- Teoreticheskiy_minimum

Содержание
- 2. Введение Живая природа подчиняется иерархической организации. Иерархический принцип организации позволяет выделить в живой природе отдельные уровни,
- 3. Микроскоп - (от греческого mikros - малый и skopeo - смотрю), оптический прибор для получения увеличенного
- 5. Световая микроскопия. Устройство светового микроскопа Изучение препарата осуществляется в проходящем свете с помощью светового микроскопа. Источник
- 6. Основными частями светового микроскопа являются объектив и окуляр, заключенные в цилиндрический корпус – тубус. Большинство моделей,
- 7. Основание микроскопа Тубусодержатель Тубус Окуляр (чаще ×10) Револьвер микроскопа Объективы а) сухие: ×10, ×20, ×40 б)
- 8. Характеристики оптического микроскопа К основным характеристикам микроскопа относятся увеличение и разрешающая способность. Общее увеличение, которое дает
- 9. Электронная микроскопия. Устройство электронного микроскопа Электронный микроскоп — прибор, позволяющий получать изображение объектов с максимальным увеличением
- 10. Электронный микроскоп состоит из электронной пушки (устройства для получения пучка электронов) и системы электромагнитных линз, размещенных
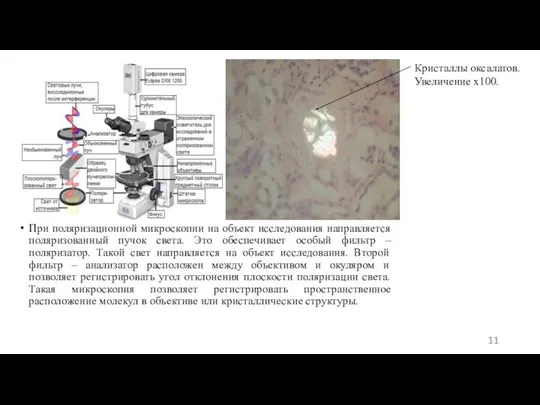
- 11. При поляризационной микроскопии на объект исследования направляется поляризованный пучок света. Это обеспечивает особый фильтр – поляризатор.
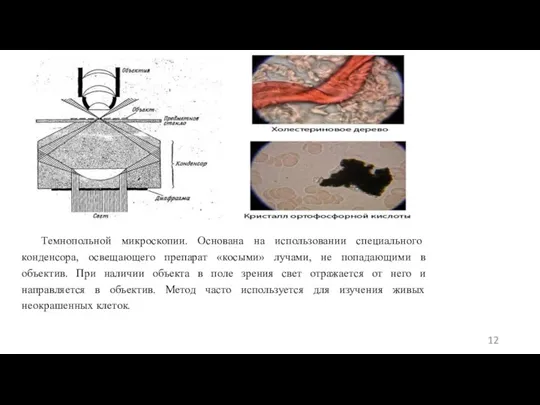
- 12. Темнопольной микроскопии. Основана на использовании специального конденсора, освещающего препарат «косыми» лучами, не попадающими в объектив. При
- 13. Метод фазово-контрастной микроскопии служит для получения контрастных изображений прозрачных и бесцветных объектов. Даже при очень малых
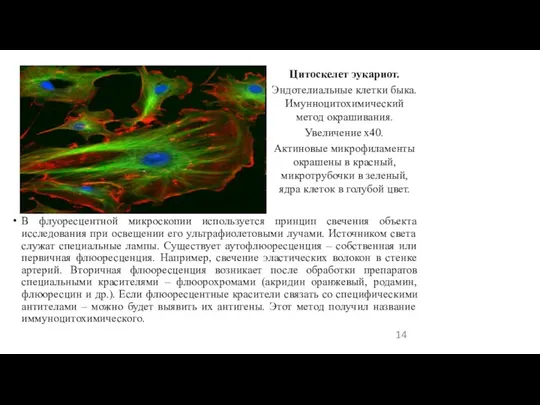
- 14. В флуоресцентной микроскопии используется принцип свечения объекта исследования при освещении его ультрафиолетовыми лучами. Источником света служат
- 15. Правила работы с микроскопом Установите микроскоп слева, штативом к себе; Поставьте в рабочее положение объектив малого
- 16. Поверхностный аппарат клетки
- 17. Определение клетки, ее компоненты Клетка - целостная элементарная система, способная к самовоспроизведению и саморегуляции метаболических процессов.
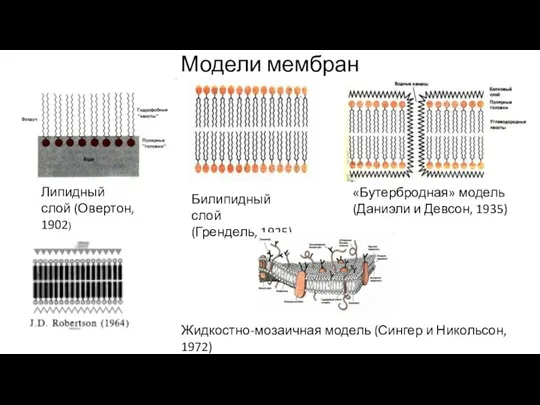
- 18. Модели мембран Липидный слой (Овертон, 1902) Билипидный слой (Грендель, 1925) «Бутербродная» модель (Даниэли и Девсон, 1935)
- 19. Поверхностный аппарат клетки
- 20. Плазматическая мембрана образована белками (-60%) и липидами (-40%). В состав липидов входят гликолипиды, стеролы, но наиболее
- 21. Свойства плазматической мембраны Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою структуру, т.к. молекулы
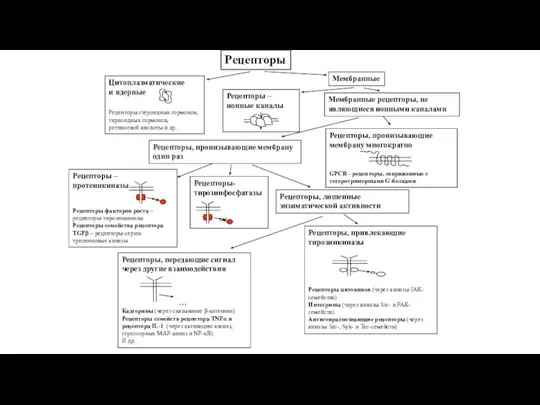
- 23. Белки Полуинтегральные (белки адгезии, рецепторы); Интегральные (поры, ионные каналы, переносчики, насосы, рецепторы ) Периферические (рецепторы, белки
- 25. Функциональная классификация мембранных белков
- 27. Липиды мембраны Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. Фосфолипиды в составе мембран
- 28. Фосфолипиды Фосфолипиды являются неотъемлемым компонентом всех клеточных мембран . Между плазмой и эритроцитами постоянно происходит обмен
- 29. Гликолипиды Гликолипиды составляют 5% липидных молекул наружного монослоя и сильно различаются у разных видов и даже
- 30. Холестерин Холестерин - это стерин, содержащий стероидное ядро из четырех колец и гидроксильную группу. Холестерин содержится
- 32. ВЛИЯНИЕ ХИМИЧЕСКОГО СОСТАВА Зависимость свойств мембраны от химического состава называется лиотропным полиморфизмом. Ненасыщенных жирные кислоты способствуют
- 33. ТРАНСПОРТ ВЕЩЕСТВ В МЕМБРАНАХ
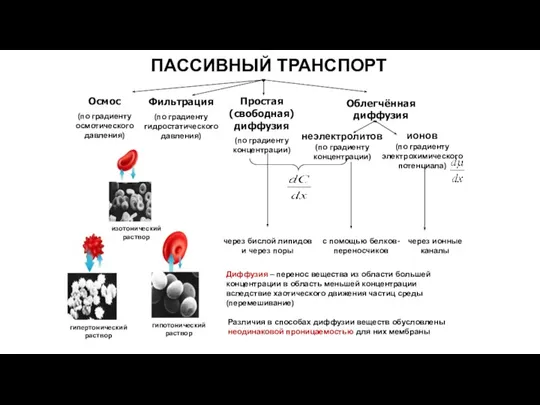
- 34. Виды транспорта
- 36. Осмос и электроосмос Это движение растворителя из раствора с меньшей концентрацией в раствор с большей концентрацией.
- 37. Фильтрация Это движение растворителя под действием гидростатического давления
- 38. ПРОНИЦАЕМОСТЬ МЕМБРАНЫ Свободнаядиффузия Облегчённая диффузия
- 39. Простая диффузия Линейно зависит от градиента концентрации вещества; Характеризуется ненасыщаемостью.
- 40. Простая диффузия осуществляется через: Мембрану (для незаряженных жирорастворимых) веществ; Поры; Кинки.
- 41. Простая диффузия идет через кинки При повышении температуры тепловая подвижность жирнокислотных цепей приводит к спонтанному возникновению
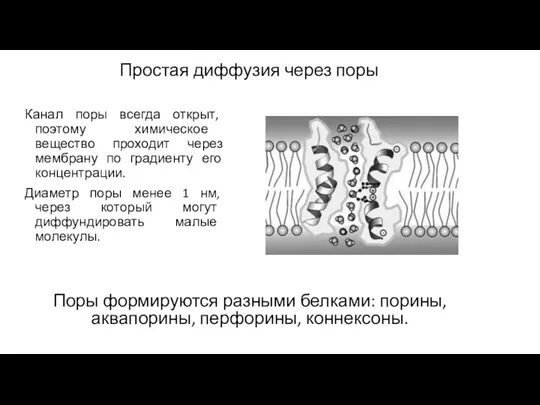
- 42. Простая диффузия через поры Канал поры всегда открыт, поэтому химическое вещество проходит через мембрану по градиенту
- 43. Переносчики (транспортёры) специфичны: каждый конкретный переносчик переносит через липидный бислой, как правило одно вещество.
- 44. ЭЛЕКТРОДИФФУЗИЯ ;
- 45. Ионные каналы Cостоят из связанных между собой белковых субъединиц, формирующих в мембране гидрофильную селективную пору
- 46. Свойства ионных каналов Специфичность Проводимость Наличие сенсора Наличие воротной системы
- 47. Четыре вида каналов: А - ионселективный канал (открытый) В – хемо-чувствительный канал С – потенциал-зависимый канал
- 48. ИОННЫЕ НАСОСЫ переносят вещества против их градиентов за счёт энергии гидролиза АТФ ПЕРЕНОСЧИКИ транспортируют вещества против
- 49. Активный транспорт Осуществляется против электрохимического градиента Система в высшей степени специфична Необходимы источники энергии в виде
- 50. Первичный активный транспорт (насосы, АТФ-азы) Обеспечивает перенос веществ против градиента их концентрации с затратой энергии АТФ
