- Строение синапса и его медиаторы. Виды синапсов

Содержание
- 2. Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством специального структурного образования —
- 3. Понятие и физиология химических и электрических синапсов Синапс — это специализированная структура, обеспечивающая межклеточную передачу сигналов
- 4. Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка — это синапс между нейронами,
- 5. Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
- 6. Виды синапсов Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала. По месту положения выделяют нервно-мышечные
- 7. Таблица 1. Классификация и виды синапсов
- 8. Классификация синапсов и механизм передачи возбуждения Синапсы классифицируют следующим образом: по местоположению — периферические и центральные;
- 9. Свойства медиаторов Синтезируются в нейроне Накапливаются в окончании клетки Выделяются при появлении иона Са2+ в пресинаптическом
- 10. Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется, если последующий сигнал приходит
- 11. Электрические синапсы Кроме синапсов с химической передачей возбуждения в организме есть синапсы с электрической передачей. Эти
- 12. Электрические синапсы образуются между клетками, формирующими между мембранами плотные щелевые контакты. Ширина щели составляет около 3
- 13. Электрические синапсы широко распространены в нервной системе беспозвоночных, а у млекопитающих встречаются крайне редко. Вместе с
- 14. Благодаря наличию каналов, размеры которых позволяют переходить из клетки в клетку неорганическим ионам и даже небольшим
- 15. Химические синапсы Физиологические свойства химических синапсов Синапсы с химической передачей возбуждения обладают определенными свойствами: возбуждение проводится
- 16. Химические синапсы образуются специализированными структурами двух клеток в области их контакта (рис. 2). Одной из этих
- 17. Передача информации в химических синапсах осуществляется через синаптическую щель — область внеклеточного пространства шириной 10-50 нм,
- 18. Характеристика химического синапса Принцип «физиологического клапана» При участии посредника-медиатора Синаптическая задержка Принцип Дейла Трансформация ритма возбуждения
- 19. Передача информации в химических синапсах При поступлении потенциала действия к пресинаптическому окончанию происходит деполяризация пресинаптической мембраны
- 20. ПКП возникает в нервно-мышечных синапсах, в остальных — возбуждающий постсинаптический потенциал (ВПСП) или тормозной постсинаптический потенциал
- 21. Медиаторы — это биологически активные вещества, посредством которых осуществляются межклеточные взаимодействия в синапсах. К ним относятся
- 22. Нервно-мышечный синапс Особенности передачи сигналов в нервно-мышечном синапсе Нервно-мышечный синапс образован окончанием аксона моторного нейрона и
- 23. Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и постсинантическая мембрана, являющаяся частью
- 24. Между пре- и постсинаптическими мембранами располагается синаптическая щель шириной 50-100 им. Она заполнена межклеточным веществом и
- 25. Рис. 2. Структура нервно-мышечного синапса в покое и при активации
- 26. При поступлении к аксонной терминали одного ПД в синаптическую щель выбрасывается ацетилхолин из десятков синаптических пузырьков.
- 27. Положительно заряженные ионы Na+, входящие в мышечное волокно, деполяризуют постсинаптическую мембрану, нейтрализуя часть отрицательных зарядов на
- 28. Почему же потенциал действия возникает на прилежащей к синапсу мембране, а не па постсинаптической мембране? Постсинаптическая
- 29. Когда содержание медиатора понизится до уровня 10 нмолей, ацетилхолин диссоциирует из связи с рецептором, восстанавливается способность
- 30. Имеется много факторов, способных влиять на состояние синаптической передачи сигналов к скелетным мышцам. Это влияние можно
- 31. Следующий способ воздействия на синаптическую передачу также связан с влиянием на количество ацетилхолина в синаптической щели
- 32. Ряд веществ может легко связываться с никотинчувствительными холинорецепторами и блокировать при этом открытие лигандзависимых ионных каналов.
- 35. Скачать презентацию
Слайд 2Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством
Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством

Слайд 3Понятие и физиология химических и электрических синапсов
Синапс — это специализированная структура, обеспечивающая межклеточную
Понятие и физиология химических и электрических синапсов
Синапс — это специализированная структура, обеспечивающая межклеточную

Слайд 4Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка —
Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка —
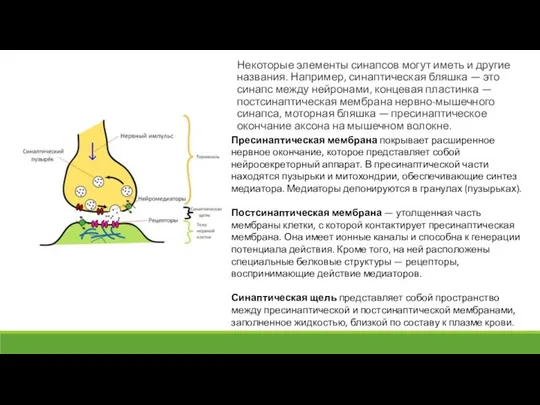
Слайд 5Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Слайд 6Виды синапсов
Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По месту положения выделяют
Виды синапсов
Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По месту положения выделяют

