- Связь физиологии ЦНС с психологией

Содержание
- 2. Нервная система - координатор деятельности всех органов и систем живого организма. Она воспринимает стимулы (раздражители) от
- 3. 1. Основные понятия физиологии. Принципы организации физиологических функций. ФИЗИОЛОГИЯ (от лат. phisis – природа, logos –
- 4. ОСНОВНЫЕ ПОНЯТИЯ ФИЗИОЛОГИИ Физиологическая система – наследственно закрепленная совокупность органов и тканей, выполняющая общую функцию, иногда
- 5. Учение о функциональных системах в живых организмах создано академиком П. К. Анохиным (1935). Функциональная система –
- 6. Достижение полезного приспособительного результата возможно на основе регуляции Регуляция – это комплекс процессов, обеспечивающих приспособление уровня
- 7. Конечный полезный приспособительный результат – это поддержание некоего постоянства внутренней среды организма - гомеостаза. Гомеостаз -
- 8. Нейрофизиология - специальный раздел физиологии, изучающий функции нервной системы и ее основных структурных единиц – нейронов.
- 9. ПРЕДМЕТ НЕЙРОФИЗИОЛОГИИ функционирование нервной системы и нейрона ЗАДАЧИ закономерности функционирования центральной нервной системы (ЦНС), нейронная организация
- 10. МЕТОДЫ метод наблюдения; метод экстирпации метод поперечных перерезок метод раздражения стереотаксическая методика метод условных рефлексов нейрофизиологическое
- 11. Физиологические свойства нервных клеток: Раздражимость – способность живых клеток реагировать на воздействия внешней среды Возбудимость –
- 12. Раздражимость – это универсальное свойство всех биологических систем (от клетки до популяционного уровня) отвечать на любое
- 13. Все ткани организма в зависимости от свойств клеточных мембран делятся на Электровозбудимые; Хемовозбудимые; Механовозбудимые. В основе
- 14. 3. Возбудимые ткани и их свойства
- 15. Возбуждение – такое состояние возбудимой ткани, которое характеризуется быстрым колебанием электрического потенциала клеточной мембраны, что проявляется
- 16. Раздражимость. Типы Раздражителей. Любая живая ткань может находиться как в состоянии покоя, так и в деятельном
- 17. Раздражитель – любое изменение внешней среды или внутреннего состояния организма, воспринимаемое клетками и вызывающее ответную реакцию.
- 18. Химические соединения: образующиеся в организме гормоны, метаболиты поступающие извне кислоты, щелочи, лекарственные препараты яды КЛАССИФИКАЦИЯ РАЗДРАЖИТЕЛЕЙ
- 19. По физиологическому значению Адекватные Неадекватные Раздражитель, к которому клетка в процессе эволюции приобрела наибольшую чувствительность вследствие
- 20. Схема строения клеточной мембраны: молекула липида; липидный бислой; интегральные белки; полуинтегральные белки; периферические белки; гликокаликс; субмембранный
- 21. Функции мембран Мембрана – граница между внутренней и внешней средами клетки, отличающимися по физико-химическим показателям; Участие
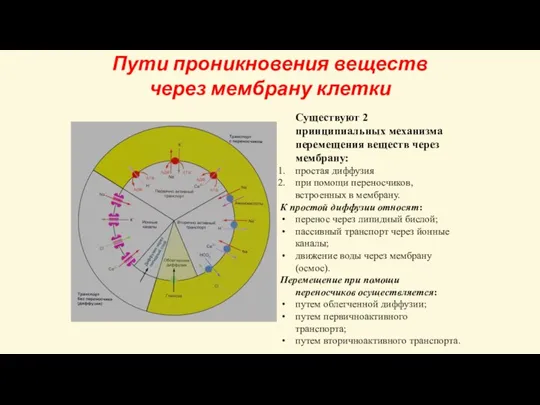
- 22. Пути проникновения веществ через мембрану клетки Существуют 2 принципиальных механизма перемещения веществ через мембрану: простая диффузия
- 23. Транспортные системы клетки
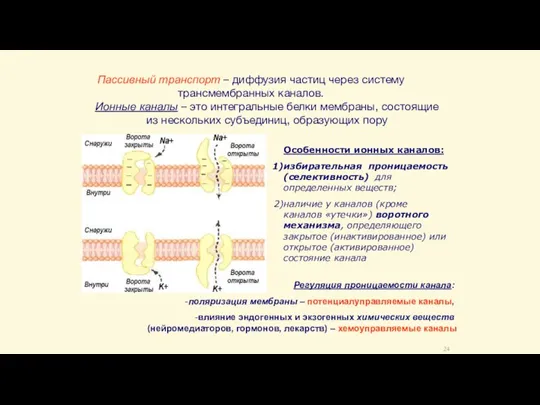
- 24. Пассивный транспорт – диффузия частиц через систему трансмембранных каналов. Ионные каналы – это интегральные белки мембраны,
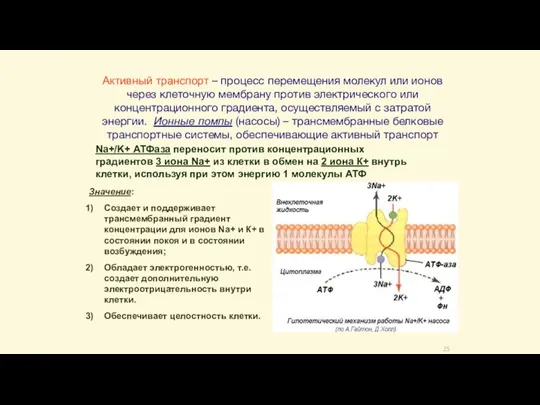
- 25. Активный транспорт – процесс перемещения молекул или ионов через клеточную мембрану против электрического или концентрационного градиента,
- 26. Основные виды электрических ответов нервных клеток: Локальный ответ; Потенциал действия; Возбуждающий и тормозный постсинаптические потенциалы; Генераторный
- 27. Мембранно-ионная теория. Природа потенциала покоя 1902 г. - И. Бернштейн сформулировал основные положения мембранно-ионной теории возбудимости
- 28. Потенциал покоя ПП - это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка
- 29. Потенциал покоя создается Во-первых, за счет работы АТФ-зависимого Na+/K+ насоса (асимметричного обмена Na+ на K+ в
- 30. Механизмы формирования градиентов: пассивный транспорт – простая диффузия ионов через мембрану клетки, активный транспорт – специализированные
- 31. АТФ-зависимый Na+/K+ насос
- 32. Основные положения мембранно-ионной теории возбуждения 1. В покое мембрана клетки находится в состоянии статической поляризации, которая
- 33. Порог возбудимости аксонного холмика - 10 мВ, тела клетки - 50 мВ; Величина потенциала покоя составляет
- 34. В процессе возбуждения происходит быстрое (в течение долей секунды) изменение проницаемости мембраны для ионов Мембрана становится
- 36. Локальный ответ ПД ЛО При действии допороговых раздражителей (сила раздражителя меньше пороговой) на мембране возникает местная
- 37. Потенциал действия ПД - быстрое колебание мембранного потенциала, возникающее при возбуждении нервных и мышечных клеток (волокон),
- 38. Потенциал действия Потенциал действия (ПД) – это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала вследствие
- 39. Механизм ПД ВОСХОДЯЩАЯ ФАЗА ПД: раздражитель пороговой или сверхпороговой силы увеличивает проницаемость мембраны для ионов Na+;
- 40. Механизм ПД
- 41. Свойства потенциала действия Возникает в ответ на действие пороговых и сверхпороговых раздражителей (подчиняется закону «всё или
- 42. Фазы потенциала действия Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный
- 43. График ПД – местная деполяризация (локальный ответ) – распространяющаяся деполяризация, восходящая часть «спайк»-потенциала – овершут (инверсия)
- 48. Закон «всё-или-ничего» Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо
- 49. Проводимость нейрона Скорость проведения по нейрону зависит от строения нервного волокна (наличия, отсутствия миелиновой оболочки) и
- 50. Распространение потенциала действия по немиелинизированным волокнам По немиелинизированному волокну ПД распространяется непрерывно. Возбуждение распространяется за счет
