- Общая вирусология

Содержание
- 2. Основные отличия вирусов от других форм жизни один тип нуклеиновой кислоты отсутствие клеточного строения белоксинтезирующих систем
- 3. Формы существования вирусов внеклеточная = вирион (структура) : НК капсид [суперкапсид] . Н-р, вирион имеет форму…
- 4. Основные признаки, используемые для классификации вирусов тип нуклеиновой кислоты (ДНК/РНК), структура генома – количество нитей (цепочек)
- 5. Иерархическая система таксонов, применяемых в вирусологии Царство: Vira Подцарства: ДНК-геномные вирусы, РНК-геномные вирусы. Семейство - название
- 6. КЛАССИФИКАЦИЯ ВИРУСОВ
- 7. КЛАССИФИКАЦИЯ ВИРУСОВ
- 8. Принцип строения вириона Простой: НК+ капсид = нуклеокапсид Сложный: нуклеокапсид + суперкапсид
- 9. Типы симметрии капсида спиральная кубическая
- 10. Принцип строения суперкапсида билипидный слой матричный белок гликопротеины (шипы, ворсинки)
- 11. Форма вирионов
- 12. Общая характеристика ДНК вирионов форма: Линейная, Кольцевая. на концах – идентичные повторы: маркеры вирусной ДНК (не
- 13. Общая характеристика РНК вирусов форма: Линейная, Кольцевая. структура: Цельная, Фрагментированная. информационная функция: +нить (позитивный геном) =
- 14. Общая характеристика белков вирусов Структурные Капсидные – образуют капсид, «Внутренние», гистоноподобные – связаны с нуклеиновой кислотой
- 15. Схема строения простоустроенного вириона = паповавируса (вирус имеет двунитевую кольцевую ДНК)
- 16. Схема строения вируса гепатита А (вирус имеет однонитевую +РНК)
- 17. Схема строения сложноустроенного вириона= вируса герпеса (вирус с линейной двухнитевой ДНК)
- 18. Схема строения вируса гриппа = вирус с однонитевой фрагментированной (8 фрагментов) минус РНК
- 19. Свойство вирусов = строгий цитотропизм = Избирательность поражения вирусами определенных клеток, = способность вирусов к репликации
- 20. Патологические процессы, вызываемые вирусами инфекционные (микробные) болезни = вирусные инфекции, Опухоли.
- 21. Исходы вирусной инфекции клетки
- 22. Репродукция вирусов Различают три типа взаимодействия вируса с клеткой: 1. продуктивный тип, при котором образуются новые
- 23. Продуктивный тип взаимодействия вируса с клеткой Этапы размножения вирусов в чувствительной клетке: адсорбция вирионов на клетке
- 24. 1. Адсорбция вирионов на клетке = прикрепление вириона к поверхности клетки: 2 фазы: - неспецифическая –
- 25. 2. Проникновение вируса в клетку 3 пути: Рецептор-зависимый эндоцитоз, Слияние оболочки вириона с клеточной мембраной, Смешанный.
- 26. 2.1.Проникновение вируса в клетку: Рецептор-зависимый эндоцитоз =захватывание и поглощение вириона клеткой: 1. Клеточная мембрана с вирионом
- 27. 2.2.Проникновение вируса в клетку - слияние оболочки вириона с клеточной мембраной = виропексис: – характерно для
- 28. 2а.Депротеинизация вирусов= «раздевание» = освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых) оболочки (-чек) При виропексисе
- 29. 3. Синтез вирусных компонентов = дизъюнктивная репродукция = синтез вирусных белков и нуклеиновых кислот, = происходит
- 30. 1. Синтез вирусных белков В зараженной клетке вирусный геном кодирует синтез 2-х групп белков: Структурные =
- 31. 3.1.а.Синтез вирусных белков –варианты: ДНК-содержащие вирусы: Геномная ДНК вируса ↓ транскрипция иРНК ↓ трансляция белка вируса.
- 32. 3.1.б. Синтез вирусных белков -варианты: Плюс-нитевые РНК-содержащие вирусы = вирусный геном выполняет функцию иРНК (пикорнавирусы, флавивирусы,
- 33. 3.1.в. Синтез вирусных белков -варианты: Минус-нитевые РНК-содержащие вирусы (ортомиксовирусы, парамиксовирусы, рабдовирусы) и двунитевые (реовирусы): Геномная РНК
- 34. 3.1.г.Синтез вирусных белков -варианты: Ретровирусы: – геном состоит из 2-х одинаковых молекул РНК = диплоидный, имеют
- 35. 3.2. Репликация вирусных геномов – зависит от типа нуклеиновой кислоты, - наличия вирусоспецифических или клеточных полимераз,
- 36. 3.2.а. Репликация вирусных геномов - варианты: Двунитевые ДНК-вирусы (аденовирусы, герпесвирусы, поксвирусы) = полуконсервативный механизм: - происходит
- 37. 3.2.б.Репликация вирусных геномов Однонитевые ДНК-вирусы (парвовирусы) – используют клеточные ДНК-полимеразы: - на исходной вирусной ДНК (+нить)
- 38. 3.2.в.Репликация вирусных геномов Плюс-однонитевые РНК-вирусы (пикорнавирусы, флавивирусы, тогавирусы, полиовирусы) = геномная нить РНК выполняет функцию иРНК:
- 39. 3.2.г.Репликация вирусных геномов Минус-однонитевые РНК-вирусы (Рабдовирусы, парамиксовирусы, ортомиксовирусы) – имеют РНК-зависимую РНК-полимеразу: Минус-нитевая РНК + РНК-полимераза
- 40. 3.2.д.Репликация вирусных геномов Двунитевые РНК-вирусы (реовирусы, ротавирусы) – как у минус нитевых, но в цитоплазме клеток.
- 41. 3.2.е.Репликация вирусных геномов Ретровирусы = плюс-нитевые диплоидные РНК-содержащие вирусы, имеют обратную транскриптазу: обратная транскриптаза на матрице
- 42. 4.Формирование вирусов Происходит путем самосборки = составные части вируса транспортируются в определенный участок цитоплазмы или ядра
- 43. 5.Выход вирусов из клетки 1. взрывной путь: клетка погибает и вирусы выходят наружу = простоустроенные вирусы,
- 44. Абортивный тип взаимодействия вируса с клеткой = прерывание инфекционного процесса в клетке на одном из этапов,
- 45. Интегративный тип взаимодействия вируса с клеткой = вирогения = нуклеиновая кислота вируса встраивается в хромосому клетки
- 46. Значение вирогении 1. Сохранение вирусной информации в составе клеточного генома = персистенция: → клетка при этом
- 47. Исходы активации персистирующего вируса рецидив того же заболевания, развитие другого заболевания, вызываемого тем же самым вирусом,
- 48. Способы культивирования вирусов 3 модели: куриный эмбрион культура клеток организм лабораторного животного ⇓ обнаружение наличия вируса
- 49. Использование для вирусологического метода куриного эмбриона
- 50. Использование для вирусологического метода куриного эмбриона 5-7-дневные, реже – 10-11-дневные Основные способы заражения куриных эмбрионов на
- 52. Обнаружение вирусов в курином эмбрионе индикация: гибель эмбриона, морфологические изменения эмбриона/оболочек, РГА с жидкостью из полостей
- 53. Использование культур клеток Культуры клеток = соматические или эмбриональные клетки человека или животных, культивируемые в лабораторных
- 55. Первичные (эмбриональные) культуры клеток получают из тканей (эмбриональных или нормальных) многоклеточных организмов. Такие клетки не способны
- 57. Перевиваемые культуры клеток Перевиваемые = стабильные = готовят из опухолевых клеток, способных длительно расти и размножаться
- 59. Преимущества перевиваемых культур клеток перед первичными: продолжительность культивирования – десятки лет, высокая скорость размножения, меньшая трудоемкость,
- 60. Полуперевиваемые культуры клеток = диплоидные клетки различных тканей и органов, способные к ограниченному размножению in vitro.
- 62. Условия культивирования клеток: Использование лабораторной посуды из нейтрального стекла – пробирки, флаконы, матрасы (=флакон 4-х гранной
- 68. Обнаружение = индикация вирусов в культуре клеток проводят на основе следующих феноменов: - цитопатогенного действия (ЦПД)
- 69. ЦПД = видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения от стекла), возникающие в
- 70. Виды ЦПД округление и сморщивание клеток – пикорнавирусы, нарастающая деструкция – герпесвирусы, пролиферация (образование дырок) –
- 71. ЦПД вируса полиомиелита Культура клеток до заражения ЦПД
- 74. Включения = скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом
- 75. Тельца Бабеша-Негри при бешенстве
- 77. Бляшки, или “негативные” колонии = ограниченные участки разрушенных вирусами клеток, культивируемых на питательной среде под агаровым
- 79. Бляшки под агаровым слоем
- 80. Реакция гемагглютинации (РГА) основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов за счет вирусных гликопротеиновых
- 81. Реакция гемадсорбции =РГАдс = способность культур клеток, инфицированных вирусами, адсорбировать на своей поверхности эритроциты.
- 82. Реакция иммунофлюоресценции (РИФ)
- 85. Идентификация вируса при выделении на культуре клеток РН (в т.ч. РТГАдс) РСК РИФ
- 86. Использование лабораторных животных взрослые или новорожденные белые мыши, хомяки, кролики, обезьяны применяется для выделения тех вирусов,
- 88. Способы заражения лабораторных животных интраназально, подкожно, внутримышечно, внутрибрюшинно, интрацеребрально,
- 90. Скачать презентацию
Слайд 2Основные отличия вирусов от других форм жизни
один тип нуклеиновой кислоты
отсутствие
клеточного строения
белоксинтезирующих систем
энергозапасающих
Основные отличия вирусов от других форм жизни
один тип нуклеиновой кислоты
отсутствие
клеточного строения
белоксинтезирующих систем
энергозапасающих