- 51. Натрий-калий- магний - зависимая АТФаза Натрий-калий насос- интегральный белок мембраны, способный расщеплять АТФ. Можно ингибировать убаином.
- 52. Вторичный активный транспорт В качестве источника энергии использует химический или электрохимический градиент какого-либо вещества
- 53. Различают: однонаправленный (унипорт), сочетанный (симпорт) и разнонаправленный (антипорт) транспорт.
- 54. Эндоцитоз Образуется окаймленная везикула (3), Везикула сливается с эндосомой(4). Вакуоль и ее содержимое претерпевают превращения (5),
- 55. Экзоцитоз В аппарате Гольджи из предшественника образуется конечный секрет; Везикула с секретом доставляется к плазматической мембране;
- 56. Рецепторная функция. Постоянные межклеточные контакты. Цитоскелет
- 57. Этапы проведения сигнала 1) Прием сигнала (рецепция) 2) Обработка и передача сигнала (системы сигнальных путей /
- 58. Стимулы (информация, приходящая к клетке) 1) Высокоаффинные лиганды (связываются с рецептором высокоаффинно и высокоселективно) 2) Малоспецифичные
- 59. Ответы клетки Влияние на пролиферацию (стимуляция деления клеток / блок клеточного цикла); Влияние на апоптоз (стимуляция
- 60. Типы секреции (классификация лигандов по дальности действия) 1) Эндокринная секреция (в кровоток; все гормоны, многие цитокины,
- 61. Типы секреции (классификация лигандов по дальности действия) 2) Паракринная секреция (лиганды локального действия; простогландины, лейкотриены, многие
- 62. Типы секреции (классификация лигандов по дальности действия) 3) Аутокринная секреция (действие на ту же клетку или
- 63. Типы секреции (классификация лигандов по дальности действия) 4) Юкстакринная, или зависимая от контактов секреция (jucstacrine; лиганды
- 64. Типы секреции (классификация лигандов по дальности действия) 5) Секреция компонентов внеклеточного матрикса (нерастворимые лиганды рецептируются контактами
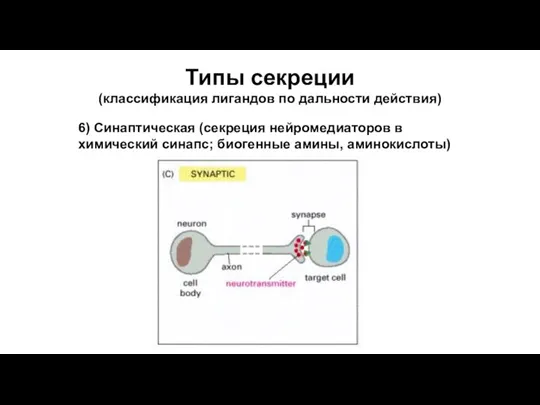
- 65. Типы секреции (классификация лигандов по дальности действия) 6) Синаптическая (секреция нейромедиаторов в химический синапс; биогенные амины,
- 66. Типы секреции (классификация лигандов по дальности действия) 7) Секреция через щелевые контакты (gap junctions; обмен внутриклеточными
- 67. Типы секреции (классификация лигандов по дальности действия) 6) Синаптическая (секреция нейромедиаторов в химический синапс; биогенные амины,
- 68. Клетки могут взаимодействовать друг с другом по средствам: На расстоянии через секрецию определенных веществ; Через сигнальные
- 69. Рецепторы
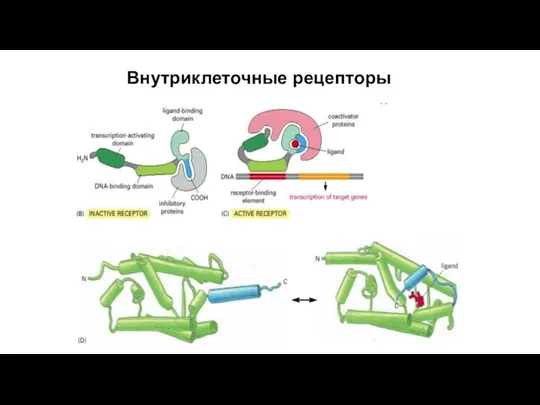
- 70. Внутриклеточные рецепторы
- 71. Цикл работы рецептора связанного с G-белком Субъединица альфа умеет расщеплять ГТФ. В активированном состоянии взаимодействует с
- 72. Мембранно-внутриклеточный механизм Вторичные посредники цАМФ - циклический аденозинмонофосфат; цГМФ - циклический гуанозинмонофосфат; ИФ3 - инозитол-3-фосфат; ДАГ
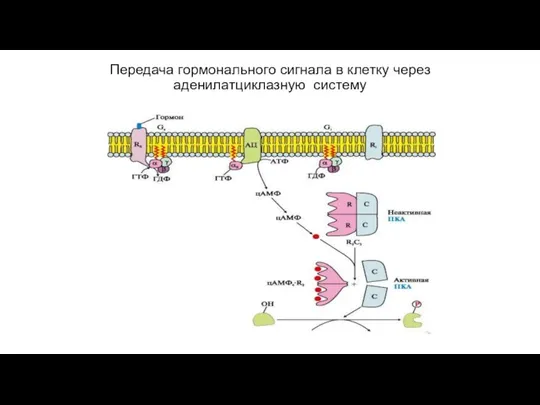
- 73. Вариант 1. (с участием аденилатциклазной системы) Вторичный посредник - цАМФ; Мембранно-связанный фермент - аденилатциклаза Гормон связывается
- 74. Передача гормонального сигнала в клетку через аденилатциклазную систему
- 75. Прекращение влияния цАМФ на процесс активации протеинкиназы А и, следовательно, фосфорилирования белков происходит под действием фосфодиэстеразы
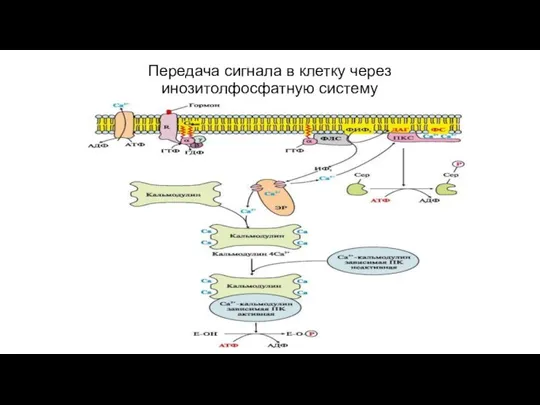
- 76. Вариант 2. (с участием инозитолфосфатной системы) Вторичные посредники- инозитол-3-фосфат (ИФ3), ДАГ Образуются путем гидролиза фосфолипида клеточной
- 77. Основные этапы передачи гормонального сигнала 1.Связывание гормона с рецептором меняется конформация рецептора. 2.Изменение конформации соответствующего G-белка,
- 78. Передача сигнала в клетку через инозитолфосфатную систему
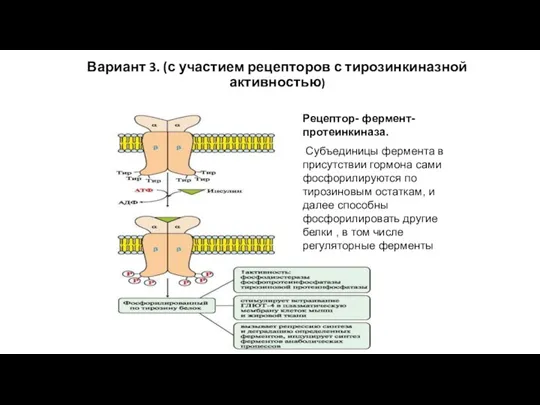
- 79. Вариант 3. (с участием рецепторов с тирозинкиназной активностью) Рецептор- фермент- протеинкиназа. Субъединицы фермента в присутствии гормона
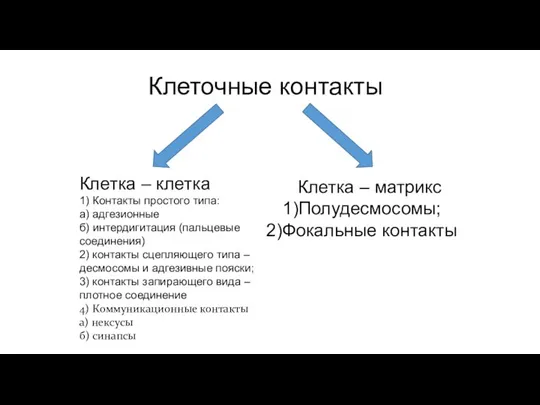
- 80. Клеточные контакты Клетка – клетка 1) Контакты простого типа: а) адгезионные б) интердигитация (пальцевые соединения) 2)
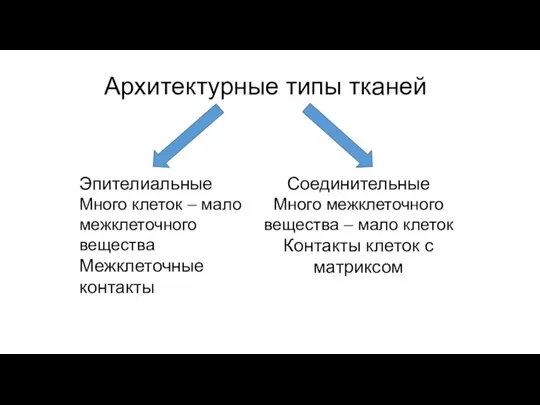
- 81. Архитектурные типы тканей Эпителиальные Много клеток – мало межклеточного вещества Межклеточные контакты Соединительные Много межклеточного вещества
- 82. Общая схема строения клеточных контактов Межклеточные контакты образуются по следующей схеме: Элемент цитоскелета (актиновые- или промежуточные
- 83. Контакты простого типа Адгезионные соединения Это простое сближение плазмолемм соседних клеток на расстояние 15-20 нм без
- 84. Интердигитация (пальцевидное соединение) (№ 2 на рисунке) представляет собой контакт, при котором плазмолемма двух клеток, сопровождая
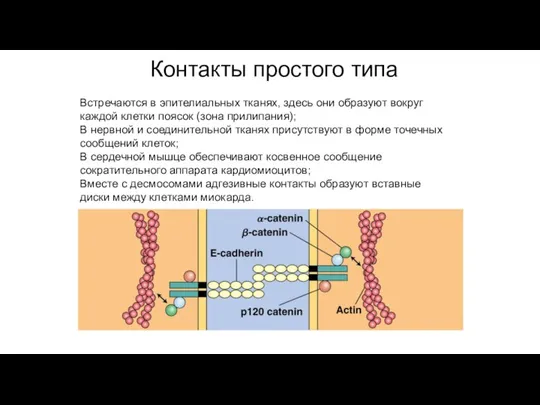
- 85. Контакты простого типа Встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки поясок (зона прилипания);
- 86. Контакты сцепляющего типа Десмосомы Полудесмосомы Поясок сцепления
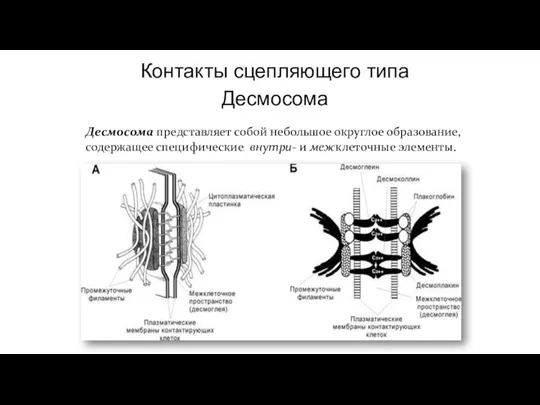
- 87. Контакты сцепляющего типа Десмосома Десмосома представляет собой небольшое округлое образование, содержащее специфические внутри- и межклеточные элементы.
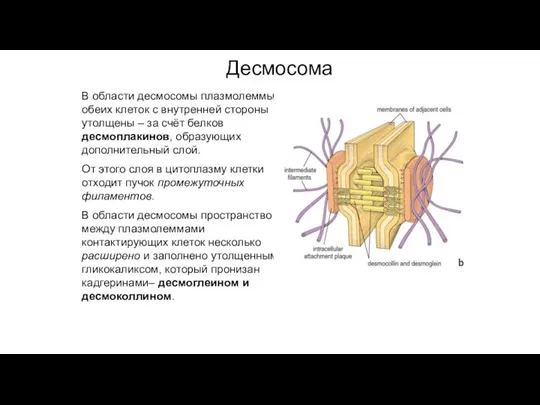
- 88. В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены – за счёт белков десмоплакинов, образующих
- 89. Полудесмосома Полудесмосома обеспечивает контакт клеток с базальной мембраной. По структуре гемидесмосомы напоминают десмосомы и тоже содержат
- 90. Полудесмосома
- 91. Поясок сцепления Адгезивный поясок, (поясок сцепления, поясная десмосома) (zonula adherens), – парное образование в виде лент,