Слайд 7Таблица 1. Классификация и виды синапсов
Таблица 1. Классификация и виды синапсов

Слайд 8Классификация синапсов и механизм передачи возбуждения
Синапсы классифицируют следующим образом:
по местоположению — периферические
Классификация синапсов и механизм передачи возбуждения
Синапсы классифицируют следующим образом:
по местоположению — периферические

Слайд 9Свойства медиаторов
Синтезируются в нейроне
Накапливаются в окончании клетки
Выделяются при появлении иона Са2+ в
Свойства медиаторов
Синтезируются в нейроне
Накапливаются в окончании клетки
Выделяются при появлении иона Са2+ в

Слайд 10Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется,
Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется,

Слайд 11Электрические синапсы
Кроме синапсов с химической передачей возбуждения в организме есть синапсы с
Электрические синапсы
Кроме синапсов с химической передачей возбуждения в организме есть синапсы с

Слайд 12Электрические синапсы образуются между клетками, формирующими между мембранами плотные щелевые контакты. Ширина щели
Электрические синапсы образуются между клетками, формирующими между мембранами плотные щелевые контакты. Ширина щели

Слайд 13Электрические синапсы широко распространены в нервной системе беспозвоночных, а у млекопитающих встречаются
Электрические синапсы широко распространены в нервной системе беспозвоночных, а у млекопитающих встречаются

Слайд 14Благодаря наличию каналов, размеры которых позволяют переходить из клетки в клетку неорганическим
Благодаря наличию каналов, размеры которых позволяют переходить из клетки в клетку неорганическим

Слайд 15Химические синапсы
Физиологические свойства химических синапсов
Синапсы с химической передачей возбуждения обладают определенными свойствами:
возбуждение
Химические синапсы
Физиологические свойства химических синапсов
Синапсы с химической передачей возбуждения обладают определенными свойствами:
возбуждение

Слайд 16Химические синапсы образуются специализированными структурами двух клеток в области их контакта (рис. 2).
Химические синапсы образуются специализированными структурами двух клеток в области их контакта (рис. 2).

Слайд 17Передача информации в химических синапсах осуществляется через синаптическую щель — область внеклеточного
Передача информации в химических синапсах осуществляется через синаптическую щель — область внеклеточного

Слайд 18Характеристика химического синапса
Принцип «физиологического клапана»
При участии посредника-медиатора
Синаптическая задержка
Принцип Дейла
Трансформация ритма возбуждения
Синаптическое облегчение
Характеристика химического синапса
Принцип «физиологического клапана»
При участии посредника-медиатора
Синаптическая задержка
Принцип Дейла
Трансформация ритма возбуждения
Синаптическое облегчение
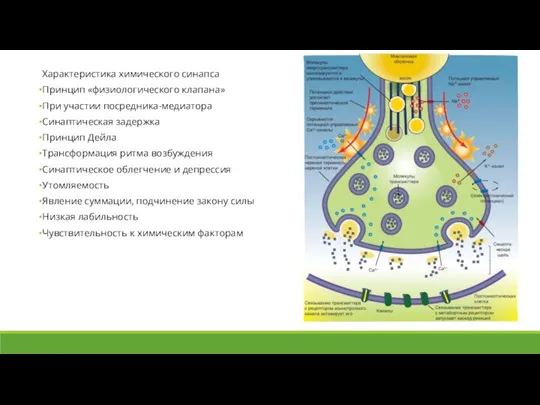
Слайд 19Передача информации в химических синапсах
При поступлении потенциала действия к пресинаптическому окончанию происходит
Передача информации в химических синапсах
При поступлении потенциала действия к пресинаптическому окончанию происходит

Слайд 20ПКП возникает в нервно-мышечных синапсах, в остальных — возбуждающий постсинаптический потенциал (ВПСП)
ПКП возникает в нервно-мышечных синапсах, в остальных — возбуждающий постсинаптический потенциал (ВПСП)

Слайд 21Медиаторы — это биологически активные вещества, посредством которых осуществляются межклеточные взаимодействия в синапсах.
Медиаторы — это биологически активные вещества, посредством которых осуществляются межклеточные взаимодействия в синапсах.

