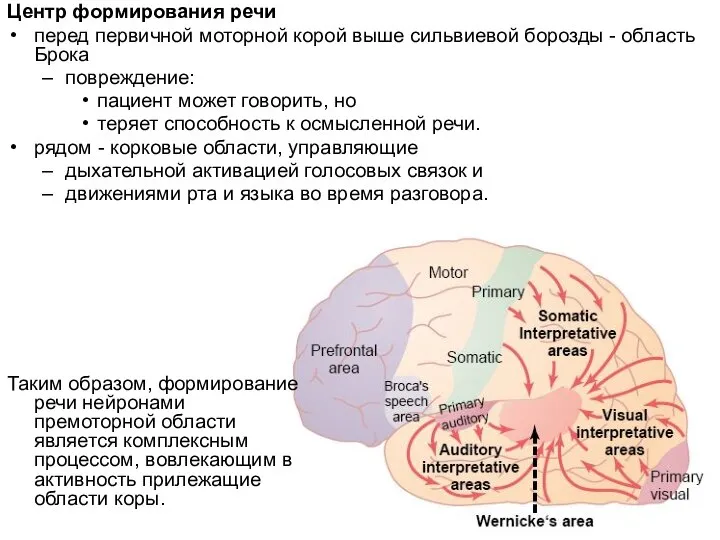
- 41. Центр формирования речи перед первичной моторной корой выше сильвиевой борозды - область Брока повреждение: пациент может
- 42. Центр произвольного движения глаз повреждение - лишает человека способности смещать глаза в направлении различных объектов. Центр
- 43. Сенсорные входы в моторную кору афференты из других областей коры соматосенсорной области теменной коры, передних отделов
- 44. Обратная связь мышечных сокращений с моторной корой сокращения мышц - сигналы обратно в моторную кору от
- 45. Нарушение функций моторной коры при инсульте поражение первичной моторной коры, соседних областей и более глубоких структур
- 46. Организация манипуляторных движений Манипуляторные движения – пример произвольных движений, которые обусловлены мотивацией. Эти движения локальны и
- 47. Иерархия форм двигательной активности по Н.А. Бернштейну Теория уровней построения движений: морфологические отделы нервной системы: спинной
- 48. Руброспинальный уровень эволюционно наиболее древний и созревающий раньше других, функционирует с первых недель жизни определяет мышечный
- 49. В. Таламопалидарный уровень обеспечивает переработку сигналов от мышечно-суставных рецепторов, которые сообщают о взаимном расположении частей тела.
- 50. С. Уровень пространственного поля или пирамидно-стриальный уровень поступает информация о состоянии внешней среды от экстерорецепторных анализаторов,
- 51. Д. Уровень предметных действий функционирует при обязательном участии коры (теменных и премоторных зон) и обеспечивает организацию
- 52. Е. Высший уровень организации движений интеллектуализированные двигательные акты: работа артикуляционного аппарата в звучащей речи, движения руки
- 53. Чувствительные и двигательные компоненты аппарата движения
- 54. Регуляция двигательной фунции МВ скелетных мышц α и γ-мотонейроны спинного мозга, двигательные ядра черепных нервов псевдоуниполярные
- 55. Мышечные волокна млекопитающих делятся на два основных типа; Экстрафузальные Интрафузальные Эмбриогенез: экстрафузальные приобретают полный комплект сократительных
- 56. Экстрафузальные МВ сократительная функция, участвуют в различных метаболических процессах, иннервируются альфа-мотонейронами, различают фазные и тонические фазные
- 57. ФАЗНЫЕ И ТОНИЧЕСКИЕ ВОЛОКНА Экстрафузальные мышечные волокна подразделяют на фазные, осуществляющие энергичные сокращения, произвольная мускулатура человека
- 58. Интрафузальные мышечные волокна и рецептор натяжения мышц- веретено Интрафузальные мышечные волокна специализированы для детектирования натяжения входят
- 59. Мышечные веретёна - чувствительный воспринимающий аппарат скелетной мышцы: их количество в мышцах варьирует структурные элементы мышечного
- 60. Орган Гольджи описан итальянским анатомо Камилло Гольджи (1843-1926) в 1800г. перерастяжение или выраженное напряжение мышцы активирует
- 61. Орган Гольджи (описан Камилло Гольджи (1843-1926) в 1800г.) расположены в концевой части сухожилия, а также в
- 62. Эффектный аппарат представлен несколькими типами мышечных волокон быстрые /белые промежуточные медленные/красные
- 63. Быстрые мышечные волокна (БМВ) клетки белого цвета (мало миоглобина и цитохромовых пигментов), мало митохондрий, СР хорошо
- 64. Медленные мышечные волокна, ММВ тонкие, к-во миофибрилл в них невелико, красного цвета (миоглобин и цитохромовые пигменты)
- 65. 1 МВ иннервируется 1-м мотонейроном (его аксон ветвится и иннервирует множество МВ - мультитерминальная иннервация) Группа
- 66. Моторная единица и моторный нейронный пул Моторная единица представлена альфа-мотонейроном и иннервируемыми им мышечными волокнами Моторный
- 67. Полинейронная иннервация окончательное формирование двигательных единиц происходит в постнатальном периоде до рождения каждое МВ иннервируется несколькими
- 68. Функциональная характеристика двигательных/моторных единиц (ДЕ=МЕ) МЕ содержат только один вид специфических мышечных волокон (тип 1 или
- 70. Типы волокон скелетной мышцы Гистохимическое выявление активности АТФазы миозина и сукцинатдегидрогеназы (СДГ) A (слева) — активность
- 71. ОКИСЛИТЕЛЬНЫЕ И ГЛИКОЛИТИЧЕСКИЕ МВ источник энергии МВ - гидролиз макроэрга АТФ митохондрии параллельно миофибриллам гликоген в
- 72. Креатин ген SC6A8 кодирует Na- и Cl-зависимый транспортёр креатина преимущественно ген экспрессируется в скелетной мышце и
- 73. БЫСТРЫЕ И МЕДЛЕННЫЕ МВ скоростные характеристики скелетной мышцы отражает активность АТФазы миозина скорость сокращения мышечного волокна
- 74. Окислительные, или красные мышечные волокна небольшого диаметра, окружены массой капилляров, содержат много миоглобина, многочисленные митохондрии имеют
- 76. Соотношение числа медленных и быстрых ДЕ в одной и той же мышце определено генетически и может
- 77. КОНТРОЛЬ ФЕНОТИПА МЫШЕЧНЫХ ВОЛОКОН иннервация, уровень физической активности, гуморальные факторы (в том числе анаболические стероиды) наследственно
- 78. ДЕНЕРВАЦИОННЫЙ СИНДРОМ повреждение мотонейронов или их аксонов - атрофия мышцы снижается количество миофибрилл и митохондрий, значительное
- 79. РЕИННЕРВАЦИЯ аксон растёт к собственному синапсу (>95%) новые терминали занимают старые постсинаптические складки прекращается синтез эмбриональной
- 80. Ветвление аксонов при реиннервации. А — интактная иннервация мышечных волокон. Б — при частичной денервации аксоны
- 81. ФИЗИОЛОГИЧЕСКАЯ РЕГЕНЕРАЦИЯ происходит постоянно обновление мышечных волокон циклы пролиферации клеток-сaтеллитов с последующей дифференцировкой в миобласты и
- 82. РЕПАРАТИВНАЯ РЕГЕНЕРАЦИЯ повторение событий эмбрионального миогенеза при гибели симпласта (после денервации) - фагоцитоз макрофагами активированные клетки-сателлиты
- 84. Скачать презентацию
Слайд 2Роль спинного мозга в процессах регуляции деятельности опорно-двигательного аппарата и вегетативных функций
Роль спинного мозга в процессах регуляции деятельности опорно-двигательного аппарата и вегетативных функций