- 92. Белки поясков сцепления 1. Утолщение плазмолеммы со стороны цитоплазмы образовано винкулином; 2. Нити, отходящие в цитоплазму
- 93. Сравнительная таблица контактов сцепляющего типа
- 94. Контакт запирающего типа Плотный контакт Плазмолеммы клеток прилегают друг к другу вплотную, сцепляясь с помощью специальных
- 95. Белки плотного контакта Основными белками плотных контактов являются клаудины и окклюдины. Через ряд специальных белков к
- 96. Контакты коммуникационного типа Щелевидные соединения (нексусы, электрические синапсы, эфапсы) Нексус имеет форму круга диаметром 0,5-0,3 мкм.
- 97. Строение эфапса (Gap junction)
- 98. Между контактирующими клетками существует электрическая и метаболическая связи. Через каналы коннексонов могут диффундировать неорганические ионы и
- 99. Контакты коммуникационного типа Синапсы Синапсы служат для передачи сигнала от одних возбудимых клеток к другим. В
- 100. Коммуникационные соединения Встречаются в возбудимых тканях (нервная и мышечная)
- 101. Коммуникационные соединения
- 102. Плазмодесмы Представляют собой цитоплазматические мостики, соединяющие соседние клетки растений. Плазмодесмы проходят через канальцы поровых полей первичной
- 103. Фокальные контакты клеток Фокальные контакты представляют собой контакты между клетками и внеклеточным матриксом. Трансмембранными белками адгезии
- 104. Ультраструктурная основа механических свойств живых клеток Цитоскелет – ультраструктурная основа механических свойств живых клеток. Включает в
- 106. Саркомер – элементарная надмолекулярная сократительная единица мышечного волокна. Соседние саркомеры соединяются друг с другом белком десмином
- 107. Саркомер состоит из тонких (актиновых) и толстых (миозиновых) филаментов, которые называют протофибриллами. В 1 миофибрилле содержится
- 108. На поперечном срезе саркомера нити актина и миозина образуют гексагональную структуру, в которой каждая миозиновая протофибрилла
- 109. Молекула тропомиозина занимает спиральный желобок между 2-я нитями F- актина, ге расположены активные центры, с которыми
- 110. Механизм мышечного сокращения После разблокирования активного цинтра актина с помощью ионов Ca2+ миозиновые головки взаимодействуют с
- 111. Максимальная сила сокращения пропорциональна мышцы пропорциональна взаимного перекрытия актиновых и миозиновых нитей. При уменьшении длины саркомера
- 112. При повышении физической нагрузки происходит гипертрофия миоцитов, т.к. это дифференцированные клетки, не способные к делению. Для
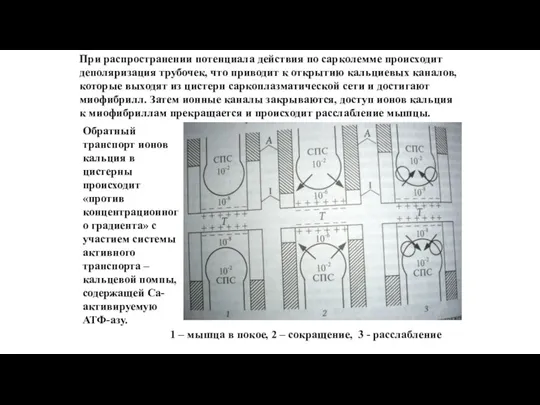
- 113. 1 – мышца в покое, 2 – сокращение, 3 - расслабление При распространении потенциала действия по
- 114. нервные импульсы, поступающие их спинного или головного мозга к к нервно-мышечному синапсу Схема нервно-мышечного сокращения Синаптическая
- 115. Метаболический аппарат клетки
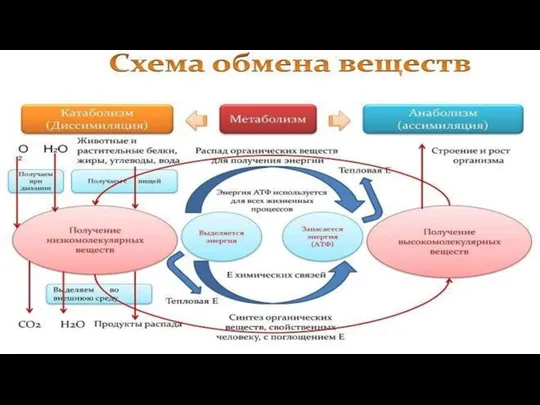
- 116. Метаболизм – это совокупность химических реакций, протекающих в живых клетках и обеспечивающих организм веществами и энергией
- 117. Структуры клетки участвующие в метаболизме Органоиды, участвующие в анаболизме: ШЭПС ГлЭПС Аппарат Гольджи Рибосомы Хлоропласты (у
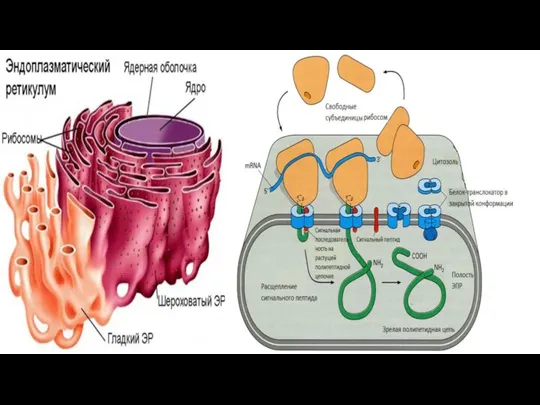
- 119. Эндоплазматический ретиккулум Эндоплазматический ретикулум (ЭПР) - это замкнутая система мембранных трубочек внутри клетки, образующих сложную переплетающуюся
- 121. Шероховатый ЭПС На рибосомах ШЭПС синтезируются следующие группы белков: Белки предназначенные для секреции-выведение через аппарат Гольджи;
- 122. Сборка белка. Этап прикрепления к ШЭПР Сборка любого белка начинается на рибосомах в цитозоле. В ЭПР
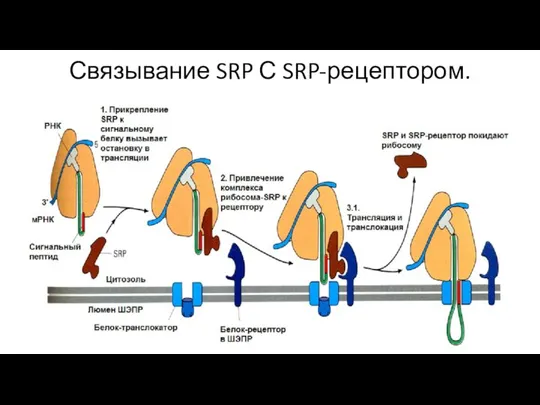
- 123. Связывание SRP С SRP-рецептором.
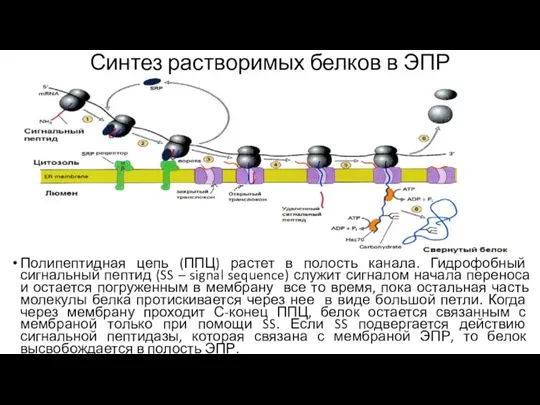
- 124. Синтез растворимых белков в ЭПР Полипептидная цепь (ППЦ) растет в полость канала. Гидрофобный сигнальный пептид (SS
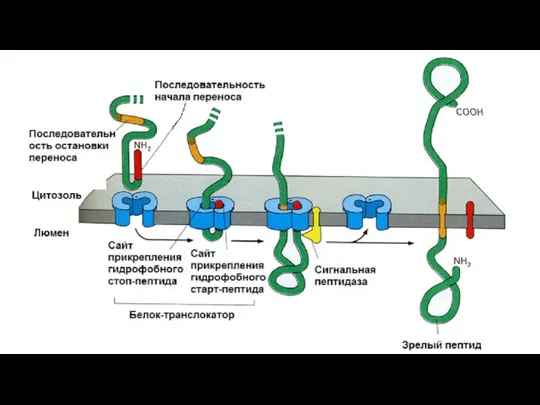
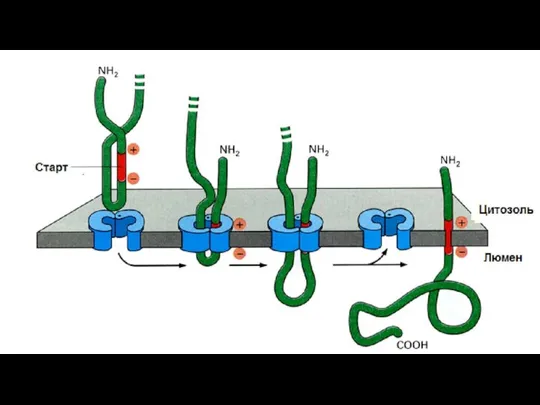
- 125. Синтез мембранных белков В синтезируемой ППЦ существует так называемая стоп-последовательность (STS – stop transport signal). Белок
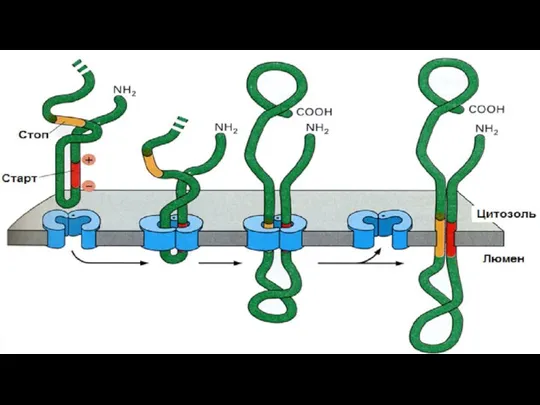
- 129. Образование политопных (то есть закрепленных в мембране несколько раз) белков. Процесс формирования ППЦ начинается как показано
- 130. Другие функции ШЭПР Большинство белков синтезированных в ШЭПР-гликопротеиды. По мере роста белковой цепи она соединяется с
- 131. Гликозилирование Почти тотчас после того, как ППЦ попадает в просвет ЭПР, она гликозилируется по доступным остаткам
- 132. Синтез липидов Синтез фосфолипидов протекает на цитоплазматической стороне мембраны ЭПР. Каждый фермент этого синтеза является интегральный
- 133. Гладкий ЭПР Очень лабилен и способен к перестройкам в зависимости от внешних условий. Например, при детоксикации
- 134. Функции ГлЭПР
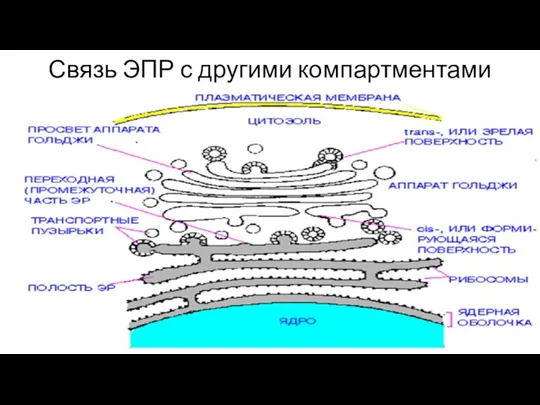
- 135. Связь ЭПР с другими компартментами
- 136. Аппарат Гольджи Аппарат Гольджи- интегрирующая часть метаболической системы эукариотической клетки. В нем происходит обособление секретируемых продуктов,
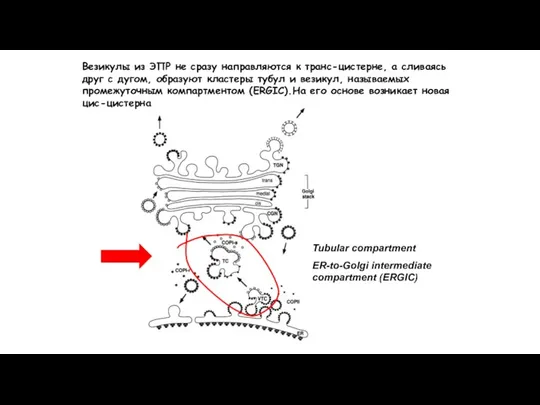
- 137. Tubular compartment ER-to-Golgi intermediate compartment (ERGIC) Везикулы из ЭПР не сразу направляются к транс-цистерне, а сливаясь
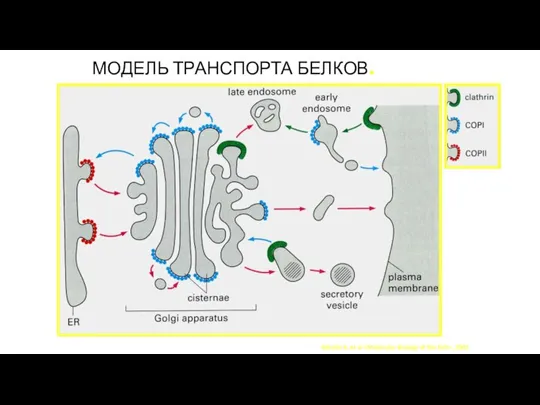
- 138. МОДЕЛЬ ТРАНСПОРТА БЕЛКОВ. Alberts B. et al «Molecular Biology of the Cell», 2002
- 139. Синтез и модификация веществ Каждая цистерна аппарата Гольджи имеет свой набор ферментов. Доработка гликопротеинов; Создание лизосомальных