- 51. Распространение потенциала действия по немиелинизированным волокнам
- 52. Распространение потенциала действия по миелинизированным волокнам В миелиновых волокнах возбуждение охватывает только участки перехватов Ранвье, т.е.
- 54. Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами
- 55. Электрический синапс Электрический синапс представляет собой щелевидное образование (размеры щели 2-4 нм) с ионными мостиками-каналами (коннексонами)
- 56. Работа электрического синапса Локальные круги тока протекающего между деполяризованной и недеполяризованной областями, показаны стрелками
- 57. Строение химического синапса
- 58. Нейромедиаторы Ацетилхолин Моноамины Аминокислоты Полипептиды
- 59. Схема экзоцитоза.
- 60. Постсинаптические потенциалы Возбуждающее воздействие – это такое изменение состояния нейрона, при котором облегчается возможность передачи возбуждения
- 61. Торможение в ЦНС – это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Роль
- 62. Центральное торможение Торможение вслед за возбуждением Пессимальное Пресинаптическое Реципрокное Возвратное Латеральное Прямое Первичное Вторичное Постсинаптическое
- 63. Торможение. Торможение – самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения. Торможение может
- 64. Пресинаптическое торможение Развивается в пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. Структурной основой
- 65. Постсинаптическое торможение Медиатор, выделяемый постсинаптическими окончаниями тормозных синапсов, изменяет свойства постсинаптической мембраны таким образом, что способность
- 66. Виды первичного торможения (ГАМК, глицин) Пресинаптическое Постсинаптическое Прямое
- 67. Торможение в ЦНС В интегративной деятельности центральной нервной системы важная роль принадлежит тормозным нейронным сетям. Выделяют
- 68. Возвратное торможение Если тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют, то
- 69. Реципрокное торможение Этот вид торможения широко представлен в спинном мозге. С помощью него происходит автоматическое взаимодействие
- 70. Латеральное торможение Данный вид торможения широко представлен в структурах нервной системы, имеющих слоистое строение. Латеральное торможение
- 71. Виды торможения
- 72. Нервный центр – это функциональное объединение нейронов, принимающих участие в осуществлении одной функции.
- 73. Нервный центр Нервным центром называется совокупность нейронов в ЦНС, обеспечивающих регуляцию какой-либо функции организма. Не следует
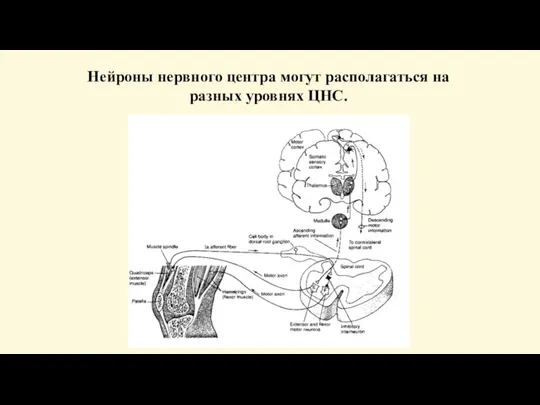
- 74. Нейроны нервного центра могут располагаться на разных уровнях ЦНС.
- 75. Обусловлены: Свойствами входящих в его состав нейронов Свойствами синаптических образований Особенностями структурной организации центра Свойства нервных
- 76. Свойства нервных центров Одностороннее проведение возбуждения - возбуждение передается с афферентного на эфферентный нейрон. Причина: химические
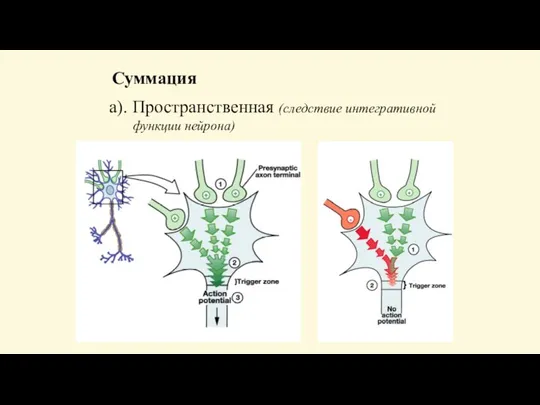
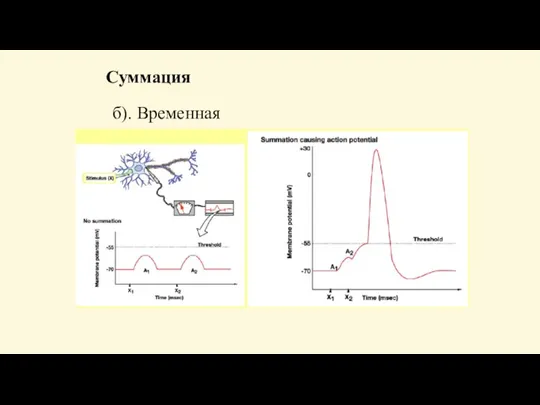
- 77. Имеется 2 вида суммации: временная и пространственная. Временная - возникает ответная реакция при действии нескольких следующих
- 78. Суммация а). Пространственная (следствие интегративной функции нейрона)
- 79. Суммация б). Временная
- 80. Посттетаническая потенция - усиление ответной реакции, наблюдается после серии нервных импульсов. Механизм: потенциация возбуждения в синапсах
- 81. Посттетаническая потенциация
- 82. Трансформация ритма возбуждения - несоответствие ответной реакции частоте наносимых раздражений. На афферентном нейроне происходит трансформация в
- 83. Свойства нервных центров Трансформация ритма возбуждения.
- 84. Свойства нервных центров Явление окклюзии отражает эффект взаимодействия между собой двух импульсных потоков, при котором имеет
- 85. Принцип окклюзии
- 86. Свойства нервных центров Явление облегчения, которое по своему внешнему проявлению противоположно окклюзии. 0но проявляется в том,
- 87. Центральное облегчение Объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный центр иннервирует определенное
- 88. Принцип пространственного облегчения
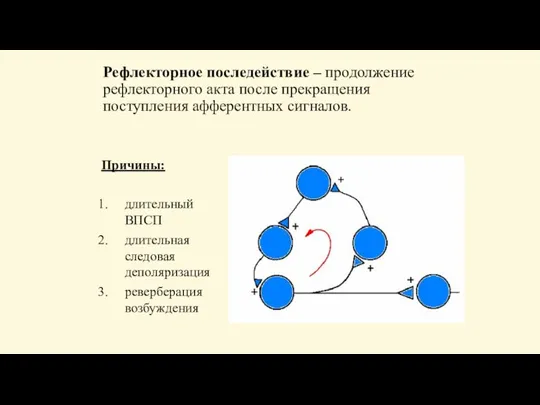
- 89. Рефлекторное последействие - продолжение ответной реакции после прекращения действия раздражителя: кратковременное последействие - в течение нескольких
- 90. Свойства нервных центров Последействие. Это одно из свойств, характерных для нейронных цепей. Оно заключается в том,
- 91. Рефлекторное последействие – продолжение рефлекторного акта после прекращения поступления афферентных сигналов. Причины: длительный ВПСП длительная следовая
- 92. Свойства нервных центров Тонус нервных центров. Для многих нейронных объединений, или нервных центров, характерна фоновая активность,
- 93. Нейроны-пейсмекеры Модулирующее влияние гуморальных факторов Афферентные сигналы от различных рефлексогенных зон Суммация миниатюрных (спонтанных) ВПСП Циркуляция
- 94. Свойства нервных центров Высокая утомляемость нервных центров. Это свойство характерно для нейронных цепей, в том числе
- 95. Высокая чувствительность к изменениям внутренней среды toC, О2, рН, глюкозы, токсинам. Пластичность (способность к перестройкам, синаптическое
- 96. Свойства нервных центров Пластичность нервных центров – это их способность к перестройке функциональных свойств и, в
- 97. Принцип общего конечного пути (активация одного эфферентного нейрона афферентами разных рефлексогенных зон). Принцип силы (приоритет более
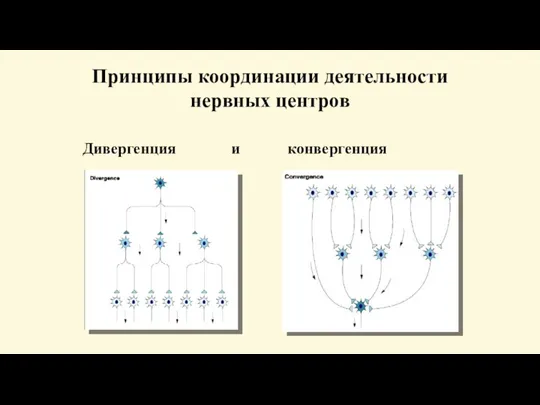
- 98. Принципы координации деятельности нервных центров Дивергенция и конвергенция
- 99. Интеграция синаптических влияний Дивергенция – способность нейрона устанавливать многочисленные синаптические связи с различными нервнымт клетками. Конвергенция
- 101. Скачать презентацию