Слайд 3Формы существования вирусов
внеклеточная = вирион (структура) :
НК
капсид
[суперкапсид]
. Н-р, вирион имеет форму…
внутриклеточная –
Формы существования вирусов
внеклеточная = вирион (структура) :
НК
капсид
[суперкапсид]
. Н-р, вирион имеет форму…
внутриклеточная –
![Формы существования вирусов внеклеточная = вирион (структура) : НК капсид [суперкапсид] .](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/840314/slide-2.jpg)
Слайд 4Основные признаки, используемые для классификации вирусов
тип нуклеиновой кислоты (ДНК/РНК),
структура генома – количество
Основные признаки, используемые для классификации вирусов
тип нуклеиновой кислоты (ДНК/РНК),
структура генома – количество

Слайд 5Иерархическая система таксонов, применяемых в вирусологии
Царство: Vira
Подцарства: ДНК-геномные вирусы,
РНК-геномные вирусы.
Семейство
Иерархическая система таксонов, применяемых в вирусологии
Царство: Vira
Подцарства: ДНК-геномные вирусы,
РНК-геномные вирусы.
Семейство

Слайд 6КЛАССИФИКАЦИЯ ВИРУСОВ
КЛАССИФИКАЦИЯ ВИРУСОВ
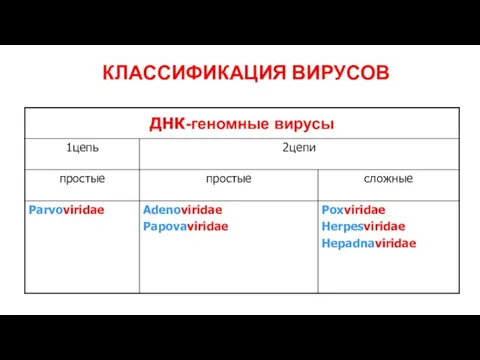
Слайд 7КЛАССИФИКАЦИЯ ВИРУСОВ
КЛАССИФИКАЦИЯ ВИРУСОВ
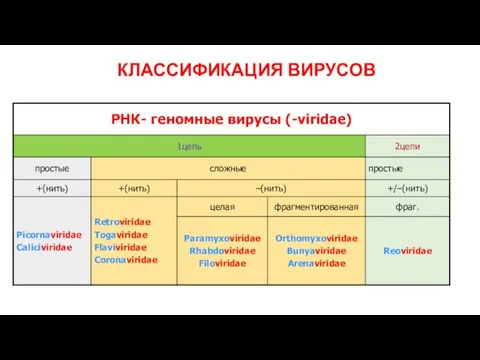
Слайд 8Принцип строения вириона
Простой:
НК+ капсид = нуклеокапсид
Сложный:
нуклеокапсид + суперкапсид
Принцип строения вириона
Простой:
НК+ капсид = нуклеокапсид
Сложный:
нуклеокапсид + суперкапсид

Слайд 9Типы симметрии капсида
спиральная
кубическая
Типы симметрии капсида
спиральная
кубическая

Слайд 10Принцип строения суперкапсида
билипидный слой
матричный белок
гликопротеины (шипы, ворсинки)
Принцип строения суперкапсида
билипидный слой
матричный белок
гликопротеины (шипы, ворсинки)
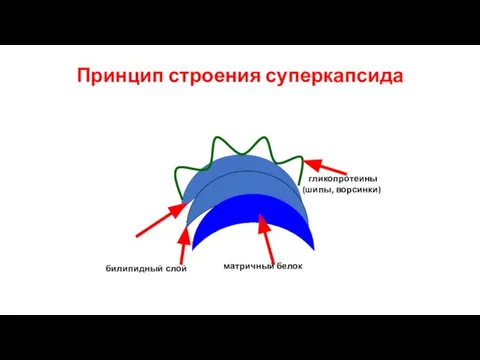
Слайд 11Форма вирионов
Форма вирионов
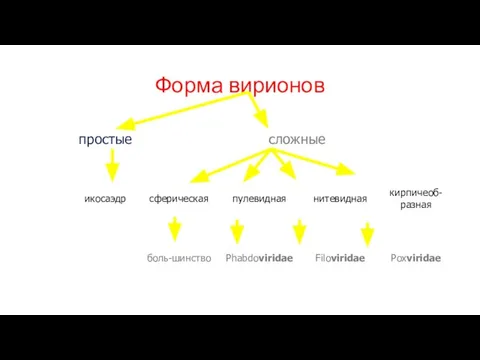
Слайд 12Общая характеристика ДНК вирионов
форма:
Линейная,
Кольцевая.
на концах – идентичные повторы:
маркеры вирусной ДНК (не клеточной),
способны
Общая характеристика ДНК вирионов
форма:
Линейная,
Кольцевая.
на концах – идентичные повторы:
маркеры вирусной ДНК (не клеточной),
способны

Слайд 13Общая характеристика РНК вирусов
форма:
Линейная,
Кольцевая.
структура:
Цельная,
Фрагментированная.
информационная функция:
+нить (позитивный геном) = иРНК (геномная РНК выполняет
Общая характеристика РНК вирусов
форма:
Линейная,
Кольцевая.
структура:
Цельная,
Фрагментированная.
информационная функция:
+нить (позитивный геном) = иРНК (геномная РНК выполняет

Слайд 14Общая характеристика белков вирусов
Структурные
Капсидные – образуют капсид,
«Внутренние», гистоноподобные – связаны с нуклеиновой
Общая характеристика белков вирусов
Структурные
Капсидные – образуют капсид,
«Внутренние», гистоноподобные – связаны с нуклеиновой

Слайд 15Схема строения простоустроенного вириона = паповавируса (вирус имеет двунитевую кольцевую ДНК)
Схема строения простоустроенного вириона = паповавируса (вирус имеет двунитевую кольцевую ДНК)