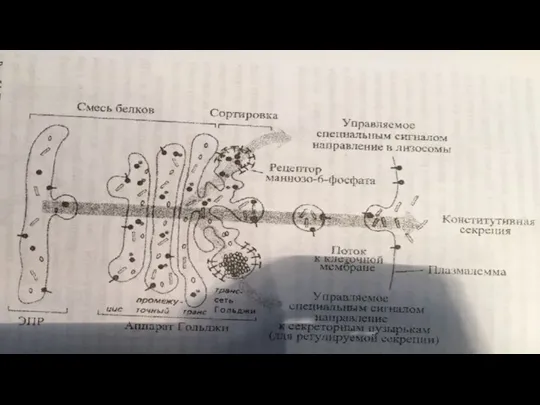
- 140. Сортировка Сортировка белков происходит в транс-сети аппарата Гольджи. Результатом является разделение белков и направление в места
- 142. Секреция Вещества выводятся из клетки одним из двух путей: в результате конструктивной (нерегулируемой) секреции (постоянная,например, путь
- 144. Функции обобщенно 1.Транспорт - через АГ проходят две группы белков: белки, предназначенные на экспорт из клетки,
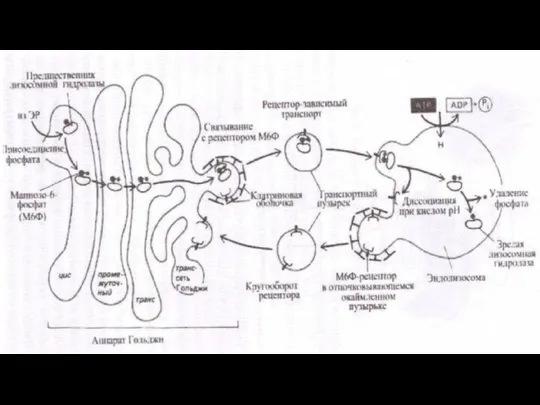
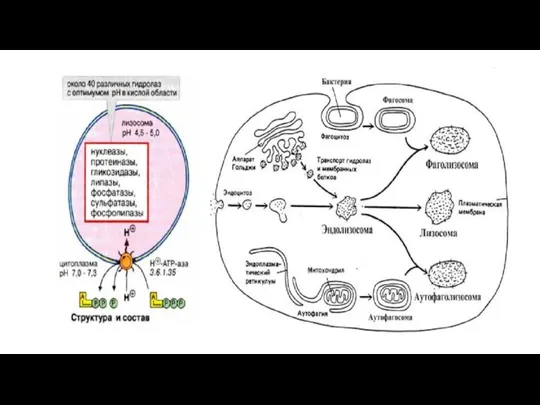
- 145. Лизосомы Лизосомы представляют собой гетерогенную (разнородную) группу цитоплазматических вакуолеподобных структур размером 1-3 мкм, отличительной особенностью которых
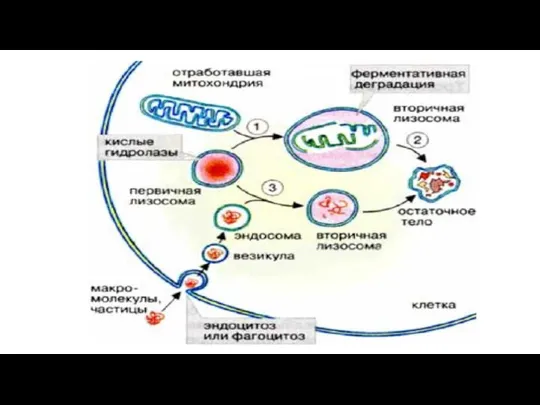
- 147. В первом случае расщепляемый материал - белки, полинуклеотиды или полисахариды - попадает в клетку путем эндоцитоза.
- 148. Второй путь формирования лизосом называется аутофагией. В процессе аутофагии происходит разрушение отработанных частей самой клетки. Известно,
- 150. Третий путь формирования лизосом имеется только у клеток, специализированных для фагоцитоза больших частиц и микроорганизмов. Такие
- 151. Эндосомы, аутофагосомы и фагосомы часто называют общим термином - прелизосомы, а эндолизосомы, аутофаголизосомы и фаголизосомы термином
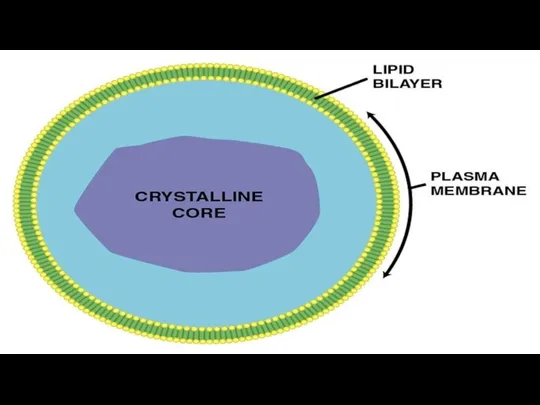
- 152. Пероксисомы Мембранные пузырьки с однородным или гранулярным матриксом. В центре-сердцевина (нуклеоид) из кристаллических структур, образованных фибриллами
- 154. Пероксисомы обнаружены во всех эукариотических клетках. Они являются главным центром утилизации кислорода (наряду с митохондрией). Кислород
- 155. Митохондрии Митохондрии, как органеллы синтеза АТФ характерны для всех эукариотических клеток как автотрофных (фотосинтезирующие растения), так
- 156. Локализация Митохондрии скапливаются в энергозависимых участках клетки. В скелетных мышцах они располагаются между микрофибриллами, у простейших
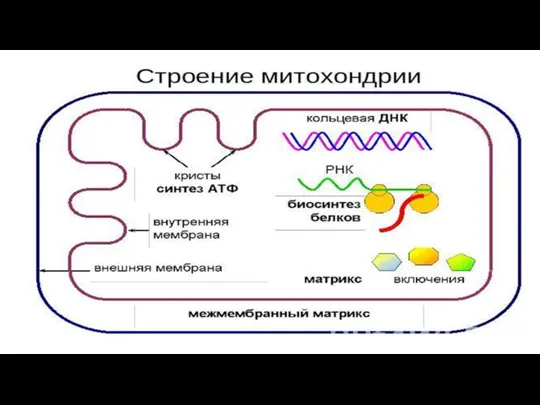
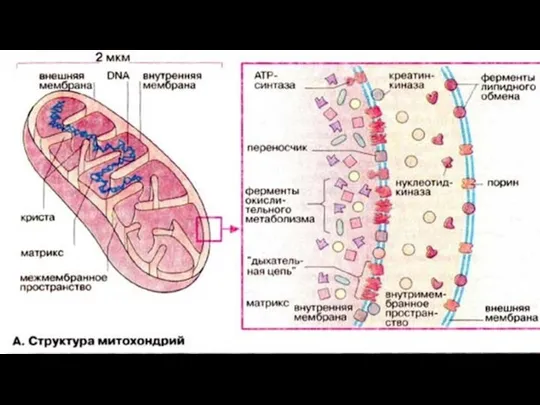
- 157. Строение Имеющиеся методы позволяют выделить из митохондрий все четыре компонента: наружную мембрану,содержимое межмембранного пространства, внутреннюю мембрану
- 160. Наружная мембрана. В состав наружной мембраны входит много молекул белка порина. Особенность его заключается в том,
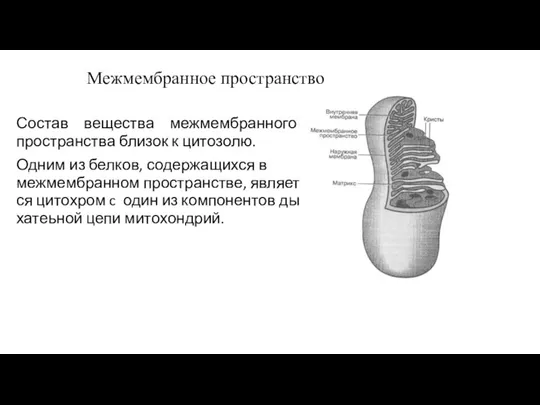
- 161. Межмембранное пространство Состав вещества межмембранного пространства близок к цитозолю. Одним из белков, содержащихся в межмембранном пространстве,
- 162. Внутренняя мембрана Внутренняя мембрана ограничивает основное рабочее пространство митохондрии. Она высокоспецифична, содержит большое количество фосфолипида кардиолипина
- 163. Матрикс Матрикс содержит высококонцентрированную смесь сотен различных ферментов, необходимых для окисления пирувата, жирных кислот и ферментов
- 164. Генетический аппарат Кольцевая молекула ДНК. Редко-линейная; Может синтезировать белок как с метохондриального иРНК, так и ядерного;
- 165. Функции Синтез АТФ; Специфический синтез-стероидные гормоны, некоторые липиды; Накопление ионов, особенно Ca2+; Накопление продуктов экскреции в
- 166. Этапы энергетического обмена: 1. Подготовительный 2. Гликолиз (бескислородное окисление) 3. Дыхание (кислородное окисление)
- 167. Первый этап Подготовительный ферментативное расщепление сложных органических веществ до простых
- 168. Где происходит: Пищеварительная система Лизосомы в клетках
- 169. Субстрат Углеводы = глюкоза + Е (1г = 17,6 кДж) Липиды = глицерин + жирные кислоты
- 170. Результат этапа Энергия не запасается, а выделяется только в тепловой форме
- 171. Гликолиз - неполное расщепление - анаэробное дыхание - брожение Второй этап Бескислородное окисление
- 172. Глюкоза – центральная молекула клеточного дыхания Полисахариды β с нее начинается путь к АТФ
- 173. 2 ПВК Клетка (под действием ферментов клеточных мембран) 10 реакций (пируват) гликолиз Где происходит:
- 174. Субстрат С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ → глюкоза 2С3Н4О3 + 2АТФ + 2Н2О +
- 175. 60% выделяется в виде тепла 40% идет на синтез АТФ Энергия Результат этапа: из одной молекулы
- 176. ГЛЮКОЗА П В К 2 АТФ Брожение – анаэробное дыхание Если мало кислорода или организм –
- 177. Выводы: Синтез АТФ в процессе гликолиза не нуждается в мембранах. Он идёт даже в пробирке, если
- 178. Третий этап Кислородное расщепление: полное расщепление пировиноградной кислоты, происходит при обязательном присутствии кислорода
- 179. О2 Митохондрия: под действием ферментов митохондриальных мембран (необходимое условие – целостность мембран) ПВК СО2 и Н2О
- 180. Стадии аэробного дыхания: 1) Окислительное декарбоксилирование 2) Цикл Кребса 3) Электронтранспортная цепь (окислительное фосфолирирование)
- 182. Электронтранспортная цепь
- 183. АТФ-ситаза Принимает электроны от цитохрома с и передает их на кислород с образованием воды. Переносит 2
- 185. Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану
- 186. Результат этапа: 2600 кДж - на 2 моля С3Н4О3 45% Рассеивается в виде тепла Сберегается в
- 187. Выводы: Для осуществления кислородного процесса необходимо наличие неповреждённых митохондриальных мембран
- 188. Выводы: Расщепление в клетке 1 молекулы глюкозы до СО2 и Н2О обеспечивает синтез 38 молекул АТФ
- 189. Ядерный аппарат
- 190. Состав ядерного аппарата: Поверхностный аппарат ядра; Ядерный матрикс; Кариоплазма; Хроматин; Ядрышко.