Слайд 31. Основные понятия физиологии.
Принципы организации физиологических функций.
ФИЗИОЛОГИЯ
(от лат. phisis – природа, logos –
1. Основные понятия физиологии.
Принципы организации физиологических функций.
ФИЗИОЛОГИЯ
(от лат. phisis – природа, logos –

Слайд 4ОСНОВНЫЕ ПОНЯТИЯ ФИЗИОЛОГИИ
Физиологическая система – наследственно закрепленная совокупность органов и тканей, выполняющая
ОСНОВНЫЕ ПОНЯТИЯ ФИЗИОЛОГИИ
Физиологическая система – наследственно закрепленная совокупность органов и тканей, выполняющая

Слайд 5Учение о функциональных системах в живых организмах создано академиком
П. К. Анохиным (1935).
Функциональная
Учение о функциональных системах в живых организмах создано академиком
П. К. Анохиным (1935).
Функциональная

Слайд 6Достижение полезного приспособительного результата возможно на основе регуляции
Регуляция – это комплекс процессов,
Достижение полезного приспособительного результата возможно на основе регуляции
Регуляция – это комплекс процессов,

Слайд 7Конечный полезный приспособительный результат
– это поддержание некоего постоянства внутренней среды организма -
Конечный полезный приспособительный результат
– это поддержание некоего постоянства внутренней среды организма -

Слайд 8Нейрофизиология - специальный раздел физиологии, изучающий функции нервной системы и ее основных
Нейрофизиология - специальный раздел физиологии, изучающий функции нервной системы и ее основных

Слайд 9ПРЕДМЕТ НЕЙРОФИЗИОЛОГИИ
функционирование нервной системы и нейрона
ЗАДАЧИ
закономерности функционирования центральной нервной системы (ЦНС),
нейронная организация
ПРЕДМЕТ НЕЙРОФИЗИОЛОГИИ
функционирование нервной системы и нейрона
ЗАДАЧИ
закономерности функционирования центральной нервной системы (ЦНС),
нейронная организация

Слайд 10МЕТОДЫ
метод наблюдения;
метод экстирпации
метод поперечных перерезок
метод раздражения
стереотаксическая методика
метод условных рефлексов
нейрофизиологическое
МЕТОДЫ
метод наблюдения;
метод экстирпации
метод поперечных перерезок
метод раздражения
стереотаксическая методика
метод условных рефлексов
нейрофизиологическое

Слайд 11Физиологические свойства нервных клеток:
Раздражимость – способность живых клеток реагировать на воздействия внешней
Физиологические свойства нервных клеток:
Раздражимость – способность живых клеток реагировать на воздействия внешней

Слайд 12Раздражимость – это универсальное свойство всех биологических систем (от клетки до популяционного
Раздражимость – это универсальное свойство всех биологических систем (от клетки до популяционного

Слайд 13
Все ткани организма в зависимости от свойств клеточных мембран делятся на
Электровозбудимые;
Хемовозбудимые;
Механовозбудимые.
В
Все ткани организма в зависимости от свойств клеточных мембран делятся на
Электровозбудимые;
Хемовозбудимые;
Механовозбудимые.
В

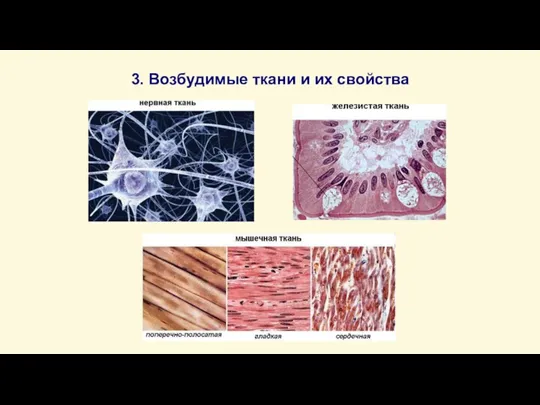
Слайд 143. Возбудимые ткани и их свойства
3. Возбудимые ткани и их свойства
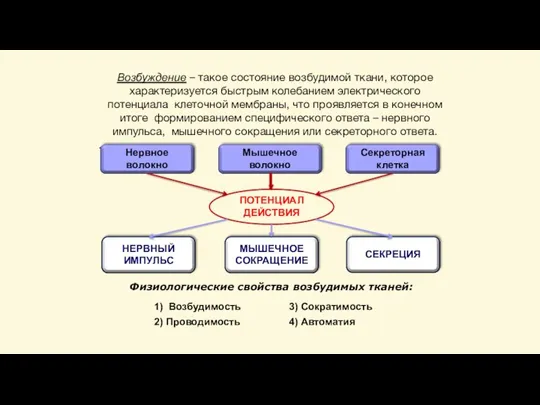
Слайд 15Возбуждение – такое состояние возбудимой ткани, которое характеризуется быстрым колебанием электрического потенциала
Возбуждение – такое состояние возбудимой ткани, которое характеризуется быстрым колебанием электрического потенциала
Слайд 16Раздражимость. Типы Раздражителей.
Любая живая ткань может находиться как в состоянии покоя, так
Раздражимость. Типы Раздражителей.
Любая живая ткань может находиться как в состоянии покоя, так

Слайд 17Раздражитель – любое изменение внешней среды или внутреннего состояния организма, воспринимаемое клетками
Раздражитель – любое изменение внешней среды или внутреннего состояния организма, воспринимаемое клетками