Слайд 16
Схема строения вируса гепатита А
(вирус имеет однонитевую +РНК)
Схема строения вируса гепатита А
(вирус имеет однонитевую +РНК)

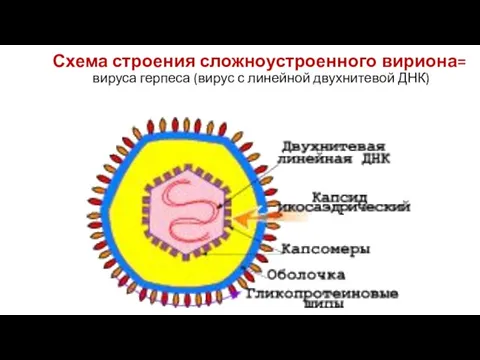
Слайд 17Схема строения сложноустроенного вириона=
вируса герпеса (вирус с линейной двухнитевой ДНК)
Схема строения сложноустроенного вириона=
вируса герпеса (вирус с линейной двухнитевой ДНК)
Слайд 18
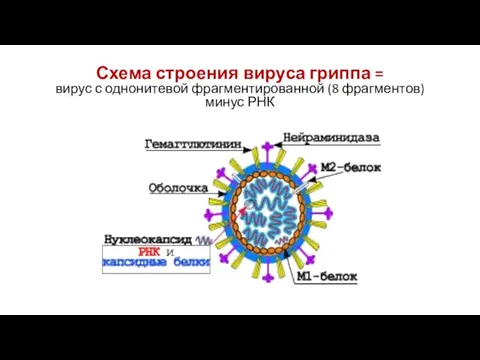
Схема строения вируса гриппа =
вирус с однонитевой фрагментированной (8 фрагментов) минус
Схема строения вируса гриппа = вирус с однонитевой фрагментированной (8 фрагментов) минус
Слайд 19Свойство вирусов = строгий цитотропизм
= Избирательность поражения вирусами определенных клеток,
= способность
Свойство вирусов = строгий цитотропизм
= Избирательность поражения вирусами определенных клеток,
= способность

Слайд 20Патологические процессы, вызываемые вирусами
инфекционные (микробные) болезни = вирусные инфекции,
Опухоли.
Патологические процессы, вызываемые вирусами
инфекционные (микробные) болезни = вирусные инфекции,
Опухоли.

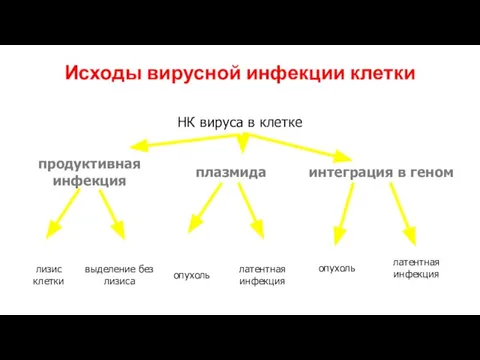
Слайд 21Исходы вирусной инфекции клетки
Исходы вирусной инфекции клетки
Слайд 22
Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный тип, при
Репродукция вирусов
Различают три типа взаимодействия вируса с клеткой:
1. продуктивный тип, при

Слайд 23Продуктивный тип взаимодействия вируса с клеткой
Этапы размножения вирусов в чувствительной клетке:
адсорбция вирионов
Продуктивный тип взаимодействия вируса с клеткой
Этапы размножения вирусов в чувствительной клетке:
адсорбция вирионов

Слайд 241. Адсорбция вирионов на клетке = прикрепление вириона к поверхности клетки:
2 фазы:
1. Адсорбция вирионов на клетке = прикрепление вириона к поверхности клетки:
2 фазы:

Слайд 252. Проникновение вируса в клетку
3 пути:
Рецептор-зависимый эндоцитоз,
Слияние оболочки вириона с клеточной мембраной,
Смешанный.
2. Проникновение вируса в клетку
3 пути:
Рецептор-зависимый эндоцитоз,
Слияние оболочки вириона с клеточной мембраной,
Смешанный.

Слайд 262.1.Проникновение вируса в клетку:
Рецептор-зависимый эндоцитоз
=захватывание и поглощение вириона клеткой:
1. Клеточная мембрана
2.1.Проникновение вируса в клетку:
Рецептор-зависимый эндоцитоз
=захватывание и поглощение вириона клеткой:
1. Клеточная мембрана

Слайд 272.2.Проникновение вируса в клетку -
слияние оболочки вириона с клеточной мембраной = виропексис:
2.2.Проникновение вируса в клетку - слияние оболочки вириона с клеточной мембраной = виропексис:

Слайд 282а.Депротеинизация вирусов= «раздевание»
= освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых)
2а.Депротеинизация вирусов= «раздевание»
= освобождение нуклеиновой кислоты путём сброса вирусом белковой (-ых)

Слайд 293. Синтез вирусных компонентов = дизъюнктивная репродукция
= синтез вирусных белков и нуклеиновых
3. Синтез вирусных компонентов = дизъюнктивная репродукция
= синтез вирусных белков и нуклеиновых

Слайд 301. Синтез вирусных белков
В зараженной клетке вирусный геном кодирует синтез 2-х групп
1. Синтез вирусных белков
В зараженной клетке вирусный геном кодирует синтез 2-х групп