- 191. 1 — ядерная оболочка (две мембраны - внутренняя и внешняя, и перинуклеарное пространство); 2 — ядерная
- 192. Поверхностный аппарат Ядерная оболочка с порами: Разделена на внешнюю и внутреннюю ядерные оболочки. Пространство между ними
- 193. Поровый комплекс
- 194. РНК должны попасть из ядра в цитоплазму. Для этого нужно преодолеть ядерную мембрану. Мембрана пронизана большим
- 195. Функции поверхностного аппарата Отграничение ядра от цитоплазмы. В результате этого происходит разделение транскрипции и трансляции-возможен процессинг.
- 196. Ядерный матрикс Ядерный матрикс-это скелет ядра. Белки связывают определенные последовательности ДНК, образующие основание петель хроматина. Благодаря
- 197. Ядрышко Ядрышко – высокоорганизованная шаровидная структура ядра. Содержит большие петли ДНК, выступающие из хромосом, при этом
- 198. Хроматин Хроматин: основными компонентами являютсяДНК,гистоны инегистоновые белки (ДНП). Соотношение ДНК и белка составляет ~1:1,3, а основная
- 199. Урони упаковки хроматина
- 200. Урони упаковки хроматина Нуклеосомный уровень-8 молекул гистонов закручивают фрагмент 140 пар оснований. Линкерный участок-60 пар оснований.
- 202. Аминокислотые остатки гистонов могут подвергаться пост-трансляционным модификациям: Ацетилирование по определённым остаткам лизина. —Метилирование определённых остатков лизина
- 204. Метилирование ДНК
- 206. Что происходит? Метильные группы нарушают ДНК-белковые взаимодействия, выступая в большую бороздку ДНК и препятствуя связыванию специфических
- 207. Архитектурные белки высокомобильной группы (HMG) Были названы так потому, что обладают высокой электрофоретической мобильностью в полиакриламидном
- 208. Хроматин-перестраивающие комплексы В середине 90-хгодов стало известно, что хроматин содержит комплексы, перестраивающие нуклеосомы и использующие энергию
- 210. Функция белков хроматина Определяют уровень компактизации ДНК; Влияют на уровень транскрипции; Осуществляют процесс транскрипции.
- 211. Функции ядра Хранение и передача генетической информации. Реализация генетической информации.
- 212. Основы молекулярной биологии клетки
- 213. Что такое молекулярная биология? Жизнь — способ существования биополимеров, из которых основными являются белки и нуклеиновые
- 214. НУКЛЕОТИД Остаток фосфорной кислоты пентоза Азотистое основание
- 215. Рибоза Дезоксирибоза
- 216. Нуклеиновые кислоты Нуклеиновые кислоты выполняют генетическую функцию (ДНК) и участвуют в реализации генетической информации (РНК). Молекулы
- 218. Связи в нуклеотидах Гликозидная связь— это тип ковалентной связи, которая соединяет молекулу сахара с другой молекулой.
- 219. Производные нуклиотидов Нуклиотиды, помимо того,что являются мономерами ДНК и РНК, учавствуют в большом количестве других важных
- 220. Эксперимент Гриффита на пневмококках
- 221. Трансдукция
- 222. Строение ДНК Нуклеотиды одной цепи соединяются связями, возникающими между остатком фосфорной кислоты и углеводом через фосфат,
- 224. Двойная спираль ДНК
- 225. Комплементарность и правила Чаргаффа Комплементарность - это принцип взаимного соответствия парных нуклеотидов или способность нуклеотидов объединяться
- 226. Функции ДНК
- 227. РНК Молекула РНК представляет собой неразветвленный полинуклеотид, который может иметь первичную структуру – последовательность нуклеотидов, вторичную
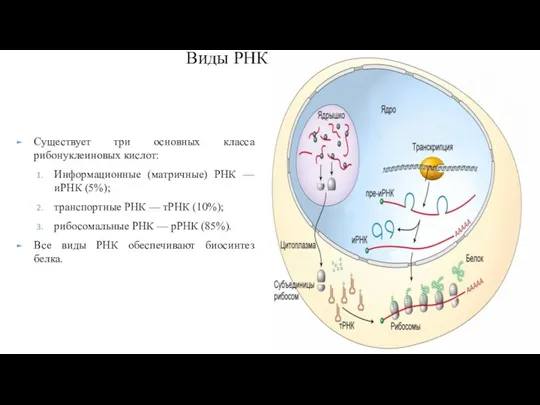
- 228. Виды РНК Существует три основных класса рибонуклеиновых кислот: Информационные (матричные) РНК — иРНК (5%); транспортные РНК
- 229. Информационная РНК. Наиболее разнообразный по размерам и стабильности класс. Все они являются переносчиками генетической информации из
- 230. тРНК Молекулы тРНК— сравнительно короткие молекулы, состоящие из 71–94 нуклеотидов. тРНК приходится до 10% от общего
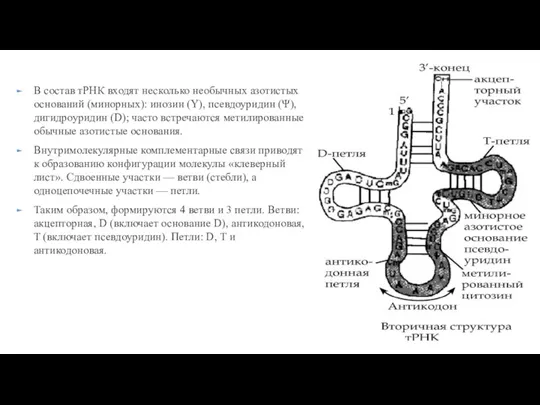
- 231. В состав тРНК входят несколько необычных азотистых оснований (минорных): инозин (Y), псевдоуридин (Ψ), дигидроуридин (D); часто
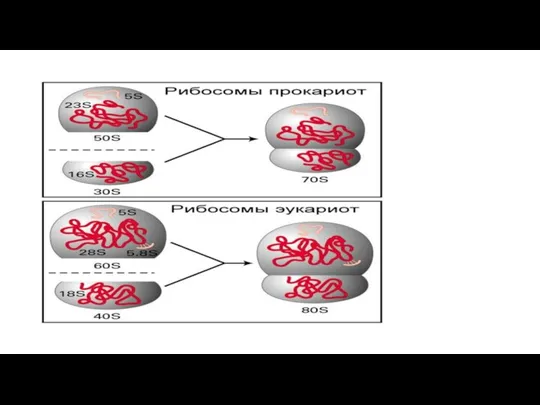
- 232. Рибосомы В состав субъединиц рибосом входят разнообразные рРНК, различающиеся по константе седиментации (единица Сведберга — S).
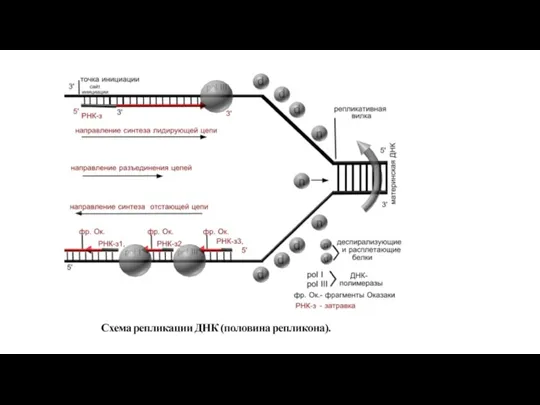
- 234. Репликация Самовоспроизведение ДНК (процесс, приводящий к удвоению молекул ДНК), называется репликацией. Репликация необходима перед делением клетки,
- 235. Участок молекулы ДНК, на котором идет репликация, называется репликон. У прокариот репликон один, а у эукариот
- 236. Начинается процесс с разрыва водородных связей между азотистыми основаниями ДНК на участке, включающем около 300 пар
- 237. На другой половине репликона, где разъединение цепей ДНК идёт в другую сторону, также, в одном направлении
- 239. Повреждения ДНК Спонтанные повреждения ДНК: Ошибки репликации (появление некомплементарных пар нуклеотидов); Апуринизация (отщепление азотистых оснований из
- 240. Фоторепарация 1. Нормальная молекула ДНК ↓ Облучение УФ-светом ↓ 2. Мутантная молекула ДНК – образование пиримидиновых
- 242. Эксцизионная репарация 1. Узнавание повреждения ДНК эндонуклеазой; 2. Инцизия (надрезание) цепи ДНК ферментом по обе стороны
- 243. Пострепликативная репарация ДНК Пострепликативная репарация осуществляется в тех случаях, когда повреждение доживает до фазы репликации (слишком
- 245. Скачать презентацию
Слайд 2Введение
Живая природа подчиняется иерархической организации. Иерархический принцип организации позволяет выделить в живой
Введение
Живая природа подчиняется иерархической организации. Иерархический принцип организации позволяет выделить в живой