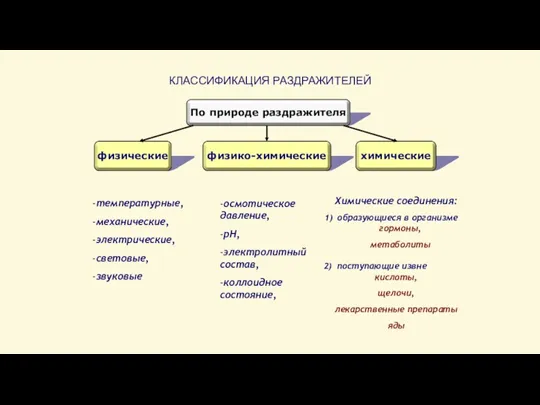
Слайд 18Химические соединения:
образующиеся в организме
гормоны, метаболиты
поступающие извне
кислоты, щелочи,
лекарственные препараты яды
КЛАССИФИКАЦИЯ РАЗДРАЖИТЕЛЕЙ
По природе раздражителя
физические
физико-химические
химические
-температурные,
-механические,
-электрические,
-световые,
-звуковые
-осмотическое
Химические соединения:
образующиеся в организме
гормоны, метаболиты
поступающие извне
кислоты, щелочи,
лекарственные препараты яды
КЛАССИФИКАЦИЯ РАЗДРАЖИТЕЛЕЙ
По природе раздражителя
физические
физико-химические
химические
-температурные,
-механические,
-электрические,
-световые,
-звуковые
-осмотическое
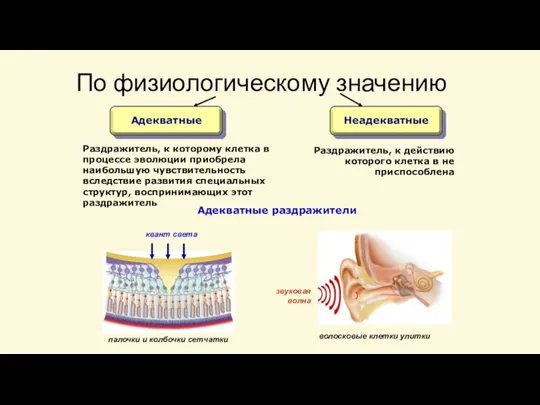
Слайд 19По физиологическому значению
Адекватные
Неадекватные
Раздражитель, к которому клетка в процессе эволюции приобрела наибольшую чувствительность
По физиологическому значению
Адекватные
Неадекватные
Раздражитель, к которому клетка в процессе эволюции приобрела наибольшую чувствительность
Слайд 20Схема строения клеточной мембраны:
молекула липида;
липидный бислой;
интегральные белки;
полуинтегральные белки;
Схема строения клеточной мембраны:
молекула липида;
липидный бислой;
интегральные белки;
полуинтегральные белки;

Слайд 21Функции мембран
Мембрана – граница между внутренней и внешней средами клетки, отличающимися по
Функции мембран
Мембрана – граница между внутренней и внешней средами клетки, отличающимися по

Слайд 22Пути проникновения веществ через мембрану клетки
Существуют 2 принципиальных механизма перемещения веществ через
Пути проникновения веществ через мембрану клетки
Существуют 2 принципиальных механизма перемещения веществ через
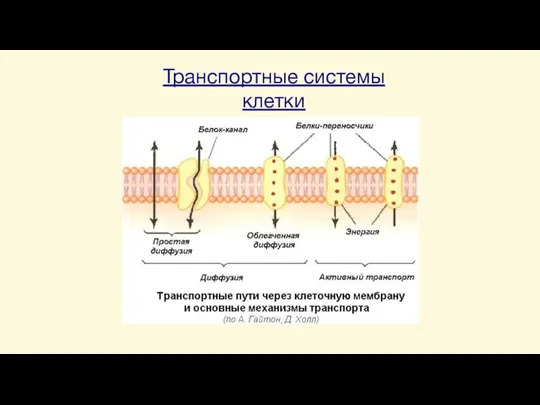
Слайд 23Транспортные системы клетки
Транспортные системы клетки
Слайд 24Пассивный транспорт – диффузия частиц через систему трансмембранных каналов.
Ионные каналы – это
Пассивный транспорт – диффузия частиц через систему трансмембранных каналов.
Ионные каналы – это
Слайд 25Активный транспорт – процесс перемещения молекул или ионов через клеточную мембрану против
Активный транспорт – процесс перемещения молекул или ионов через клеточную мембрану против
Слайд 26Основные виды электрических ответов нервных клеток:
Локальный ответ;
Потенциал действия;
Возбуждающий и тормозный постсинаптические потенциалы;
Генераторный
Основные виды электрических ответов нервных клеток:
Локальный ответ;
Потенциал действия;
Возбуждающий и тормозный постсинаптические потенциалы;
Генераторный

Слайд 27Мембранно-ионная теория. Природа потенциала покоя
1902 г. - И. Бернштейн сформулировал основные положения
Мембранно-ионная теория. Природа потенциала покоя
1902 г. - И. Бернштейн сформулировал основные положения

Слайд 28Потенциал покоя
ПП - это разность электрических потенциалов между внутренней и наружной
Потенциал покоя
ПП - это разность электрических потенциалов между внутренней и наружной

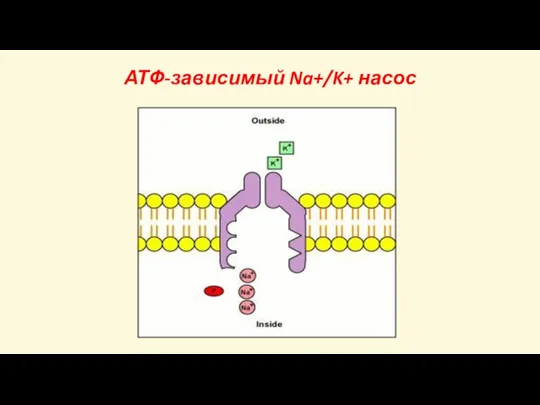
Слайд 29Потенциал покоя создается
Во-первых, за счет работы АТФ-зависимого Na+/K+ насоса (асимметричного обмена Na+
Потенциал покоя создается
Во-первых, за счет работы АТФ-зависимого Na+/K+ насоса (асимметричного обмена Na+

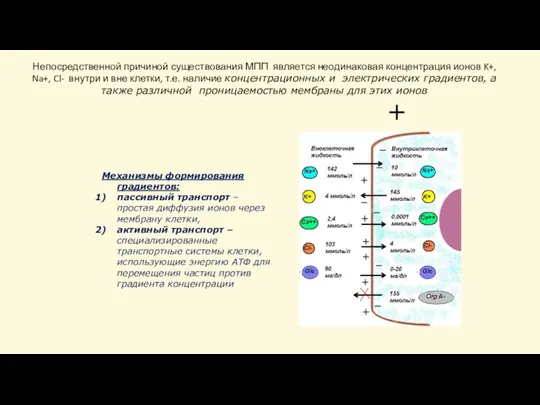
Слайд 30Механизмы формирования градиентов:
пассивный транспорт – простая диффузия ионов через мембрану клетки,
активный транспорт
Механизмы формирования градиентов:
пассивный транспорт – простая диффузия ионов через мембрану клетки,
активный транспорт
Слайд 31АТФ-зависимый Na+/K+ насос
АТФ-зависимый Na+/K+ насос
Слайд 32Основные положения
мембранно-ионной теории возбуждения
1.
В покое мембрана клетки находится в состоянии
статической поляризации,
Основные положения
мембранно-ионной теории возбуждения
1.
В покое мембрана клетки находится в состоянии
статической поляризации,

Слайд 33Порог возбудимости аксонного холмика - 10 мВ, тела клетки - 50 мВ;
Порог возбудимости аксонного холмика - 10 мВ, тела клетки - 50 мВ;