Слайд 313.1.а.Синтез вирусных белков –варианты:
ДНК-содержащие вирусы:
Геномная ДНК вируса
↓
транскрипция иРНК
↓
трансляция белка вируса.
Ферменты:
- клеточная
3.1.а.Синтез вирусных белков –варианты:
ДНК-содержащие вирусы:
Геномная ДНК вируса
↓
транскрипция иРНК
↓
трансляция белка вируса.
Ферменты:
- клеточная

Слайд 323.1.б. Синтез вирусных белков -варианты:
Плюс-нитевые РНК-содержащие вирусы
= вирусный геном выполняет функцию
3.1.б. Синтез вирусных белков -варианты:
Плюс-нитевые РНК-содержащие вирусы
= вирусный геном выполняет функцию

Слайд 333.1.в. Синтез вирусных белков -варианты:
Минус-нитевые РНК-содержащие вирусы
(ортомиксовирусы, парамиксовирусы, рабдовирусы) и двунитевые
3.1.в. Синтез вирусных белков -варианты:
Минус-нитевые РНК-содержащие вирусы
(ортомиксовирусы, парамиксовирусы, рабдовирусы) и двунитевые

Слайд 343.1.г.Синтез вирусных белков -варианты:
Ретровирусы:
– геном состоит из 2-х одинаковых молекул РНК =
3.1.г.Синтез вирусных белков -варианты:
Ретровирусы:
– геном состоит из 2-х одинаковых молекул РНК =

Слайд 353.2. Репликация вирусных геномов
– зависит от типа нуклеиновой кислоты,
- наличия вирусоспецифических
3.2. Репликация вирусных геномов
– зависит от типа нуклеиновой кислоты,
- наличия вирусоспецифических

Слайд 363.2.а. Репликация вирусных геномов - варианты:
Двунитевые ДНК-вирусы
(аденовирусы, герпесвирусы, поксвирусы)
= полуконсервативный
3.2.а. Репликация вирусных геномов - варианты:
Двунитевые ДНК-вирусы
(аденовирусы, герпесвирусы, поксвирусы)
= полуконсервативный

Слайд 373.2.б.Репликация вирусных геномов
Однонитевые ДНК-вирусы
(парвовирусы)
– используют клеточные ДНК-полимеразы:
- на исходной
3.2.б.Репликация вирусных геномов
Однонитевые ДНК-вирусы
(парвовирусы)
– используют клеточные ДНК-полимеразы:
- на исходной

Слайд 383.2.в.Репликация вирусных геномов
Плюс-однонитевые РНК-вирусы
(пикорнавирусы, флавивирусы, тогавирусы, полиовирусы)
= геномная нить
3.2.в.Репликация вирусных геномов
Плюс-однонитевые РНК-вирусы
(пикорнавирусы, флавивирусы, тогавирусы, полиовирусы)
= геномная нить

Слайд 393.2.г.Репликация вирусных геномов
Минус-однонитевые РНК-вирусы
(Рабдовирусы, парамиксовирусы, ортомиксовирусы)
– имеют РНК-зависимую
3.2.г.Репликация вирусных геномов
Минус-однонитевые РНК-вирусы
(Рабдовирусы, парамиксовирусы, ортомиксовирусы)
– имеют РНК-зависимую

Слайд 403.2.д.Репликация вирусных геномов
Двунитевые РНК-вирусы
(реовирусы, ротавирусы)
– как у минус
3.2.д.Репликация вирусных геномов
Двунитевые РНК-вирусы
(реовирусы, ротавирусы)
– как у минус

Слайд 413.2.е.Репликация вирусных геномов
Ретровирусы
= плюс-нитевые диплоидные РНК-содержащие вирусы, имеют обратную
3.2.е.Репликация вирусных геномов
Ретровирусы
= плюс-нитевые диплоидные РНК-содержащие вирусы, имеют обратную

Слайд 424.Формирование вирусов
Происходит путем самосборки = составные части вируса транспортируются в определенный участок
4.Формирование вирусов
Происходит путем самосборки = составные части вируса транспортируются в определенный участок

Слайд 43
5.Выход вирусов из клетки
1. взрывной путь: клетка погибает и вирусы выходят наружу
5.Выход вирусов из клетки
1. взрывной путь: клетка погибает и вирусы выходят наружу

Слайд 44Абортивный тип взаимодействия вируса с клеткой
= прерывание инфекционного процесса в клетке на
Абортивный тип взаимодействия вируса с клеткой
= прерывание инфекционного процесса в клетке на

Слайд 45Интегративный тип взаимодействия вируса с клеткой = вирогения
= нуклеиновая кислота вируса встраивается
Интегративный тип взаимодействия вируса с клеткой = вирогения
= нуклеиновая кислота вируса встраивается

Слайд 46
Значение вирогении
1. Сохранение вирусной информации в составе клеточного генома = персистенция:
→ клетка
Значение вирогении
1. Сохранение вирусной информации в составе клеточного генома = персистенция:
→ клетка

Слайд 47Исходы активации персистирующего вируса
рецидив того же заболевания,
развитие другого заболевания, вызываемого тем же
Исходы активации персистирующего вируса
рецидив того же заболевания,
развитие другого заболевания, вызываемого тем же