Слайд 3Микроскоп - (от греческого mikros - малый и skopeo - смотрю), оптический прибор
Микроскоп - (от греческого mikros - малый и skopeo - смотрю), оптический прибор


Слайд 5Световая микроскопия. Устройство светового микроскопа
Изучение препарата осуществляется в проходящем свете с помощью
Световая микроскопия. Устройство светового микроскопа
Изучение препарата осуществляется в проходящем свете с помощью

Слайд 6Основными частями светового микроскопа являются объектив и окуляр, заключенные в цилиндрический корпус
Основными частями светового микроскопа являются объектив и окуляр, заключенные в цилиндрический корпус

Слайд 7Основание микроскопа
Тубусодержатель
Тубус
Окуляр (чаще ×10)
Револьвер микроскопа
Объективы
а) сухие: ×10, ×20, ×40
б) иммерсионные х60, ×90
Предметный
Основание микроскопа
Тубусодержатель
Тубус
Окуляр (чаще ×10)
Револьвер микроскопа
Объективы
а) сухие: ×10, ×20, ×40
б) иммерсионные х60, ×90
Предметный

Слайд 8Характеристики оптического микроскопа
К основным характеристикам микроскопа относятся увеличение и разрешающая способность.
Общее
Характеристики оптического микроскопа
К основным характеристикам микроскопа относятся увеличение и разрешающая способность.
Общее

Слайд 9Электронная микроскопия. Устройство электронного микроскопа
Электронный микроскоп — прибор, позволяющий получать изображение объектов
Электронная микроскопия. Устройство электронного микроскопа
Электронный микроскоп — прибор, позволяющий получать изображение объектов

Слайд 10Электронный микроскоп состоит из электронной пушки (устройства для получения пучка электронов) и
Электронный микроскоп состоит из электронной пушки (устройства для получения пучка электронов) и

Слайд 11При поляризационной микроскопии на объект исследования направляется поляризованный пучок света. Это обеспечивает
При поляризационной микроскопии на объект исследования направляется поляризованный пучок света. Это обеспечивает
Слайд 12Темнопольной микроскопии. Основана на использовании специального конденсора, освещающего препарат «косыми» лучами, не
Темнопольной микроскопии. Основана на использовании специального конденсора, освещающего препарат «косыми» лучами, не
Слайд 13Метод фазово-контрастной микроскопии служит для получения контрастных изображений прозрачных и бесцветных объектов.
Метод фазово-контрастной микроскопии служит для получения контрастных изображений прозрачных и бесцветных объектов.