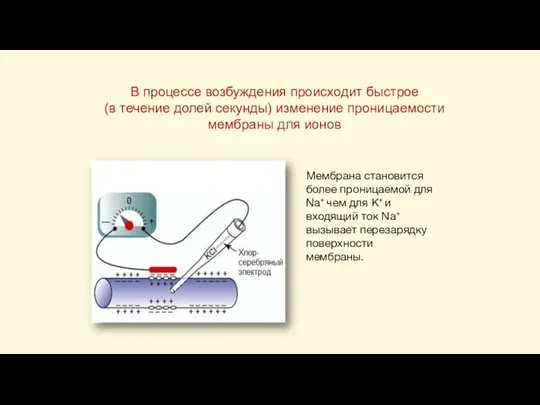
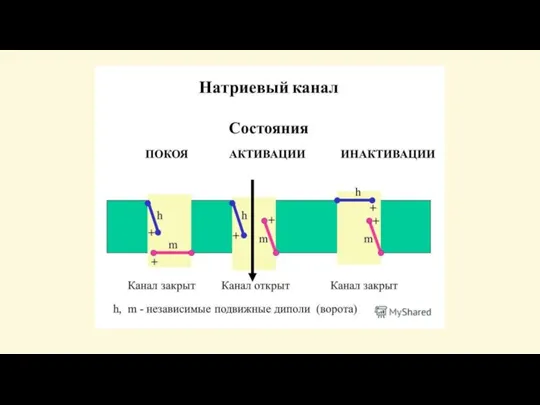
Слайд 34В процессе возбуждения происходит быстрое
(в течение долей секунды) изменение проницаемости мембраны для
В процессе возбуждения происходит быстрое
(в течение долей секунды) изменение проницаемости мембраны для
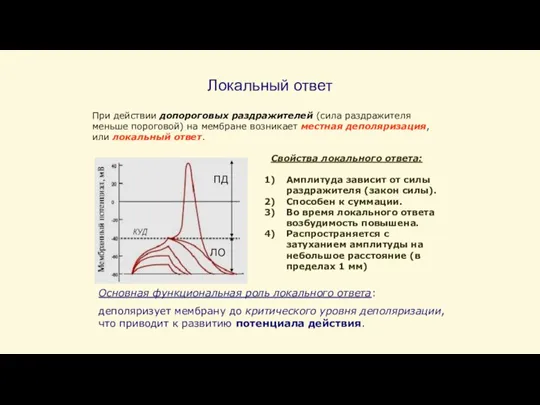
Слайд 36Локальный ответ
ПД
ЛО
При действии допороговых раздражителей (сила раздражителя меньше пороговой) на мембране возникает
Локальный ответ
ПД
ЛО
При действии допороговых раздражителей (сила раздражителя меньше пороговой) на мембране возникает
Слайд 37Потенциал действия
ПД - быстрое колебание мембранного потенциала, возникающее при возбуждении нервных
Потенциал действия
ПД - быстрое колебание мембранного потенциала, возникающее при возбуждении нервных

Слайд 38Потенциал действия
Потенциал действия (ПД) – это электрофизиологический процесс, выражающийся в быстром колебании
Потенциал действия
Потенциал действия (ПД) – это электрофизиологический процесс, выражающийся в быстром колебании

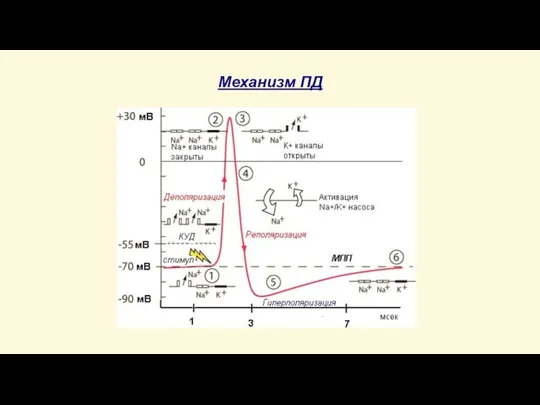
Слайд 39Механизм ПД
ВОСХОДЯЩАЯ ФАЗА ПД:
раздражитель пороговой или сверхпороговой силы увеличивает проницаемость мембраны для
Механизм ПД
ВОСХОДЯЩАЯ ФАЗА ПД:
раздражитель пороговой или сверхпороговой силы увеличивает проницаемость мембраны для

Слайд 40Механизм ПД
Механизм ПД
Слайд 41Свойства потенциала действия
Возникает в ответ на действие пороговых и сверхпороговых раздражителей (подчиняется
Свойства потенциала действия
Возникает в ответ на действие пороговых и сверхпороговых раздражителей (подчиняется

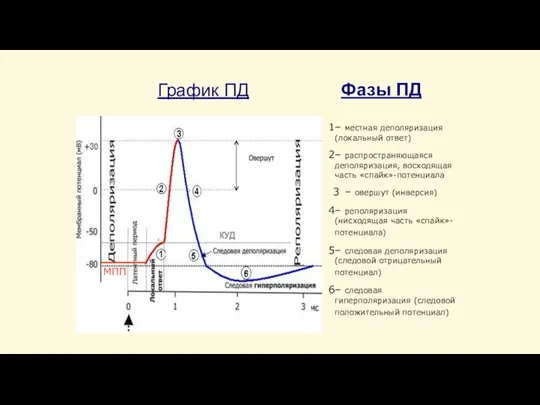
Слайд 42Фазы потенциала действия
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное
Фазы потенциала действия
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное

Слайд 43График ПД
– местная деполяризация (локальный ответ)
– распространяющаяся деполяризация, восходящая часть «спайк»-потенциала
– овершут
График ПД
– местная деполяризация (локальный ответ)
– распространяющаяся деполяризация, восходящая часть «спайк»-потенциала
– овершут
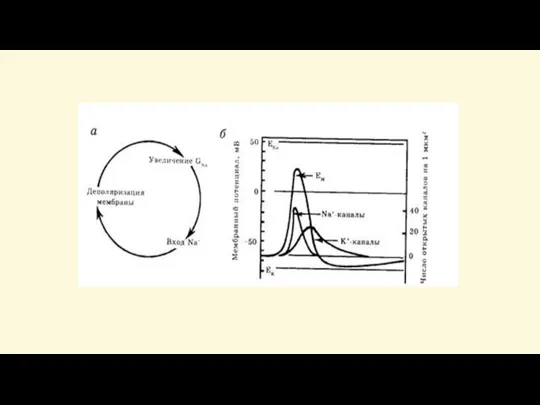
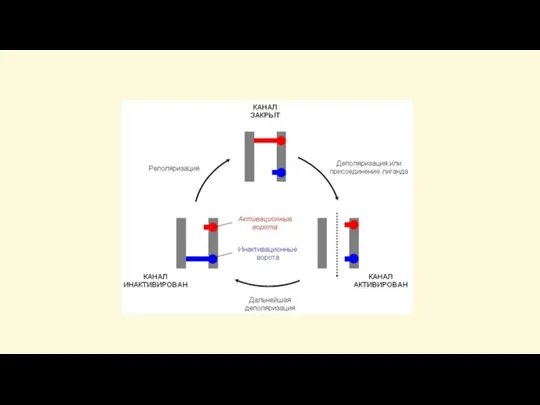
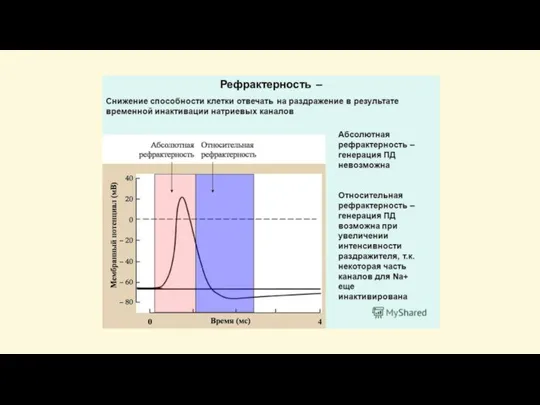
Слайд 48Закон «всё-или-ничего»
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на
Закон «всё-или-ничего»
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на