Слайд 48Способы культивирования вирусов
3 модели:
куриный эмбрион
культура клеток
организм лабораторного животного
⇓
обнаружение наличия вируса
(индикация)
⇓
определение типа вируса
(идентификация)
Способы культивирования вирусов
3 модели:
куриный эмбрион
культура клеток
организм лабораторного животного
⇓
обнаружение наличия вируса
(индикация)
⇓
определение типа вируса
(идентификация)

Слайд 49Использование для вирусологического метода куриного эмбриона
Использование для вирусологического метода куриного эмбриона

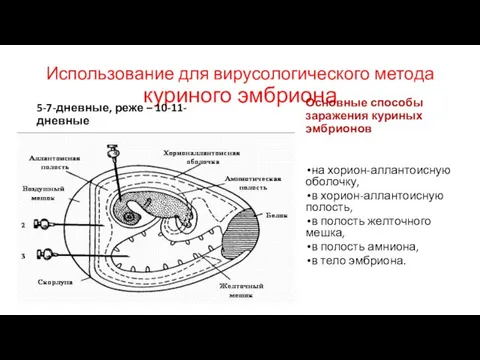
Слайд 50Использование для вирусологического метода куриного эмбриона
5-7-дневные, реже – 10-11-дневные
Основные способы заражения куриных
Использование для вирусологического метода куриного эмбриона
5-7-дневные, реже – 10-11-дневные
Основные способы заражения куриных

Слайд 52Обнаружение вирусов в курином эмбрионе
индикация:
гибель эмбриона,
морфологические изменения эмбриона/оболочек,
РГА с жидкостью из полостей
Обнаружение вирусов в курином эмбрионе
индикация:
гибель эмбриона,
морфологические изменения эмбриона/оболочек,
РГА с жидкостью из полостей

Слайд 53Использование культур клеток
Культуры клеток = соматические или эмбриональные клетки человека или животных,
Использование культур клеток
Культуры клеток = соматические или эмбриональные клетки человека или животных,


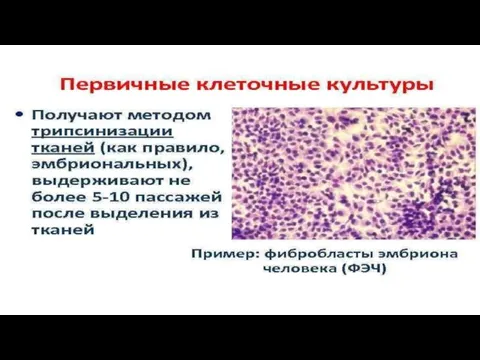
Слайд 55Первичные (эмбриональные) культуры клеток
получают из тканей (эмбриональных или нормальных) многоклеточных организмов.
Такие
Первичные (эмбриональные) культуры клеток
получают из тканей (эмбриональных или нормальных) многоклеточных организмов.
Такие

Слайд 57Перевиваемые культуры клеток
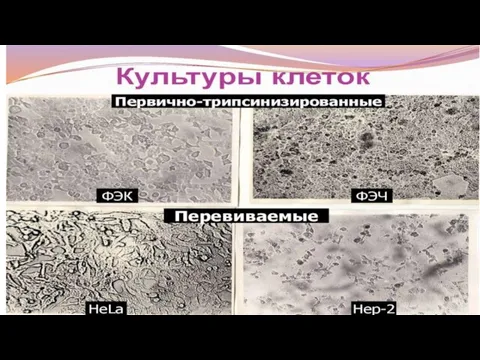
Перевиваемые = стабильные = готовят из опухолевых клеток, способных длительно
Перевиваемые культуры клеток
Перевиваемые = стабильные = готовят из опухолевых клеток, способных длительно

Слайд 59 Преимущества перевиваемых культур клеток перед первичными:
продолжительность культивирования – десятки лет,
высокая скорость
Преимущества перевиваемых культур клеток перед первичными:
продолжительность культивирования – десятки лет,
высокая скорость

Слайд 60Полуперевиваемые культуры клеток
= диплоидные клетки различных тканей и органов, способные к ограниченному
Полуперевиваемые культуры клеток
= диплоидные клетки различных тканей и органов, способные к ограниченному


Слайд 62
Условия культивирования клеток:
Использование лабораторной посуды из нейтрального стекла – пробирки, флаконы, матрасы
Условия культивирования клеток:
Использование лабораторной посуды из нейтрального стекла – пробирки, флаконы, матрасы






Слайд 68 Обнаружение = индикация вирусов в культуре клеток
проводят на основе следующих феноменов:
Обнаружение = индикация вирусов в культуре клеток
проводят на основе следующих феноменов:

Слайд 69
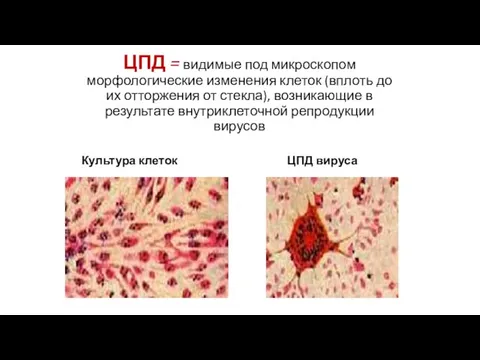
ЦПД = видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения
ЦПД = видимые под микроскопом морфологические изменения клеток (вплоть до их отторжения
Слайд 70Виды ЦПД
округление и сморщивание клеток – пикорнавирусы,
нарастающая деструкция – герпесвирусы,
пролиферация (образование дырок)
Виды ЦПД
округление и сморщивание клеток – пикорнавирусы,
нарастающая деструкция – герпесвирусы,
пролиферация (образование дырок)