Слайд 14В флуоресцентной микроскопии используется принцип свечения объекта исследования при освещении его ультрафиолетовыми
В флуоресцентной микроскопии используется принцип свечения объекта исследования при освещении его ультрафиолетовыми
Слайд 15Правила работы с микроскопом
Установите микроскоп слева, штативом к себе;
Поставьте в рабочее положение
Правила работы с микроскопом
Установите микроскоп слева, штативом к себе;
Поставьте в рабочее положение

Слайд 16Поверхностный аппарат клетки
Поверхностный аппарат клетки

Слайд 17Определение клетки, ее компоненты
Клетка - целостная элементарная система, способная к самовоспроизведению и
Определение клетки, ее компоненты
Клетка - целостная элементарная система, способная к самовоспроизведению и

Слайд 18Модели мембран
Липидный слой (Овертон, 1902)
Билипидный слой
(Грендель, 1925)
«Бутербродная» модель
(Даниэли и Девсон,
Модели мембран
Липидный слой (Овертон, 1902)
Билипидный слой
(Грендель, 1925)
«Бутербродная» модель
(Даниэли и Девсон,
Слайд 19Поверхностный аппарат клетки
Поверхностный аппарат клетки

Слайд 20Плазматическая мембрана образована белками (-60%) и липидами (-40%).
В состав липидов входят
Плазматическая мембрана образована белками (-60%) и липидами (-40%).
В состав липидов входят

Слайд 21Свойства плазматической мембраны
Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою
Свойства плазматической мембраны
Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою


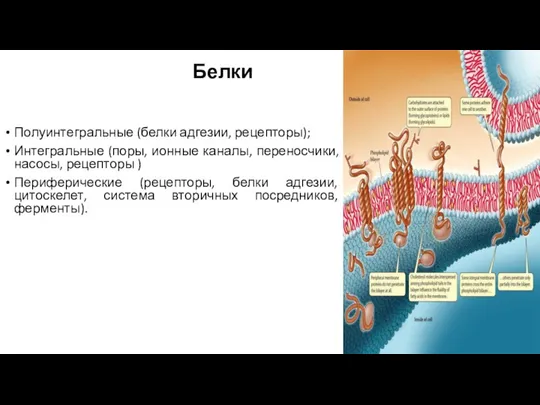
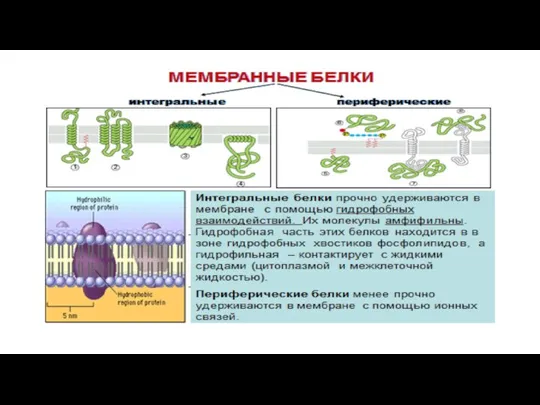
Слайд 23 Белки
Полуинтегральные (белки адгезии, рецепторы);
Интегральные (поры, ионные каналы, переносчики, насосы, рецепторы )
Периферические
Белки
Полуинтегральные (белки адгезии, рецепторы);
Интегральные (поры, ионные каналы, переносчики, насосы, рецепторы )
Периферические
Слайд 25Функциональная классификация мембранных белков
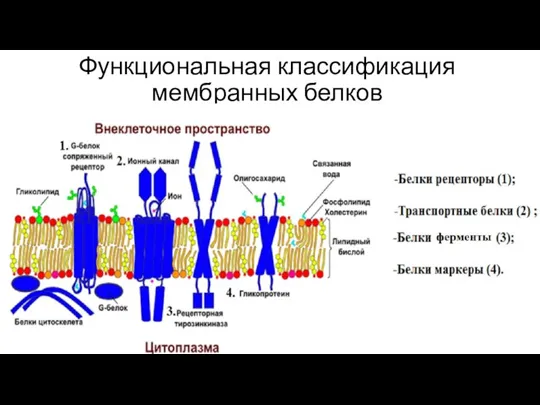
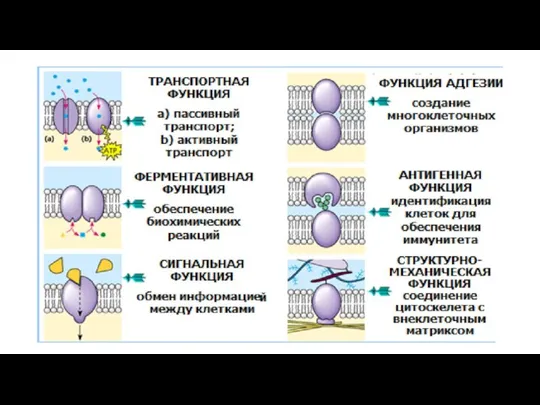
Функциональная классификация мембранных белков
Слайд 27Липиды мембраны
Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом.
Фосфолипиды в
Липиды мембраны
Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом.
Фосфолипиды в

Слайд 28Фосфолипиды
Фосфолипиды являются неотъемлемым компонентом всех клеточных мембран . Между плазмой и эритроцитами
Фосфолипиды
Фосфолипиды являются неотъемлемым компонентом всех клеточных мембран . Между плазмой и эритроцитами
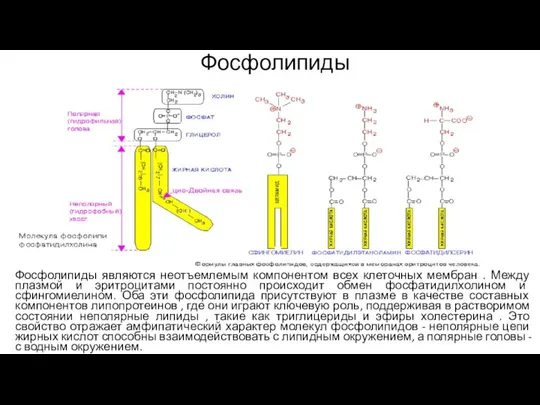
Слайд 29Гликолипиды
Гликолипиды составляют 5% липидных молекул наружного монослоя и сильно различаются у разных
Гликолипиды
Гликолипиды составляют 5% липидных молекул наружного монослоя и сильно различаются у разных

Слайд 30Холестерин
Холестерин - это стерин, содержащий стероидное ядро из четырех колец и гидроксильную
Холестерин
Холестерин - это стерин, содержащий стероидное ядро из четырех колец и гидроксильную

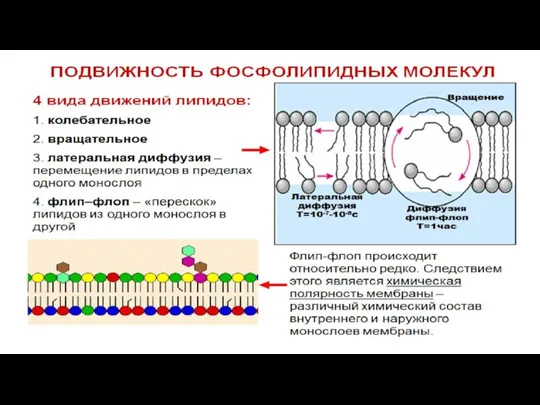
Слайд 32ВЛИЯНИЕ ХИМИЧЕСКОГО СОСТАВА
Зависимость свойств мембраны от химического состава называется лиотропным полиморфизмом.
Ненасыщенных
ВЛИЯНИЕ ХИМИЧЕСКОГО СОСТАВА
Зависимость свойств мембраны от химического состава называется лиотропным полиморфизмом.
Ненасыщенных