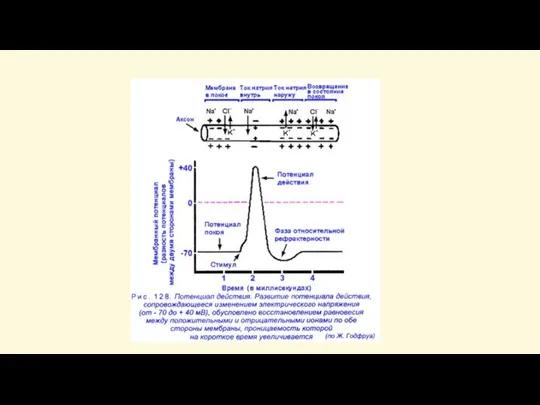
Слайд 49Проводимость нейрона
Скорость проведения по нейрону зависит от строения нервного волокна (наличия, отсутствия
Проводимость нейрона
Скорость проведения по нейрону зависит от строения нервного волокна (наличия, отсутствия

Слайд 50Распространение потенциала действия по
немиелинизированным волокнам
По немиелинизированному волокну ПД распространяется
Распространение потенциала действия по
немиелинизированным волокнам
По немиелинизированному волокну ПД распространяется

Слайд 51Распространение потенциала действия по немиелинизированным волокнам
Распространение потенциала действия по немиелинизированным волокнам

Слайд 52Распространение потенциала действия по миелинизированным волокнам
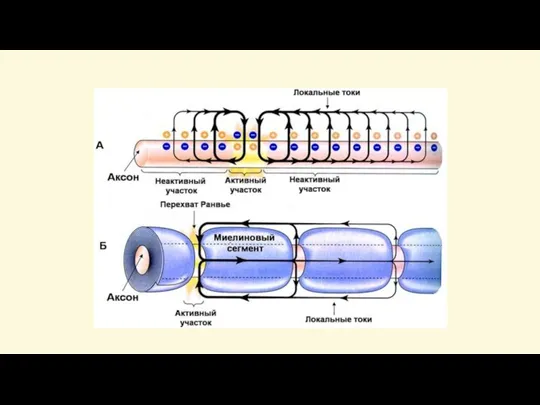
В миелиновых волокнах возбуждение охватывает только
Распространение потенциала действия по миелинизированным волокнам
В миелиновых волокнах возбуждение охватывает только
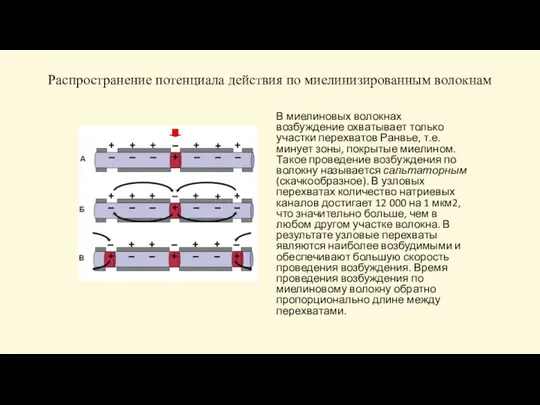
Слайд 54 Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта

Слайд 55Электрический синапс
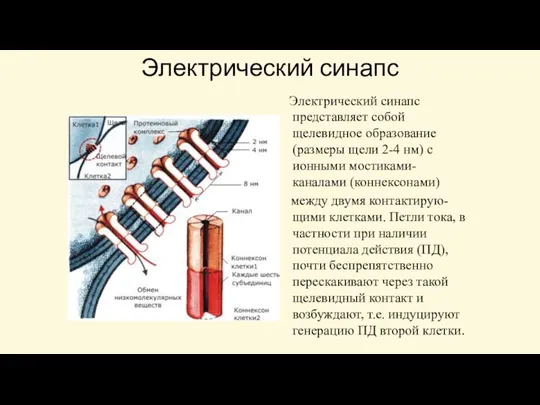
Электрический синапс представляет собой щелевидное образование (размеры щели 2-4 нм)
Электрический синапс
Электрический синапс представляет собой щелевидное образование (размеры щели 2-4 нм)
Слайд 56Работа электрического синапса
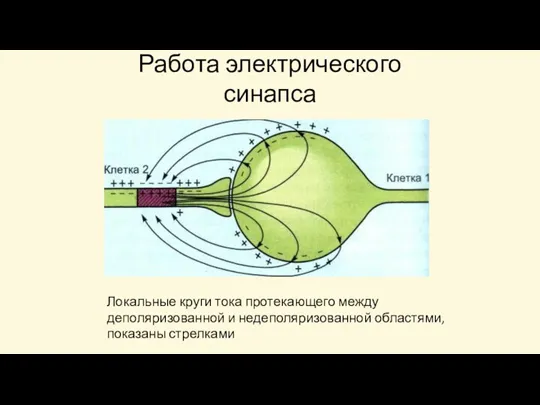
Локальные круги тока протекающего между деполяризованной и недеполяризованной областями, показаны
Работа электрического синапса
Локальные круги тока протекающего между деполяризованной и недеполяризованной областями, показаны
Слайд 57Строение химического синапса
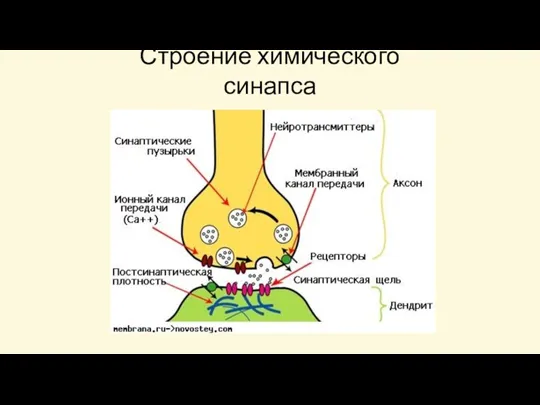
Строение химического синапса
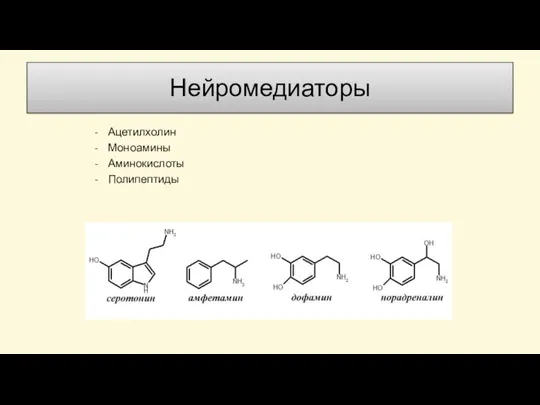
Слайд 58Нейромедиаторы
Ацетилхолин
Моноамины
Аминокислоты
Полипептиды
Нейромедиаторы
Ацетилхолин
Моноамины
Аминокислоты
Полипептиды
Слайд 59Схема экзоцитоза.
Схема экзоцитоза.