Слайд 3Целевые функции двигательной системы
Целевые функции двигательной системы

Слайд 4Функции опорно-двигательного аппарата и системы его регуляции:
поддержание равновесия (позная/статическая деятельность мышц)
движения в
Функции опорно-двигательного аппарата и системы его регуляции:
поддержание равновесия (позная/статическая деятельность мышц)
движения в

Слайд 5Представление о пирамидной и экстрапирамидной системах
Представление о пирамидной и экстрапирамидной системах

Слайд 6Структуры, отвечающие за регуляцию позы и движений, находятся в разных отделах ЦНС
Структуры, отвечающие за регуляцию позы и движений, находятся в разных отделах ЦНС

Слайд 7Самый низший уровень в организации движения связан со спинным мозгом
чувствительные, вставочные и
Самый низший уровень в организации движения связан со спинным мозгом
чувствительные, вставочные и

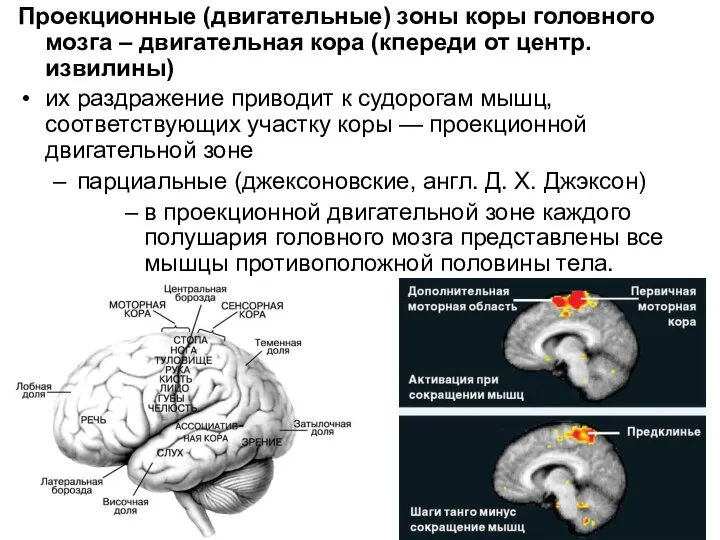
Слайд 8Проекционные (двигательные) зоны коры головного мозга – двигательная кора (кпереди от центр.извилины)
их
Проекционные (двигательные) зоны коры головного мозга – двигательная кора (кпереди от центр.извилины)
их
Слайд 9Двигательные пути, идущие от головного мозга к спинному, делятся на две системы
пирамидную
начало
Двигательные пути, идущие от головного мозга к спинному, делятся на две системы
пирамидную
начало

Слайд 10Экстрапирамидная система – совокупность структур головного мозга, участвующих в управлении движениями, поддержании
Экстрапирамидная система – совокупность структур головного мозга, участвующих в управлении движениями, поддержании

Слайд 11Экстрапирамидная система
эволюционно более древняя система моторного контроля
значение в построении и контроле движений,
Экстрапирамидная система
эволюционно более древняя система моторного контроля
значение в построении и контроле движений,

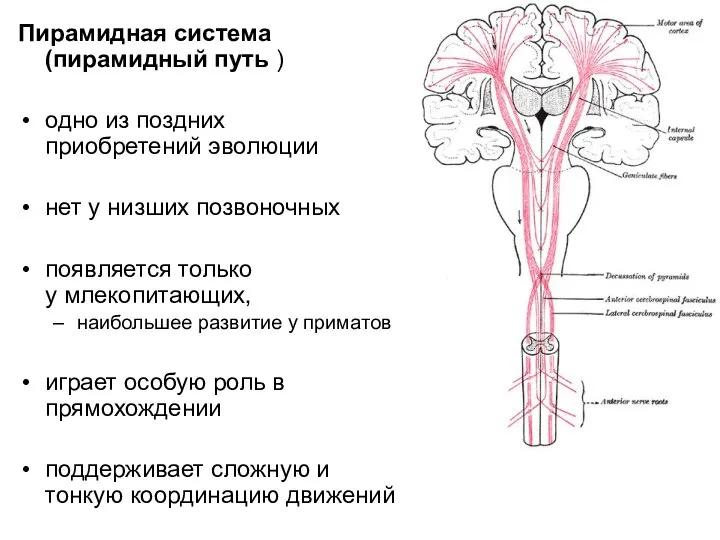
Слайд 12Пирамидная система (пирамидный путь )
одно из поздних приобретений эволюции
нет у низших позвоночных
появляется только
Пирамидная система (пирамидный путь )
одно из поздних приобретений эволюции
нет у низших позвоночных
появляется только
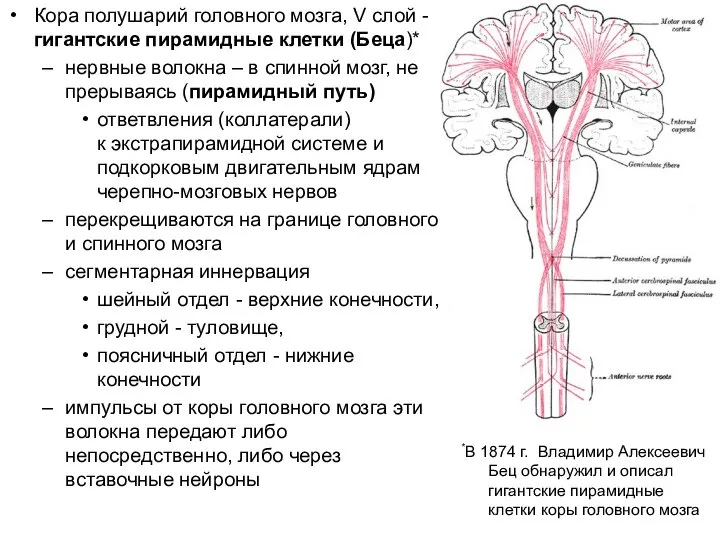
Слайд 13Кора полушарий головного мозга, V слой - гигантские пирамидные клетки (Беца)*
нервные волокна
Кора полушарий головного мозга, V слой - гигантские пирамидные клетки (Беца)*
нервные волокна
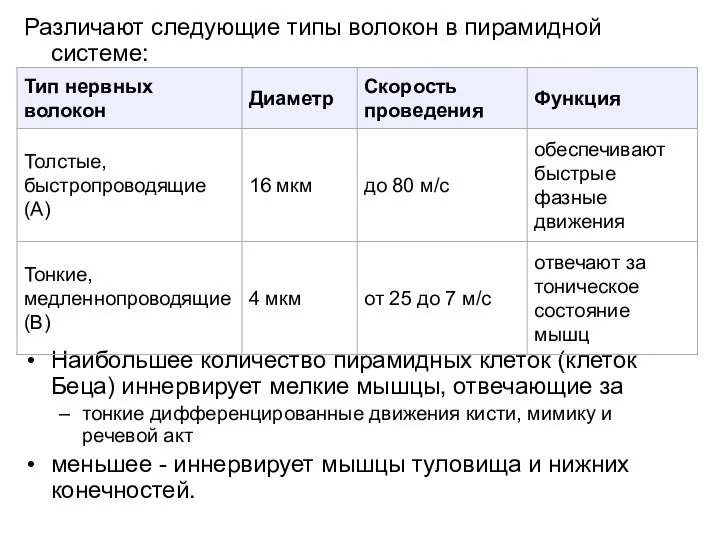
Слайд 14Различают следующие типы волокон в пирамидной системе:
Наибольшее количество пирамидных клеток (клеток Беца)
Различают следующие типы волокон в пирамидной системе:
Наибольшее количество пирамидных клеток (клеток Беца)
Слайд 15Повреждения пирамидной системы проявляются
параличами,
парезами,
патологическими рефлексами.
Поражение пирамидной системы может быть
Повреждения пирамидной системы проявляются
параличами,
парезами,
патологическими рефлексами.
Поражение пирамидной системы может быть