Слайд 22Нервно-мышечный синапс
Особенности передачи сигналов в нервно-мышечном синапсе
Нервно-мышечный синапс образован окончанием аксона моторного
Нервно-мышечный синапс
Особенности передачи сигналов в нервно-мышечном синапсе
Нервно-мышечный синапс образован окончанием аксона моторного

Слайд 23Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и
Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и

Слайд 24Между пре- и постсинаптическими мембранами располагается синаптическая щель шириной 50-100 им. Она
Между пре- и постсинаптическими мембранами располагается синаптическая щель шириной 50-100 им. Она

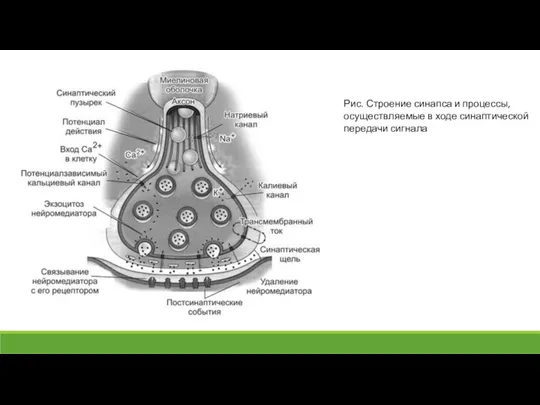
Слайд 25Рис. 2. Структура нервно-мышечного синапса в покое и при активации
Рис. 2. Структура нервно-мышечного синапса в покое и при активации
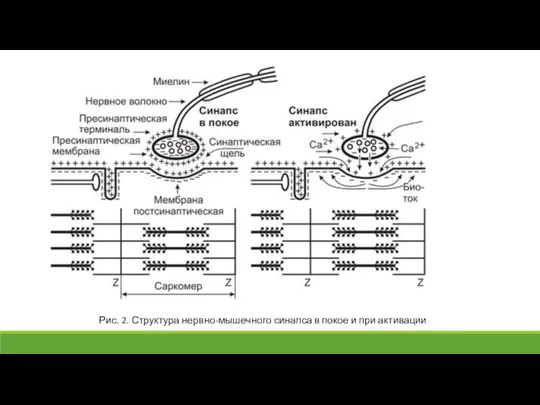
Слайд 26При поступлении к аксонной терминали одного ПД в синаптическую щель выбрасывается ацетилхолин
При поступлении к аксонной терминали одного ПД в синаптическую щель выбрасывается ацетилхолин

Слайд 27Положительно заряженные ионы Na+, входящие в мышечное волокно, деполяризуют постсинаптическую мембрану, нейтрализуя
Положительно заряженные ионы Na+, входящие в мышечное волокно, деполяризуют постсинаптическую мембрану, нейтрализуя

Слайд 28Почему же потенциал действия возникает на прилежащей к синапсу мембране, а не
Почему же потенциал действия возникает на прилежащей к синапсу мембране, а не

Слайд 29Когда содержание медиатора понизится до уровня 10 нмолей, ацетилхолин диссоциирует из связи
Когда содержание медиатора понизится до уровня 10 нмолей, ацетилхолин диссоциирует из связи

Слайд 30Имеется много факторов, способных влиять на состояние синаптической передачи сигналов к скелетным
Имеется много факторов, способных влиять на состояние синаптической передачи сигналов к скелетным

Слайд 31Следующий способ воздействия на синаптическую передачу также связан с влиянием на количество
Следующий способ воздействия на синаптическую передачу также связан с влиянием на количество

Слайд 32Ряд веществ может легко связываться с никотинчувствительными холинорецепторами и блокировать при этом
Ряд веществ может легко связываться с никотинчувствительными холинорецепторами и блокировать при этом


 Типы яйцеклеток
Типы яйцеклеток Жилища животных
Жилища животных !!!Биология как наука
!!!Биология как наука Транспорт веществ через мембрану
Транспорт веществ через мембрану Лошадки - удивительные животные
Лошадки - удивительные животные Анатомия из пластелина
Анатомия из пластелина Синдром Дауна
Синдром Дауна Путешествие во времени, учебное занятие. Эволюция органического мира
Путешествие во времени, учебное занятие. Эволюция органического мира Анализаторы человека
Анализаторы человека Легенды и загадки о цветах
Легенды и загадки о цветах Интеллект-карта. Пресмыкающиеся – наземные позвоночные животные
Интеллект-карта. Пресмыкающиеся – наземные позвоночные животные Органы дыхательной системы человека
Органы дыхательной системы человека Класс Паукообразные (Arachnida)
Класс Паукообразные (Arachnida) Анатомия дыхательной системы
Анатомия дыхательной системы Сохраним природу
Сохраним природу История возникновения ВИЧ. Биологические аспекты ВИЧ. Развитие ВИЧ-инфекции
История возникновения ВИЧ. Биологические аспекты ВИЧ. Развитие ВИЧ-инфекции Строение бактерий
Строение бактерий Свет в жизни живых организмов
Свет в жизни живых организмов Основы вирусологии. Бактериофагия
Основы вирусологии. Бактериофагия Строение и работа сердца человека
Строение и работа сердца человека Химический состав пищи (урок № 8)
Химический состав пищи (урок № 8) Қияр өсіруді бақылау
Қияр өсіруді бақылау Простейшие
Простейшие Листья деревьев и кустарников
Листья деревьев и кустарников Презентация на тему Экономика природопользования
Презентация на тему Экономика природопользования  Законы Менделя
Законы Менделя Сравнение процесса деления митоза и мейоза и их биологическое значение
Сравнение процесса деления митоза и мейоза и их биологическое значение В гости к пернатым друзьям
В гости к пернатым друзьям