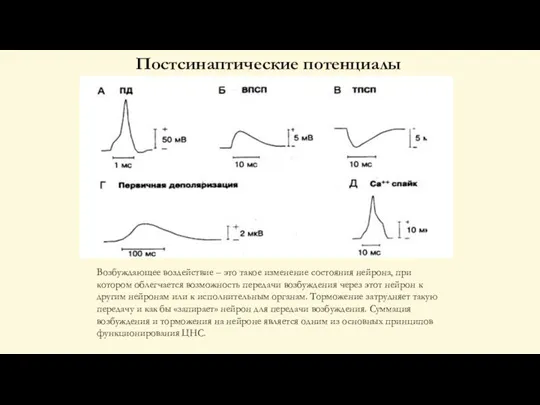
Слайд 60Постсинаптические потенциалы
Возбуждающее воздействие – это такое изменение состояния нейрона, при котором облегчается
Постсинаптические потенциалы
Возбуждающее воздействие – это такое изменение состояния нейрона, при котором облегчается
Слайд 61Торможение в ЦНС – это активный нервный процесс, результатом которого является прекращение
Торможение в ЦНС – это активный нервный процесс, результатом которого является прекращение

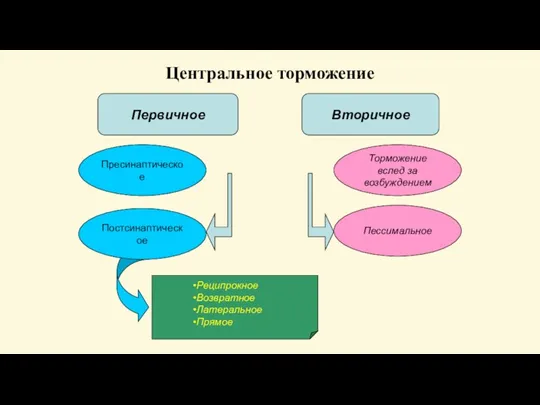
Слайд 62Центральное торможение
Торможение вслед за
возбуждением
Пессимальное
Пресинаптическое
Реципрокное
Возвратное
Латеральное
Прямое
Первичное
Вторичное
Постсинаптическое
Центральное торможение
Торможение вслед за
возбуждением
Пессимальное
Пресинаптическое
Реципрокное
Возвратное
Латеральное
Прямое
Первичное
Вторичное
Постсинаптическое
Слайд 63
Торможение.
Торможение – самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого
Торможение.
Торможение – самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого

Слайд 64Пресинаптическое торможение
Развивается в пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными
Пресинаптическое торможение
Развивается в пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными

Слайд 65Постсинаптическое торможение
Медиатор, выделяемый постсинаптическими окончаниями тормозных синапсов, изменяет свойства постсинаптической мембраны таким
Постсинаптическое торможение
Медиатор, выделяемый постсинаптическими окончаниями тормозных синапсов, изменяет свойства постсинаптической мембраны таким

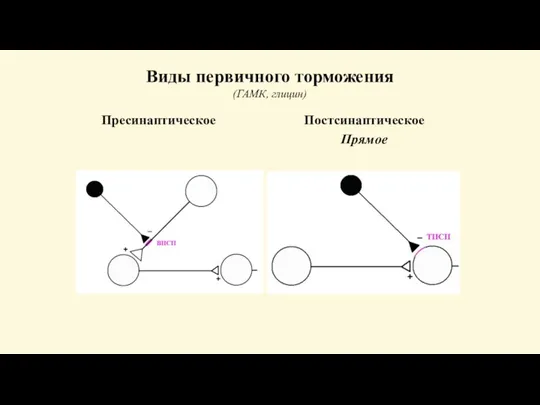
Слайд 66Виды первичного торможения
(ГАМК, глицин)
Пресинаптическое
Постсинаптическое
Прямое
Виды первичного торможения
(ГАМК, глицин)
Пресинаптическое
Постсинаптическое
Прямое
Слайд 67Торможение в ЦНС
В интегративной деятельности центральной нервной системы важная роль принадлежит тормозным
Торможение в ЦНС
В интегративной деятельности центральной нервной системы важная роль принадлежит тормозным

Слайд 68Возвратное торможение
Если тормозные вставочные нейроны действуют на те же нервные клетки,
Возвратное торможение
Если тормозные вставочные нейроны действуют на те же нервные клетки,

Слайд 69Реципрокное торможение
Этот вид торможения широко представлен в спинном мозге. С
Реципрокное торможение
Этот вид торможения широко представлен в спинном мозге. С

Слайд 70Латеральное торможение
Данный вид торможения широко представлен в структурах нервной системы,
Латеральное торможение
Данный вид торможения широко представлен в структурах нервной системы,

Слайд 71Виды торможения
Виды торможения

Слайд 72Нервный центр – это функциональное объединение нейронов, принимающих участие в осуществлении одной
Нервный центр – это функциональное объединение нейронов, принимающих участие в осуществлении одной
Слайд 73Нервный центр
Нервным центром называется совокупность нейронов в ЦНС, обеспечивающих регуляцию какой-либо функции
Нервный центр
Нервным центром называется совокупность нейронов в ЦНС, обеспечивающих регуляцию какой-либо функции

Слайд 74Нейроны нервного центра могут располагаться на разных уровнях ЦНС.
Нейроны нервного центра могут располагаться на разных уровнях ЦНС.
Слайд 75Обусловлены:
Свойствами входящих в его состав нейронов
Свойствами синаптических образований
Особенностями
Обусловлены:
Свойствами входящих в его состав нейронов
Свойствами синаптических образований
Особенностями

Слайд 76
Свойства нервных центров
Одностороннее проведение возбуждения - возбуждение передается с афферентного на эфферентный
Свойства нервных центров
Одностороннее проведение возбуждения - возбуждение передается с афферентного на эфферентный

Слайд 77
Имеется 2 вида суммации: временная и пространственная.
Временная - возникает ответная реакция
Имеется 2 вида суммации: временная и пространственная.
Временная - возникает ответная реакция

Слайд 78Суммация
а). Пространственная (следствие интегративной функции нейрона)
Суммация
а). Пространственная (следствие интегративной функции нейрона)
Слайд 79Суммация
б). Временная
Суммация
б). Временная
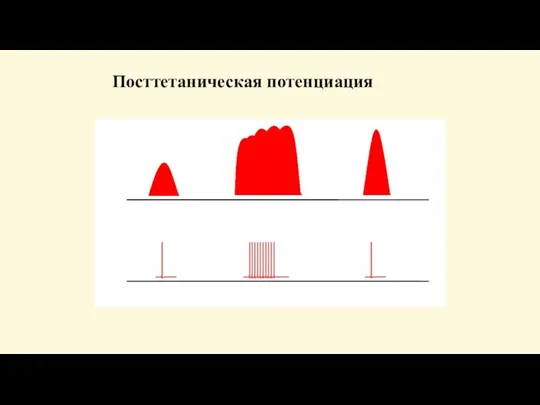
Слайд 80Посттетаническая потенция
- усиление ответной реакции, наблюдается после серии нервных импульсов. Механизм: потенциация
Посттетаническая потенция
- усиление ответной реакции, наблюдается после серии нервных импульсов. Механизм: потенциация

Слайд 81Посттетаническая потенциация
Посттетаническая потенциация
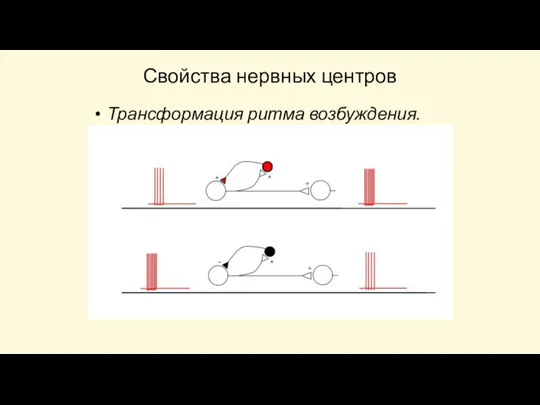
Слайд 82Трансформация ритма возбуждения
- несоответствие ответной реакции частоте наносимых раздражений. На афферентном нейроне
Трансформация ритма возбуждения
- несоответствие ответной реакции частоте наносимых раздражений. На афферентном нейроне