Слайд 16Общие принципы регуляции двигательной функции
Общие принципы регуляции двигательной функции

Слайд 17Произвольные динамические движения мышц – целенаправленное действие:
побуждение к действию (драйв, мотивация)→замысел
Произвольные динамические движения мышц – целенаправленное действие:
побуждение к действию (драйв, мотивация)→замысел

Слайд 18Двигательная система - нейронные механизмы, участвующие в реализации фазной и позной мышечной
Двигательная система - нейронные механизмы, участвующие в реализации фазной и позной мышечной

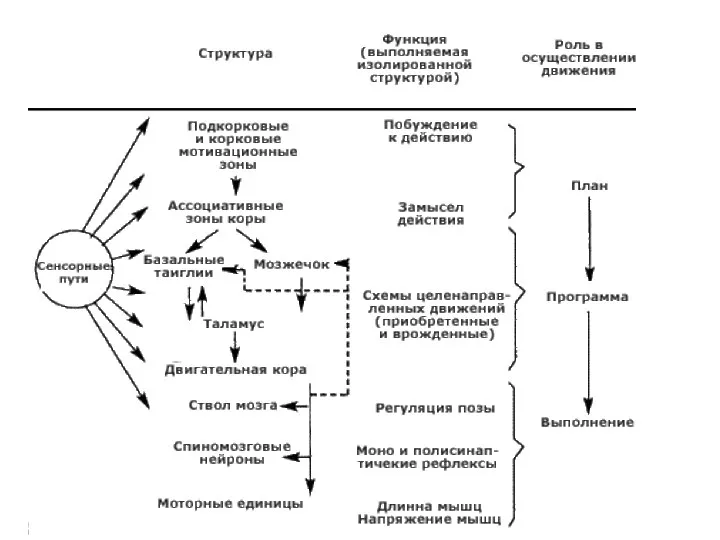
Слайд 20Общий план организации двигательной системы (ДС): 3 этажа (по Н.А. Агаджаняну, 2006)
Спинной
Общий план организации двигательной системы (ДС): 3 этажа (по Н.А. Агаджаняну, 2006)
Спинной

Слайд 21Стволовые структуры мозга (вестибулярные ядра, красное ядро, ретикулярная формация, покрышка четверохолмия) –
Стволовые структуры мозга (вестибулярные ядра, красное ядро, ретикулярная формация, покрышка четверохолмия) –

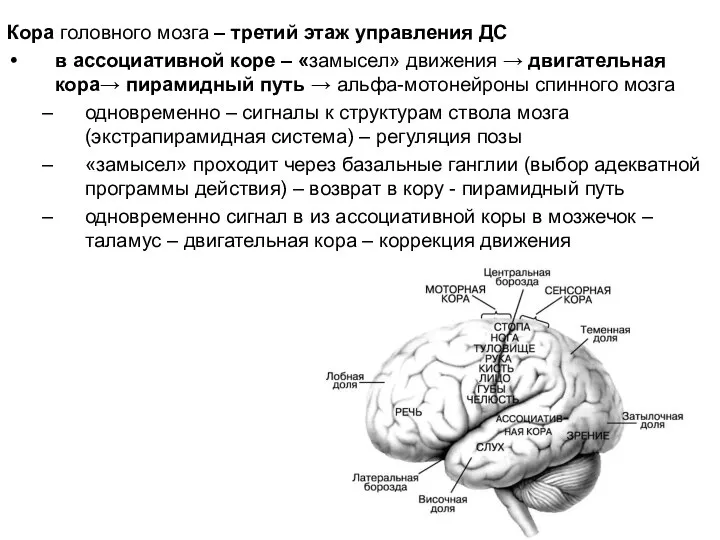
Слайд 22Кора головного мозга – третий этаж управления ДС
в ассоциативной коре – «замысел»
Кора головного мозга – третий этаж управления ДС
в ассоциативной коре – «замысел»
Слайд 23Все звенья ДС используют сенсорную информацию от
рецепторов мышц
мышечные веретена,
сухожильные рецепторы Гольджи
рецепторы
Все звенья ДС используют сенсорную информацию от
рецепторов мышц
мышечные веретена,
сухожильные рецепторы Гольджи
рецепторы

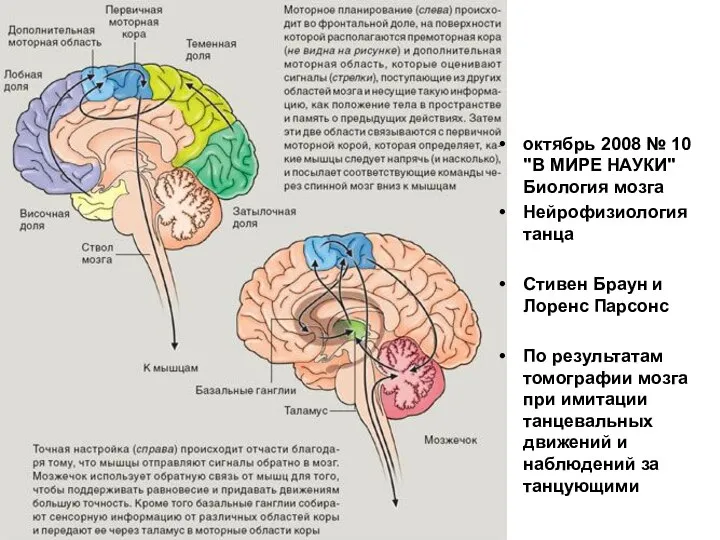
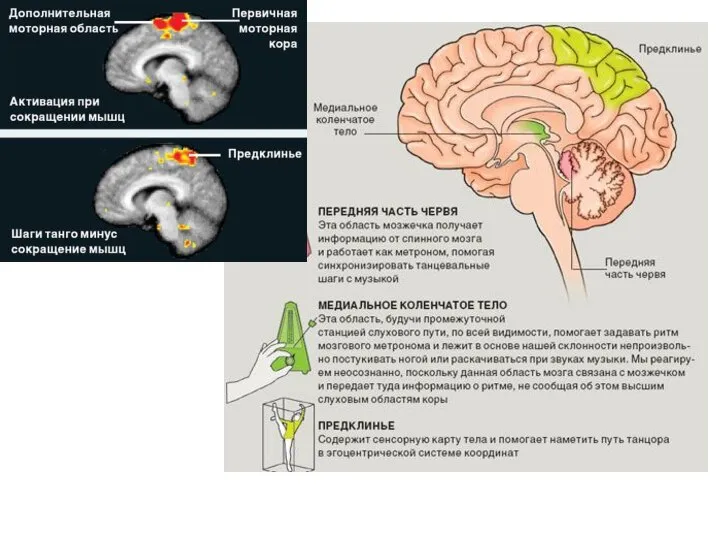
Слайд 24октябрь 2008 № 10 "В МИРЕ НАУКИ"
Биология мозга
Нейрофизиология танца
Стивен Браун и Лоренс
октябрь 2008 № 10 "В МИРЕ НАУКИ"
Биология мозга
Нейрофизиология танца
Стивен Браун и Лоренс
Слайд 26Контроль позы и движений
Контроль позы и движений