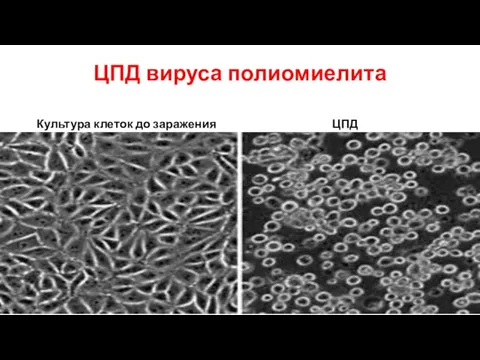
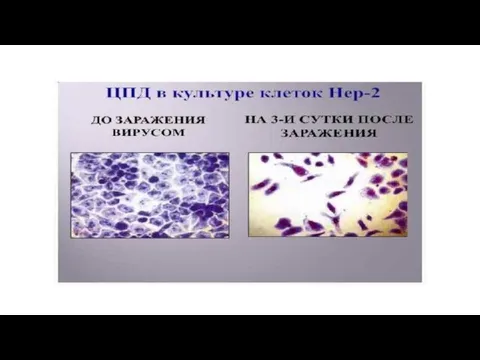
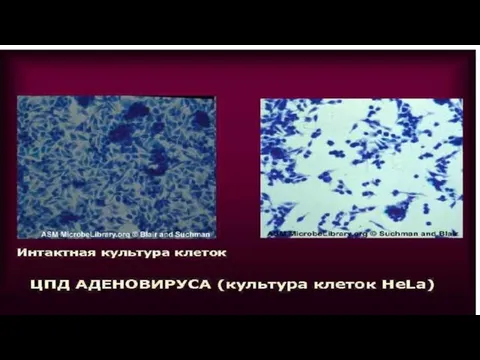
Слайд 71ЦПД вируса полиомиелита
Культура клеток до заражения
ЦПД
ЦПД вируса полиомиелита
Культура клеток до заражения
ЦПД
Слайд 74Включения
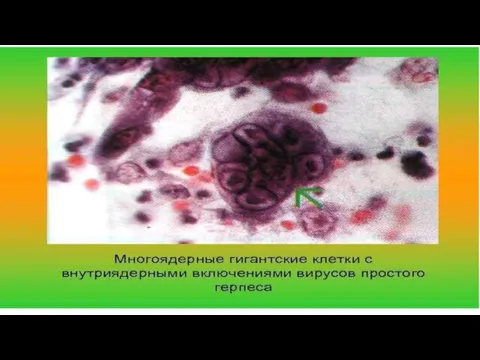
= скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток,
Включения
= скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток,

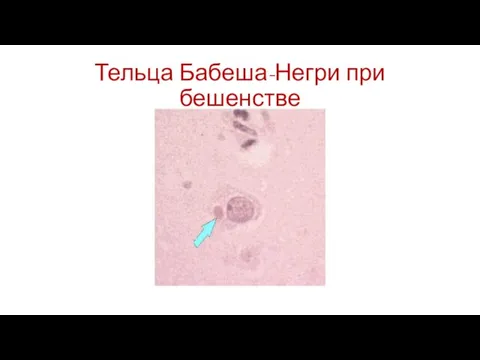
Слайд 75Тельца Бабеша-Негри при бешенстве
Тельца Бабеша-Негри при бешенстве
Слайд 77Бляшки, или “негативные” колонии
= ограниченные участки разрушенных вирусами клеток, культивируемых на питательной
Бляшки, или “негативные” колонии
= ограниченные участки разрушенных вирусами клеток, культивируемых на питательной

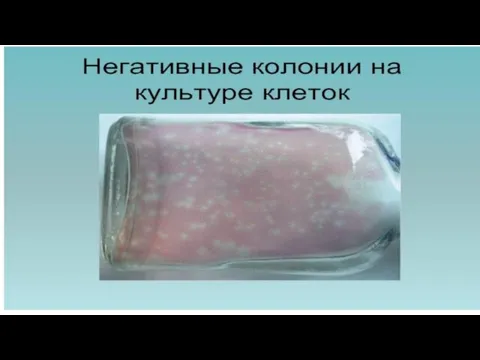
Слайд 79Бляшки под агаровым слоем
Бляшки под агаровым слоем