Слайд 83Свойства нервных центров
Трансформация ритма возбуждения.
Свойства нервных центров
Трансформация ритма возбуждения.
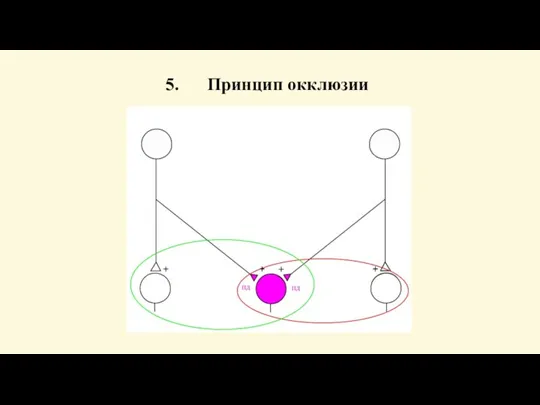
Слайд 84Свойства нервных центров
Явление окклюзии отражает эффект взаимодействия между собой двух импульсных потоков,
Свойства нервных центров
Явление окклюзии отражает эффект взаимодействия между собой двух импульсных потоков,

Слайд 85Принцип окклюзии
Принцип окклюзии
Слайд 86Свойства нервных центров
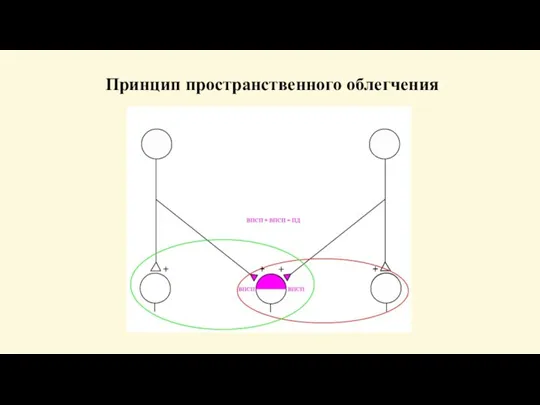
Явление облегчения, которое по своему внешнему проявлению противоположно окклюзии.
0но
Свойства нервных центров
Явление облегчения, которое по своему внешнему проявлению противоположно окклюзии.
0но

Слайд 87Центральное облегчение
Объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный
Центральное облегчение
Объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный

Слайд 88Принцип пространственного облегчения
Принцип пространственного облегчения
Слайд 89Рефлекторное последействие
- продолжение ответной реакции после прекращения действия раздражителя:
кратковременное последействие - в
Рефлекторное последействие
- продолжение ответной реакции после прекращения действия раздражителя:
кратковременное последействие - в

Слайд 90Свойства нервных центров
Последействие. Это одно из свойств, характерных для нейронных цепей. Оно
Свойства нервных центров
Последействие. Это одно из свойств, характерных для нейронных цепей. Оно

Слайд 91Рефлекторное последействие – продолжение рефлекторного акта после прекращения поступления афферентных сигналов.
Причины:
длительный ВПСП
длительная
Рефлекторное последействие – продолжение рефлекторного акта после прекращения поступления афферентных сигналов.
Причины:
длительный ВПСП
длительная
Слайд 92Свойства нервных центров
Тонус нервных центров. Для многих нейронных объединений, или нервных центров,
Свойства нервных центров
Тонус нервных центров. Для многих нейронных объединений, или нервных центров,

Слайд 93Нейроны-пейсмекеры
Модулирующее влияние гуморальных факторов
Афферентные сигналы от различных рефлексогенных зон
Суммация миниатюрных (спонтанных)
Нейроны-пейсмекеры
Модулирующее влияние гуморальных факторов
Афферентные сигналы от различных рефлексогенных зон
Суммация миниатюрных (спонтанных)

Слайд 94Свойства нервных центров
Высокая утомляемость нервных центров. Это свойство характерно для нейронных цепей,
Свойства нервных центров
Высокая утомляемость нервных центров. Это свойство характерно для нейронных цепей,

Слайд 95Высокая чувствительность к изменениям внутренней среды
toC,
О2,
рН,
глюкозы,
токсинам.
Пластичность
Высокая чувствительность к изменениям внутренней среды
toC,
О2,
рН,
глюкозы,
токсинам.
Пластичность

Слайд 96Свойства нервных центров
Пластичность нервных центров – это их способность к перестройке функциональных
Свойства нервных центров
Пластичность нервных центров – это их способность к перестройке функциональных

Слайд 97Принцип общего конечного пути
(активация одного эфферентного нейрона афферентами разных рефлексогенных зон).
Принцип
Принцип общего конечного пути
(активация одного эфферентного нейрона афферентами разных рефлексогенных зон).
Принцип

Слайд 98Принципы координации деятельности нервных центров
Дивергенция и конвергенция
Принципы координации деятельности нервных центров
Дивергенция и конвергенция
Слайд 99Интеграция синаптических влияний
Дивергенция – способность нейрона устанавливать многочисленные синаптические связи с различными
Интеграция синаптических влияний
Дивергенция – способность нейрона устанавливать многочисленные синаптические связи с различными

 Биологические системы
Биологические системы Краниальные нервы и их остеопатическое повреждение
Краниальные нервы и их остеопатическое повреждение Микроэволюция. Популяция. Выявление изменчивости у особей одного вида
Микроэволюция. Популяция. Выявление изменчивости у особей одного вида Дарыу үләндәре
Дарыу үләндәре Презентация на тему Влияние солнечной активности на социальные процессы
Презентация на тему Влияние солнечной активности на социальные процессы  Размножение растений вегетативным путям
Размножение растений вегетативным путям Сцепленное наследование признаков
Сцепленное наследование признаков Биологические (естественные) потребности
Биологические (естественные) потребности Презентация на тему КЛЕТКИ И ТКАНИ ЧЕЛОВЕКА
Презентация на тему КЛЕТКИ И ТКАНИ ЧЕЛОВЕКА  Быстрый сон
Быстрый сон Строение и работа сердца
Строение и работа сердца Координация и регуляция процессов жизнедеятельности живых организмов. Биология 6 класс
Координация и регуляция процессов жизнедеятельности живых организмов. Биология 6 класс Красная книга. Тамбовская область
Красная книга. Тамбовская область Спинной мозг Форма, топография, основные отделы спинного мозга
Спинной мозг Форма, топография, основные отделы спинного мозга Строение и биологическая роль витаминов
Строение и биологическая роль витаминов Черенкование растения
Черенкование растения Анатомия животных
Анатомия животных Актуальные и перспективные медицинские технологии
Актуальные и перспективные медицинские технологии лекция 6,5
лекция 6,5 Развитие животного мира на Земле
Развитие животного мира на Земле 12. Обеспечение клеток энергией
12. Обеспечение клеток энергией Віруси, віроїди, пріони
Віруси, віроїди, пріони Процесс фотосинтеза
Процесс фотосинтеза Животные. Викторина
Животные. Викторина Формы естественного отбора. Задание
Формы естественного отбора. Задание At the fruit farm
At the fruit farm Папоротникообразные
Папоротникообразные Cирийский хомяк
Cирийский хомяк