Слайд 27Схема контроля произвольных движений
Схема контроля произвольных движений
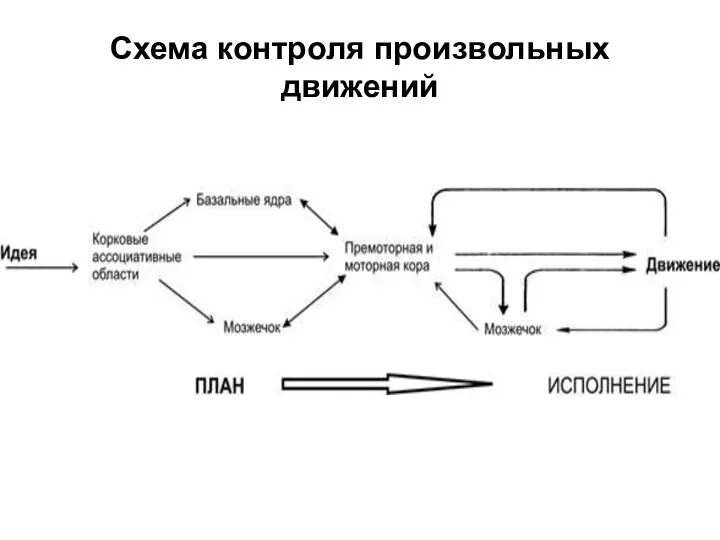
Слайд 28Общий конечный путь к скелетным мышцам от
мотонейронов спинного мозга и
нейронов двигательных
Общий конечный путь к скелетным мышцам от
мотонейронов спинного мозга и
нейронов двигательных

Слайд 29Мотонейроны спинного мозга и нейроны двигательных ядер черепных нервов — общий конечный путь
Мотонейроны спинного мозга и нейроны двигательных ядер черепных нервов — общий конечный путь
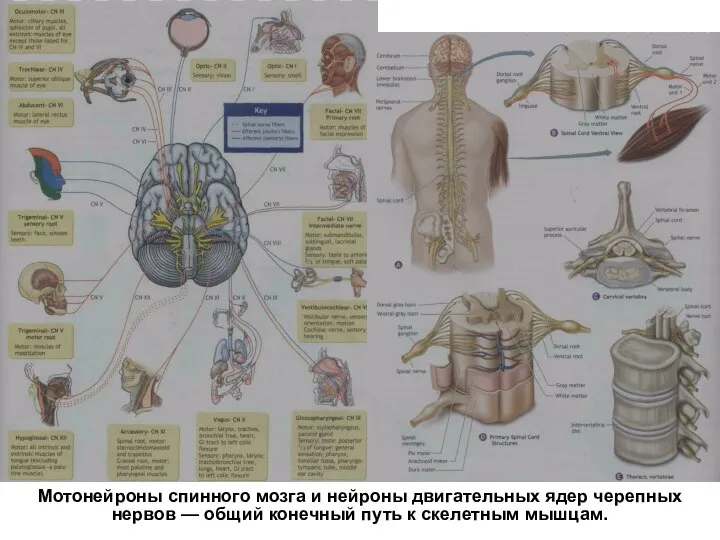
Слайд 30Роль спинного мозга в регуляций двигательной активности
Примитивные процессы регуляции скелетных мышц
фазные движения
Роль спинного мозга в регуляций двигательной активности
Примитивные процессы регуляции скелетных мышц
фазные движения

Слайд 31Роль ствола мозга в регуляции движений
Продолговатый мозг, мост, средний мозг
двигательные ядра черепно-мозговых
Роль ствола мозга в регуляции движений
Продолговатый мозг, мост, средний мозг
двигательные ядра черепно-мозговых
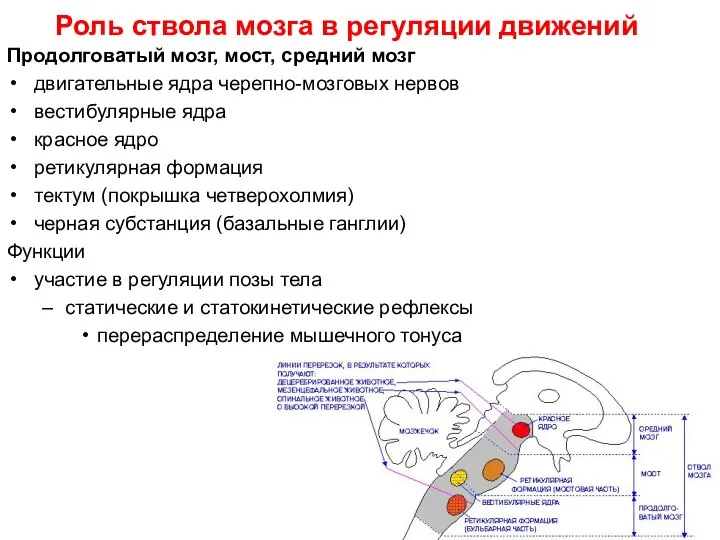
Слайд 32«Базальные ядра»
хвостатое ядро,
скорлупа,
субталамическое
(подбугорное) ядро,
бледный шар и чёрная субстанция
Координация
«Базальные ядра»
хвостатое ядро,
скорлупа,
субталамическое
(подбугорное) ядро,
бледный шар и чёрная субстанция
Координация
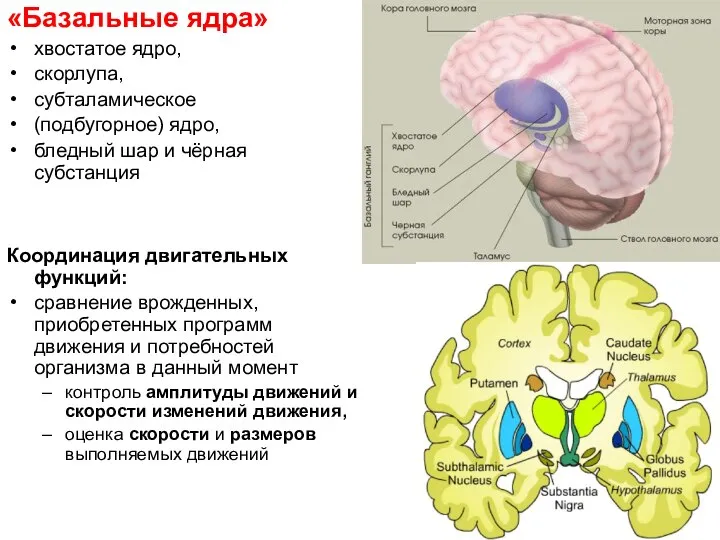
Слайд 33(Fredrik Ullén, доцент Королевского Каролинского института и Стокгольмского института мозга, 2010).
сложные движения
(Fredrik Ullén, доцент Королевского Каролинского института и Стокгольмского института мозга, 2010).
сложные движения

Слайд 34Патология базальных ядер
Гипер/гипокинетические состояния
Хорея (от греч. choreia — хоровод, пляска)
Атетоз (от греч.
Патология базальных ядер
Гипер/гипокинетические состояния
Хорея (от греч. choreia — хоровод, пляска)
Атетоз (от греч.

Слайд 35Мозжечок в системе двигательного контроля
Три уровня участия в системе контроля и координации
Мозжечок в системе двигательного контроля
Три уровня участия в системе контроля и координации

Слайд 37Мозжечковые симптомы определяются местом повреждения
Дисметрия — неспособность правильной оценки расстояния → неспособность сразу
Мозжечковые симптомы определяются местом повреждения
Дисметрия — неспособность правильной оценки расстояния → неспособность сразу

Слайд 38Роль коры больших полушарий в двигательной функции
Ассоциативная кора
замысел будущего действия
Двигательная кора
Роль коры больших полушарий в двигательной функции
Ассоциативная кора
замысел будущего действия
Двигательная кора
Слайд 39Первичная моторная кора
картированы участки, ответственные за сокращения отдельных мышц, но чаще возбуждаются
Первичная моторная кора
картированы участки, ответственные за сокращения отдельных мышц, но чаще возбуждаются