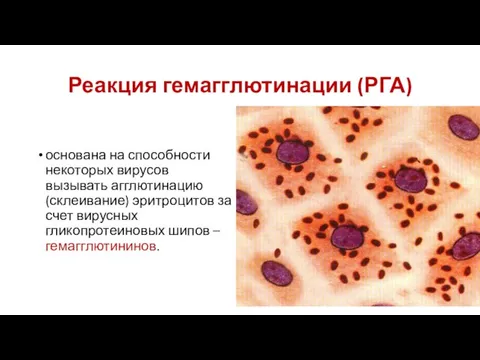
Слайд 80Реакция гемагглютинации (РГА)
основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов за
Реакция гемагглютинации (РГА)
основана на способности некоторых вирусов вызывать агглютинацию (склеивание) эритроцитов за
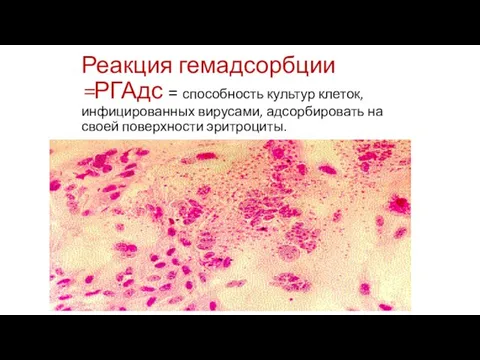
Слайд 81
Реакция гемадсорбции =РГАдс = способность культур клеток, инфицированных вирусами, адсорбировать на своей
Реакция гемадсорбции =РГАдс = способность культур клеток, инфицированных вирусами, адсорбировать на своей
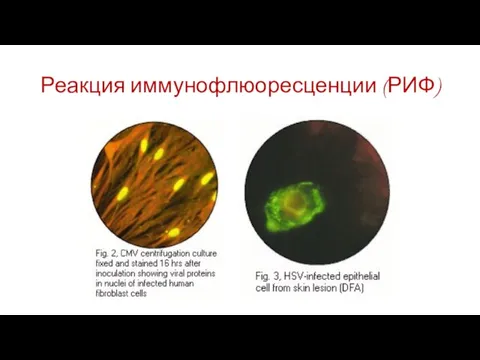
Слайд 82Реакция иммунофлюоресценции (РИФ)
Реакция иммунофлюоресценции (РИФ)


Слайд 85Идентификация вируса при выделении на культуре клеток
РН (в т.ч. РТГАдс)
РСК
РИФ
Идентификация вируса при выделении на культуре клеток
РН (в т.ч. РТГАдс)
РСК
РИФ

Слайд 86Использование лабораторных животных
взрослые или новорожденные белые мыши, хомяки, кролики, обезьяны
применяется для выделения
Использование лабораторных животных
взрослые или новорожденные белые мыши, хомяки, кролики, обезьяны
применяется для выделения


Слайд 88Способы заражения лабораторных животных
интраназально,
подкожно,
внутримышечно,
внутрибрюшинно,
интрацеребрально,
Способы заражения лабораторных животных
интраназально,
подкожно,
внутримышечно,
внутрибрюшинно,
интрацеребрально,

 Обобщающий урок Ткани животных
Обобщающий урок Ткани животных Как живут растения. Окружающий мир (1 класс)
Как живут растения. Окружающий мир (1 класс) Sprache und Gehirn. Anatomische Grundlagen
Sprache und Gehirn. Anatomische Grundlagen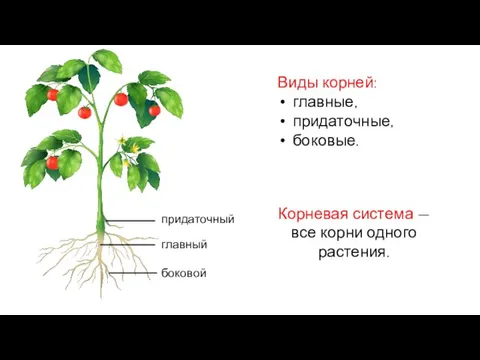 Виды корней
Виды корней Биологическое действие шума
Биологическое действие шума Динозавры и их жизнь
Динозавры и их жизнь Озиқ-овқат маҳсулотлари товаршунослиги
Озиқ-овқат маҳсулотлари товаршунослиги Презентация на тему Приспособленность организмов
Презентация на тему Приспособленность организмов  Рыбы Якутии
Рыбы Якутии Побег
Побег Болезни финиковых пальм
Болезни финиковых пальм Виды цветов
Виды цветов Фенофазы развития
Фенофазы развития Растения Самбурга
Растения Самбурга Общая экология. РИПК-6
Общая экология. РИПК-6 Причины нарушения в развитии организмов
Причины нарушения в развитии организмов Взаимоотношения организма и среды. Невидимые нити природы
Взаимоотношения организма и среды. Невидимые нити природы Новые/сложные задания в сборнике Рохлова
Новые/сложные задания в сборнике Рохлова Genetic algorithm. Phenotype
Genetic algorithm. Phenotype Органикалық дүниенің тарихи дамуы. Архей. Протерозой
Органикалық дүниенің тарихи дамуы. Архей. Протерозой Лекция №8. Тепловые свойства почв
Лекция №8. Тепловые свойства почв Аттестационная работа. Биометрическое исследование влияния дерматоглифических особенностей человека
Аттестационная работа. Биометрическое исследование влияния дерматоглифических особенностей человека Глубоководные обитатели
Глубоководные обитатели Энергетический обмен
Энергетический обмен 1_2_Metody_tsitologii_Kletochnaya_teoria_Pro_i_Eukarioty_Sravnenie_Tsarstv_RZhG (1)
1_2_Metody_tsitologii_Kletochnaya_teoria_Pro_i_Eukarioty_Sravnenie_Tsarstv_RZhG (1) Ябык орлыклы үсемлекләр
Ябык орлыклы үсемлекләр Дыхание растений. Тест
Дыхание растений. Тест Животные. Животный мир тайги
Животные. Животный мир тайги