Слайд 40Дополнительная моторная область
функционирует совместно с премоторной областью,
обеспечивает движения,
поддерживающие осанку,
фиксацию движений
Дополнительная моторная область
функционирует совместно с премоторной областью,
обеспечивает движения,
поддерживающие осанку,
фиксацию движений
Слайд 41Центр формирования речи
перед первичной моторной корой выше сильвиевой борозды - область Брока
повреждение:
пациент
Центр формирования речи
перед первичной моторной корой выше сильвиевой борозды - область Брока
повреждение:
пациент
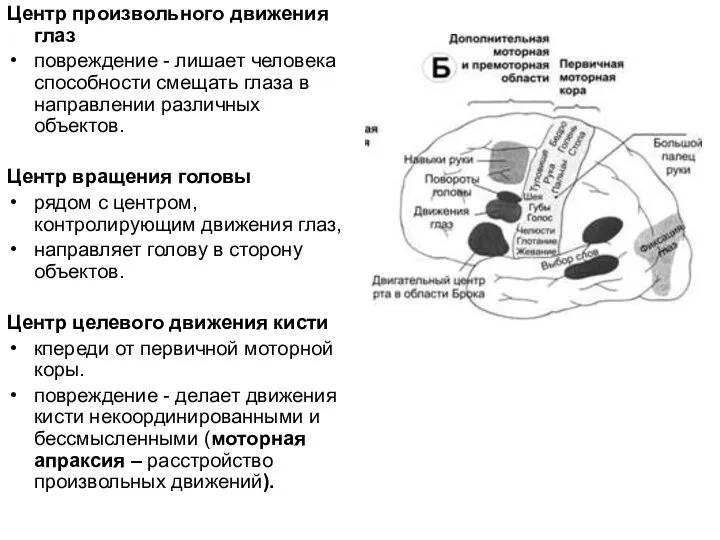
Слайд 42Центр произвольного движения глаз
повреждение - лишает человека способности смещать глаза в направлении
Центр произвольного движения глаз
повреждение - лишает человека способности смещать глаза в направлении
Слайд 43Сенсорные входы в моторную кору
афференты из других областей коры
соматосенсорной области теменной коры,
Сенсорные входы в моторную кору
афференты из других областей коры
соматосенсорной области теменной коры,

Слайд 44Обратная связь мышечных сокращений с моторной корой
сокращения мышц - сигналы обратно в
Обратная связь мышечных сокращений с моторной корой
сокращения мышц - сигналы обратно в

Слайд 45Нарушение функций моторной коры
при инсульте поражение первичной моторной коры, соседних областей и
Нарушение функций моторной коры
при инсульте поражение первичной моторной коры, соседних областей и

Слайд 46Организация манипуляторных движений
Манипуляторные движения – пример произвольных движений, которые обусловлены мотивацией.
Эти
Организация манипуляторных движений
Манипуляторные движения – пример произвольных движений, которые обусловлены мотивацией.
Эти

Слайд 47Иерархия форм двигательной активности по Н.А. Бернштейну
Теория уровней построения движений:
морфологические отделы нервной системы:
Иерархия форм двигательной активности по Н.А. Бернштейну
Теория уровней построения движений:
морфологические отделы нервной системы:

Слайд 48Руброспинальный уровень
эволюционно наиболее древний и созревающий раньше других,
функционирует с первых недель
Руброспинальный уровень
эволюционно наиболее древний и созревающий раньше других,
функционирует с первых недель

Слайд 49В. Таламопалидарный уровень
обеспечивает переработку сигналов от мышечно-суставных рецепторов, которые сообщают о взаимном
В. Таламопалидарный уровень
обеспечивает переработку сигналов от мышечно-суставных рецепторов, которые сообщают о взаимном

Слайд 50С. Уровень пространственного поля или пирамидно-стриальный уровень
поступает информация о состоянии внешней среды
С. Уровень пространственного поля или пирамидно-стриальный уровень
поступает информация о состоянии внешней среды

Слайд 51Д. Уровень предметных действий
функционирует при обязательном участии коры (теменных и премоторных зон)
Д. Уровень предметных действий
функционирует при обязательном участии коры (теменных и премоторных зон)

Слайд 52Е. Высший уровень организации движений
интеллектуализированные двигательные акты:
работа артикуляционного аппарата в звучащей речи,
движения
Е. Высший уровень организации движений
интеллектуализированные двигательные акты:
работа артикуляционного аппарата в звучащей речи,
движения

Слайд 53Чувствительные и двигательные компоненты аппарата движения
Чувствительные и двигательные компоненты аппарата движения

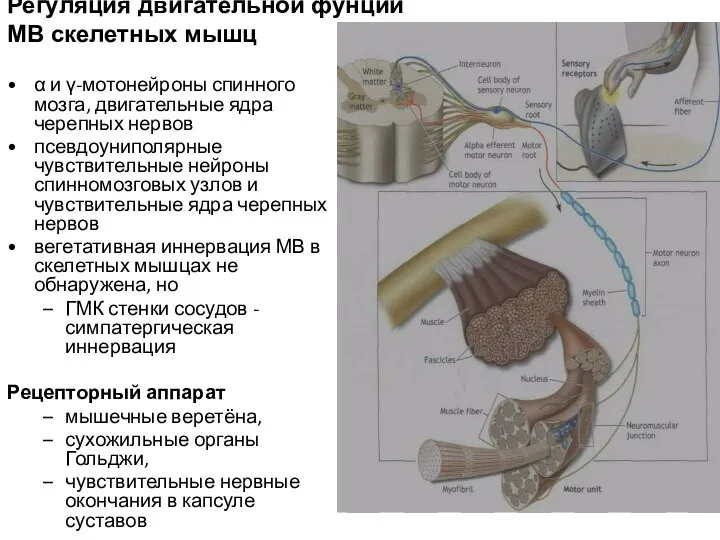
Слайд 54Регуляция двигательной фунции МВ скелетных мышц
α и γ-мотонейроны спинного мозга, двигательные
Регуляция двигательной фунции МВ скелетных мышц
α и γ-мотонейроны спинного мозга, двигательные
Слайд 55Мышечные волокна млекопитающих делятся на два основных типа;
Экстрафузальные
Интрафузальные
Эмбриогенез:
экстрафузальные приобретают полный комплект
Мышечные волокна млекопитающих делятся на два основных типа;
Экстрафузальные
Интрафузальные
Эмбриогенез:
экстрафузальные приобретают полный комплект

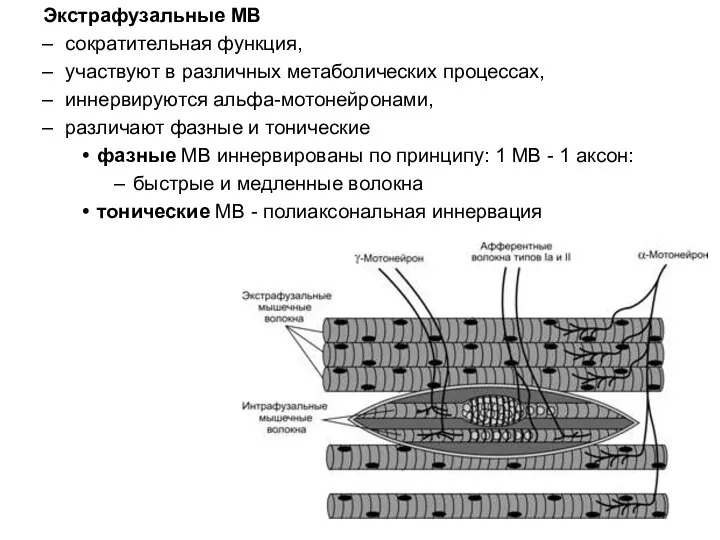
Слайд 56Экстрафузальные МВ
сократительная функция,
участвуют в различных метаболических процессах,
иннервируются альфа-мотонейронами,
различают фазные и тонические
фазные МВ
Экстрафузальные МВ
сократительная функция,
участвуют в различных метаболических процессах,
иннервируются альфа-мотонейронами,
различают фазные и тонические
фазные МВ
Слайд 57ФАЗНЫЕ И ТОНИЧЕСКИЕ ВОЛОКНА
Экстрафузальные мышечные волокна подразделяют на
фазные, осуществляющие энергичные сокращения,
ФАЗНЫЕ И ТОНИЧЕСКИЕ ВОЛОКНА
Экстрафузальные мышечные волокна подразделяют на
фазные, осуществляющие энергичные сокращения,

Слайд 58Интрафузальные мышечные волокна и рецептор натяжения мышц- веретено
Интрафузальные мышечные волокна
специализированы для
Интрафузальные мышечные волокна и рецептор натяжения мышц- веретено
Интрафузальные мышечные волокна
специализированы для

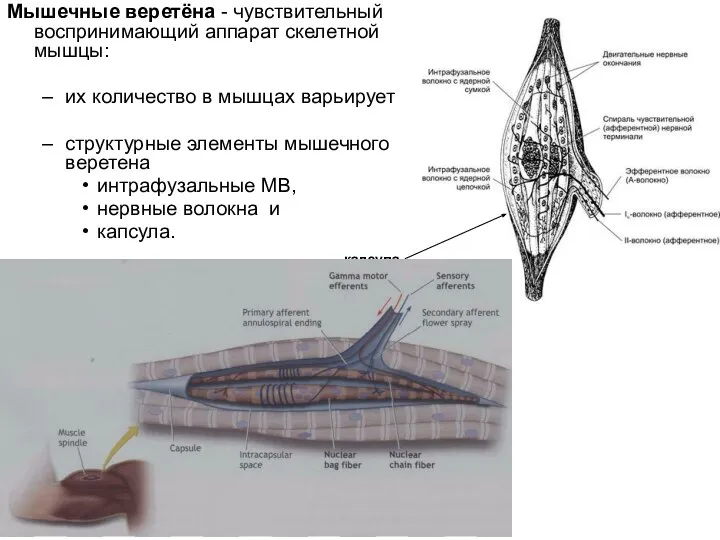
Слайд 59Мышечные веретёна - чувствительный воспринимающий аппарат скелетной мышцы:
их количество в мышцах варьирует
структурные
Мышечные веретёна - чувствительный воспринимающий аппарат скелетной мышцы:
их количество в мышцах варьирует
структурные
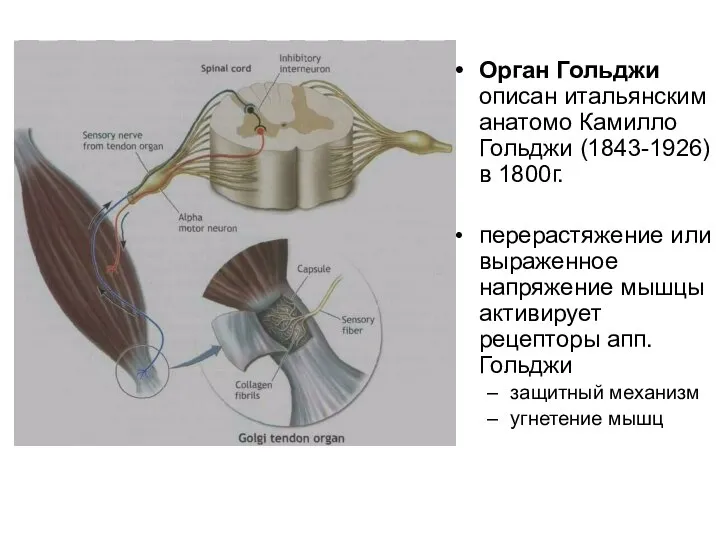
Слайд 60Орган Гольджи описан итальянским анатомо Камилло Гольджи (1843-1926) в 1800г.
перерастяжение или выраженное
Орган Гольджи описан итальянским анатомо Камилло Гольджи (1843-1926) в 1800г.
перерастяжение или выраженное
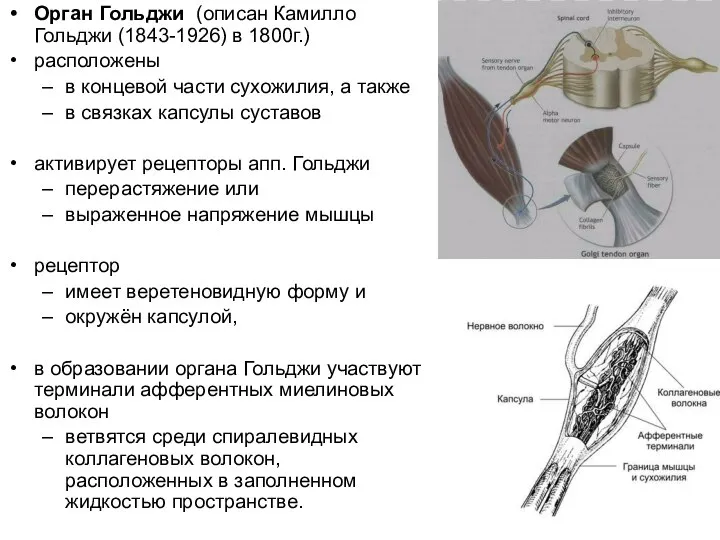
Слайд 61Орган Гольджи (описан Камилло Гольджи (1843-1926) в 1800г.)
расположены
в концевой части сухожилия,
Орган Гольджи (описан Камилло Гольджи (1843-1926) в 1800г.)
расположены
в концевой части сухожилия,
Слайд 62Эффектный аппарат представлен несколькими типами мышечных волокон
быстрые /белые
промежуточные
медленные/красные
Эффектный аппарат представлен несколькими типами мышечных волокон
быстрые /белые
промежуточные
медленные/красные

Слайд 63Быстрые мышечные волокна (БМВ)
клетки белого цвета (мало миоглобина и цитохромовых пигментов),
мало митохондрий,
Быстрые мышечные волокна (БМВ)
клетки белого цвета (мало миоглобина и цитохромовых пигментов),
мало митохондрий,

Слайд 64Медленные мышечные волокна, ММВ
тонкие, к-во миофибрилл в них невелико,
красного цвета (миоглобин и
Медленные мышечные волокна, ММВ
тонкие, к-во миофибрилл в них невелико,
красного цвета (миоглобин и

Слайд 651 МВ иннервируется 1-м мотонейроном (его аксон ветвится и иннервирует множество МВ
1 МВ иннервируется 1-м мотонейроном (его аксон ветвится и иннервирует множество МВ
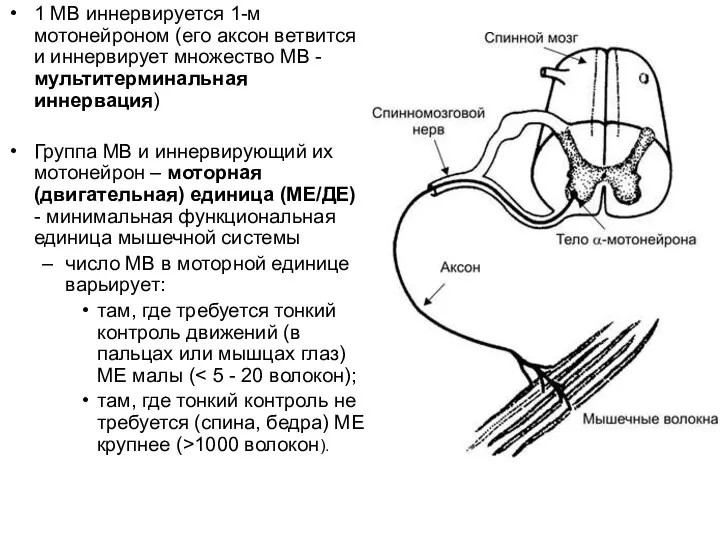
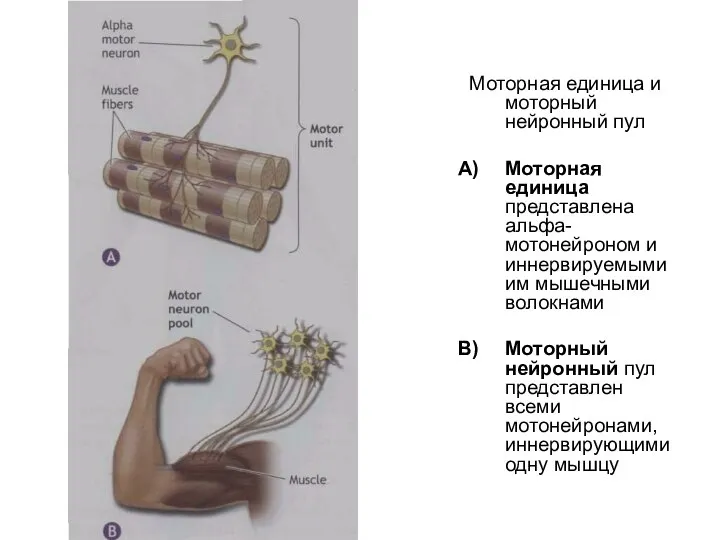
Слайд 66Моторная единица и моторный нейронный пул
Моторная единица представлена альфа-мотонейроном и иннервируемыми им
Моторная единица и моторный нейронный пул
Моторная единица представлена альфа-мотонейроном и иннервируемыми им
Слайд 67Полинейронная иннервация
окончательное формирование двигательных единиц происходит в постнатальном периоде
до рождения каждое МВ
Полинейронная иннервация
окончательное формирование двигательных единиц происходит в постнатальном периоде
до рождения каждое МВ

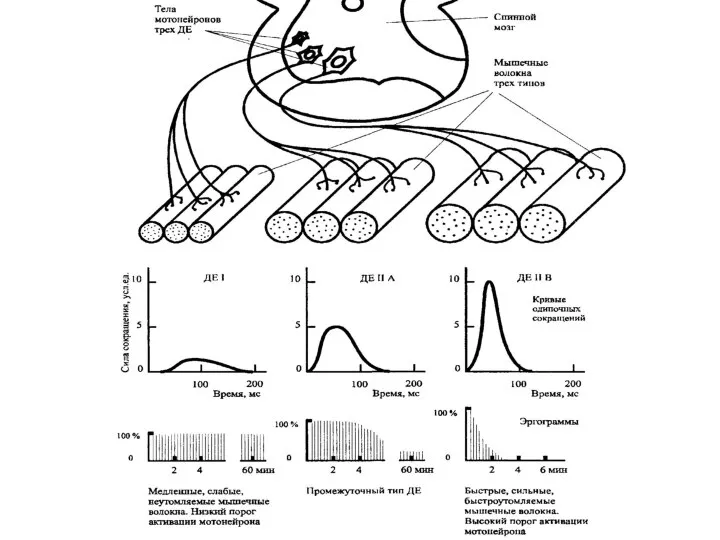
Слайд 68Функциональная характеристика двигательных/моторных единиц (ДЕ=МЕ)
МЕ содержат только один вид специфических мышечных волокон
Функциональная характеристика двигательных/моторных единиц (ДЕ=МЕ)
МЕ содержат только один вид специфических мышечных волокон

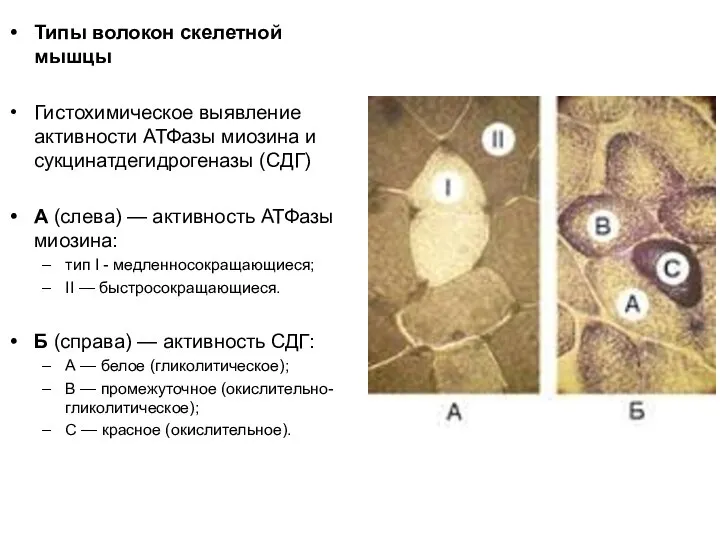
Слайд 70Типы волокон скелетной мышцы
Гистохимическое выявление активности АТФазы миозина и сукцинатдегидрогеназы (СДГ)
A (слева) — активность
Типы волокон скелетной мышцы
Гистохимическое выявление активности АТФазы миозина и сукцинатдегидрогеназы (СДГ)
A (слева) — активность
Слайд 71ОКИСЛИТЕЛЬНЫЕ И ГЛИКОЛИТИЧЕСКИЕ МВ
источник энергии МВ - гидролиз макроэрга АТФ
митохондрии параллельно миофибриллам
гликоген в
ОКИСЛИТЕЛЬНЫЕ И ГЛИКОЛИТИЧЕСКИЕ МВ
источник энергии МВ - гидролиз макроэрга АТФ
митохондрии параллельно миофибриллам
гликоген в

Слайд 72Креатин
ген SC6A8 кодирует Na- и Cl-зависимый транспортёр креатина
преимущественно ген экспрессируется в скелетной мышце и почке, в
Креатин
ген SC6A8 кодирует Na- и Cl-зависимый транспортёр креатина
преимущественно ген экспрессируется в скелетной мышце и почке, в

Слайд 73БЫСТРЫЕ И МЕДЛЕННЫЕ МВ
скоростные характеристики скелетной мышцы отражает
активность АТФазы миозина
скорость сокращения мышечного
БЫСТРЫЕ И МЕДЛЕННЫЕ МВ
скоростные характеристики скелетной мышцы отражает
активность АТФазы миозина
скорость сокращения мышечного

Слайд 74Окислительные, или красные мышечные волокна
небольшого диаметра,
окружены массой капилляров,
содержат много миоглобина,
многочисленные митохондрии
Окислительные, или красные мышечные волокна
небольшого диаметра,
окружены массой капилляров,
содержат много миоглобина,
многочисленные митохондрии

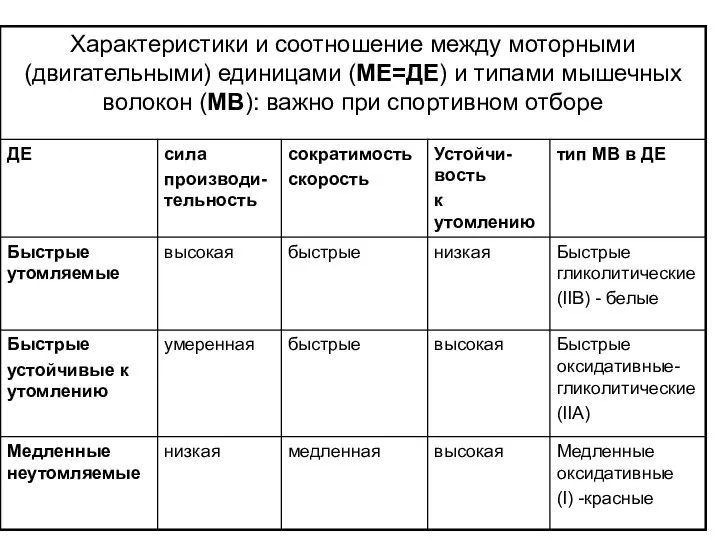
Слайд 76Соотношение числа медленных и быстрых ДЕ в одной и той же мышце
Соотношение числа медленных и быстрых ДЕ в одной и той же мышце

Слайд 77КОНТРОЛЬ ФЕНОТИПА МЫШЕЧНЫХ ВОЛОКОН
иннервация, уровень физической активности, гуморальные факторы (в том числе
КОНТРОЛЬ ФЕНОТИПА МЫШЕЧНЫХ ВОЛОКОН
иннервация, уровень физической активности, гуморальные факторы (в том числе

Слайд 78ДЕНЕРВАЦИОННЫЙ СИНДРОМ
повреждение мотонейронов или их аксонов - атрофия мышцы
снижается количество миофибрилл и
ДЕНЕРВАЦИОННЫЙ СИНДРОМ
повреждение мотонейронов или их аксонов - атрофия мышцы
снижается количество миофибрилл и

Слайд 79РЕИННЕРВАЦИЯ
аксон растёт к собственному синапсу (>95%)
новые терминали занимают старые постсинаптические складки
прекращается синтез
РЕИННЕРВАЦИЯ
аксон растёт к собственному синапсу (>95%)
новые терминали занимают старые постсинаптические складки
прекращается синтез

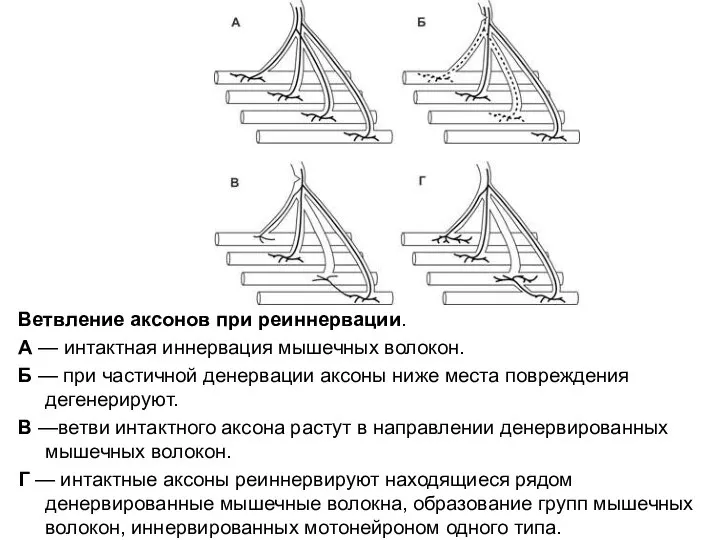
Слайд 80Ветвление аксонов при реиннервации.
А — интактная иннервация мышечных волокон.
Б — при частичной денервации аксоны
Ветвление аксонов при реиннервации.
А — интактная иннервация мышечных волокон.
Б — при частичной денервации аксоны
Слайд 81ФИЗИОЛОГИЧЕСКАЯ РЕГЕНЕРАЦИЯ
происходит постоянно
обновление мышечных волокон
циклы пролиферации клеток-сaтеллитов с последующей дифференцировкой в
ФИЗИОЛОГИЧЕСКАЯ РЕГЕНЕРАЦИЯ
происходит постоянно
обновление мышечных волокон
циклы пролиферации клеток-сaтеллитов с последующей дифференцировкой в

Слайд 82РЕПАРАТИВНАЯ РЕГЕНЕРАЦИЯ
повторение событий эмбрионального миогенеза
при гибели симпласта (после денервации) - фагоцитоз макрофагами
активированные
РЕПАРАТИВНАЯ РЕГЕНЕРАЦИЯ
повторение событий эмбрионального миогенеза
при гибели симпласта (после денервации) - фагоцитоз макрофагами
активированные

 Цветок и плод
Цветок и плод Животные Саратовской области
Животные Саратовской области Внутреннее строение листа. 6 класс
Внутреннее строение листа. 6 класс Пауки и другие паукообразные
Пауки и другие паукообразные Разнообразие внешнего строения бактериальных клеток
Разнообразие внешнего строения бактериальных клеток Взаимодействие генов. 11 класс
Взаимодействие генов. 11 класс Микроструктура нервной ткани
Микроструктура нервной ткани Царство грибов
Царство грибов Секреты бережливости. Электронная выставка
Секреты бережливости. Электронная выставка Черепные нервы
Черепные нервы Жизнь в пресных водах
Жизнь в пресных водах Координация и регуляция
Координация и регуляция Мышечная и нервная ткани
Мышечная и нервная ткани Анатомия осевого скелета
Анатомия осевого скелета Класс Паукообразные
Класс Паукообразные Покровная системы организма
Покровная системы организма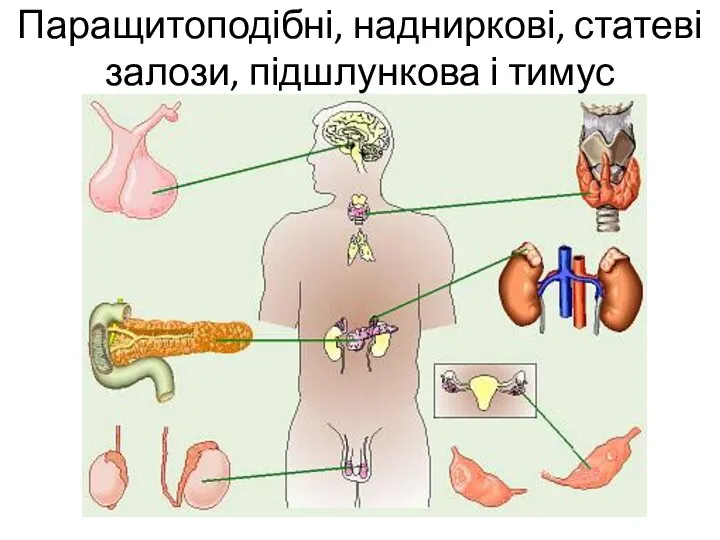 Паращитоподібні, надниркові, статеві залози, підшлункова і тимус
Паращитоподібні, надниркові, статеві залози, підшлункова і тимус Особенности поведения земноводных и пресмыкающихся осенью
Особенности поведения земноводных и пресмыкающихся осенью Мышцы груди человека
Мышцы груди человека Перелётные и зимующие птицы
Перелётные и зимующие птицы Систематическое положение человека (8)
Систематическое положение человека (8) Эти забавные домашние питомцы
Эти забавные домашние питомцы Fauna
Fauna Игра Редкие бабочки
Игра Редкие бабочки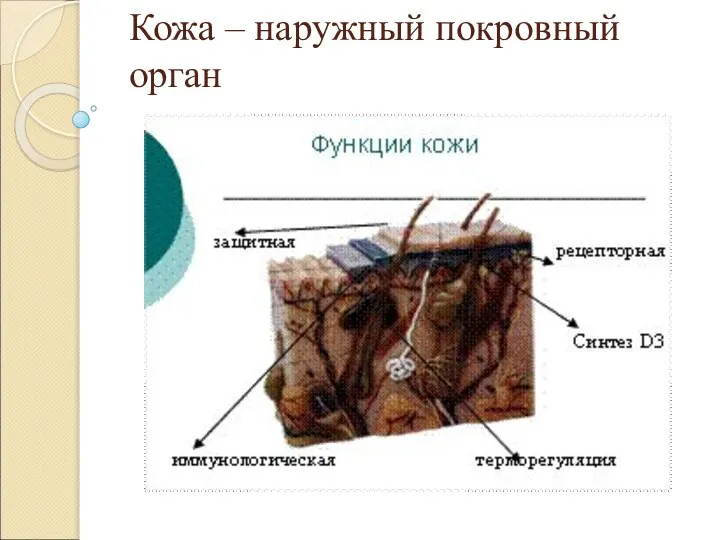 Кожа – наружный покровный орган
Кожа – наружный покровный орган Деление клетки. Митоз
Деление клетки. Митоз Презентация на тему КЛАССИФИКАЦИЯ МОЛЛЮСКОВ
Презентация на тему КЛАССИФИКАЦИЯ МОЛЛЮСКОВ  Тли
Тли