Слайд 33ТРАНСПОРТ ВЕЩЕСТВ В МЕМБРАНАХ
ТРАНСПОРТ ВЕЩЕСТВ В МЕМБРАНАХ
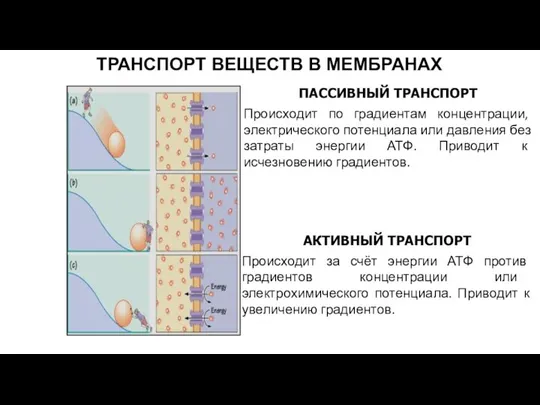
Слайд 34 Виды транспорта
Виды транспорта
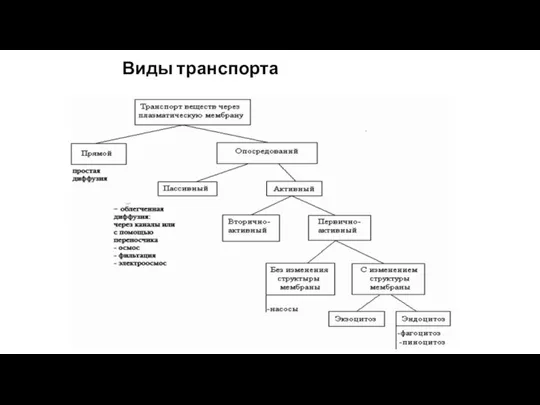
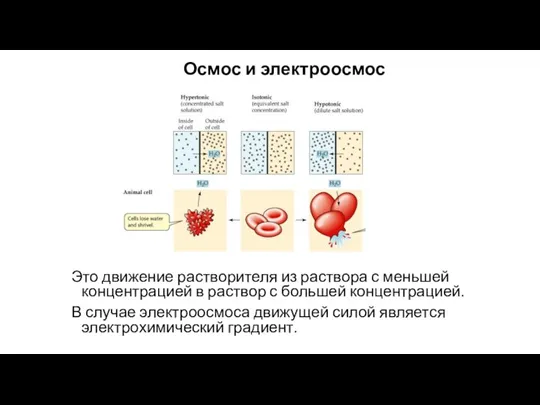
Слайд 36 Осмос и электроосмос
Это движение растворителя из раствора с меньшей концентрацией в
Осмос и электроосмос
Это движение растворителя из раствора с меньшей концентрацией в
Слайд 37Фильтрация
Это движение растворителя под действием гидростатического давления
Фильтрация
Это движение растворителя под действием гидростатического давления

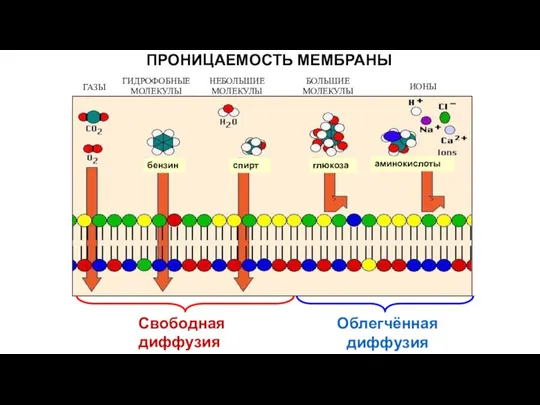
Слайд 38ПРОНИЦАЕМОСТЬ МЕМБРАНЫ
Свободнаядиффузия
Облегчённая диффузия
ПРОНИЦАЕМОСТЬ МЕМБРАНЫ
Свободнаядиффузия
Облегчённая диффузия
Слайд 39 Простая диффузия
Линейно зависит от градиента концентрации вещества;
Характеризуется ненасыщаемостью.
Простая диффузия
Линейно зависит от градиента концентрации вещества;
Характеризуется ненасыщаемостью.

Слайд 40 Простая диффузия осуществляется через:
Мембрану (для незаряженных жирорастворимых) веществ;
Поры;
Кинки.
Простая диффузия осуществляется через:
Мембрану (для незаряженных жирорастворимых) веществ;
Поры;
Кинки.

Слайд 41Простая диффузия идет через кинки
При повышении температуры тепловая подвижность жирнокислотных цепей приводит
Простая диффузия идет через кинки
При повышении температуры тепловая подвижность жирнокислотных цепей приводит

Слайд 42 Простая диффузия через поры
Канал поры всегда открыт, поэтому химическое вещество проходит
Простая диффузия через поры
Канал поры всегда открыт, поэтому химическое вещество проходит
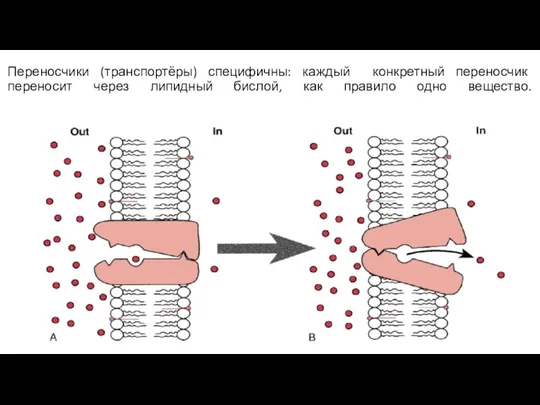
Слайд 43Переносчики (транспортёры) специфичны: каждый конкретный переносчик переносит через липидный бислой, как правило
Переносчики (транспортёры) специфичны: каждый конкретный переносчик переносит через липидный бислой, как правило
Слайд 44ЭЛЕКТРОДИФФУЗИЯ
;
ЭЛЕКТРОДИФФУЗИЯ
;

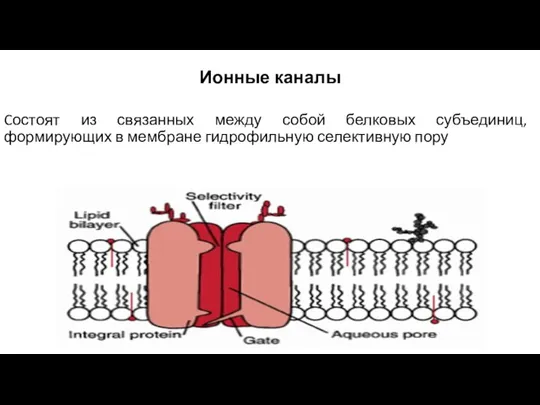
Слайд 45Ионные каналы
Cостоят из связанных между собой белковых субъединиц, формирующих в мембране гидрофильную
Ионные каналы
Cостоят из связанных между собой белковых субъединиц, формирующих в мембране гидрофильную
Слайд 46Свойства ионных каналов
Специфичность
Проводимость
Наличие сенсора
Наличие воротной системы
Свойства ионных каналов
Специфичность
Проводимость
Наличие сенсора
Наличие воротной системы

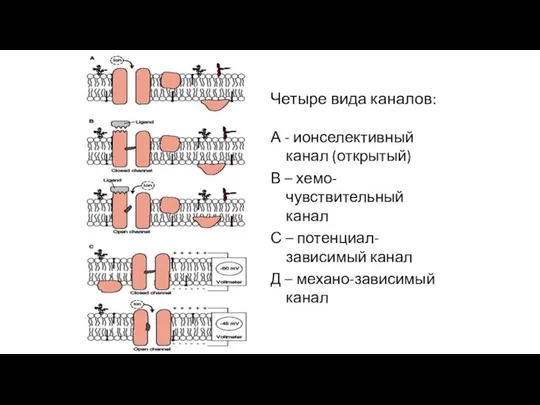
Слайд 47
Четыре вида каналов:
А - ионселективный канал (открытый)
В – хемо-чувствительный канал
С – потенциал-зависимый
Четыре вида каналов:
А - ионселективный канал (открытый)
В – хемо-чувствительный канал
С – потенциал-зависимый
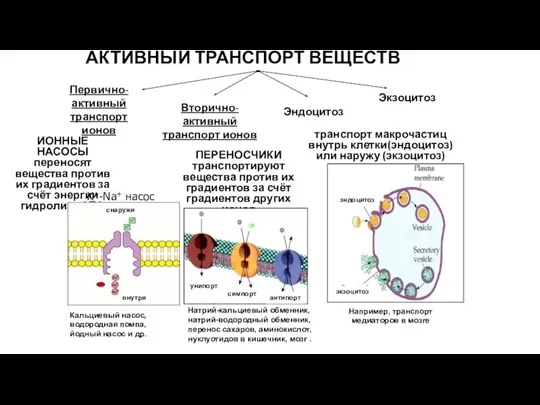
Слайд 48ИОННЫЕ НАСОСЫ
переносят вещества против их градиентов за счёт энергии гидролиза АТФ
ПЕРЕНОСЧИКИ
ИОННЫЕ НАСОСЫ
переносят вещества против их градиентов за счёт энергии гидролиза АТФ
ПЕРЕНОСЧИКИ
Слайд 49Активный транспорт
Осуществляется против электрохимического градиента
Система в высшей степени специфична
Необходимы источники энергии
Активный транспорт
Осуществляется против электрохимического градиента
Система в высшей степени специфична
Необходимы источники энергии

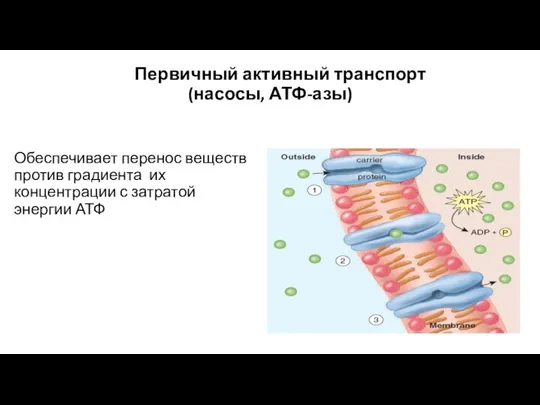
Слайд 50 Первичный активный транспорт
(насосы, АТФ-азы)
Обеспечивает перенос веществ против градиента их концентрации с
Первичный активный транспорт
(насосы, АТФ-азы)
Обеспечивает перенос веществ против градиента их концентрации с
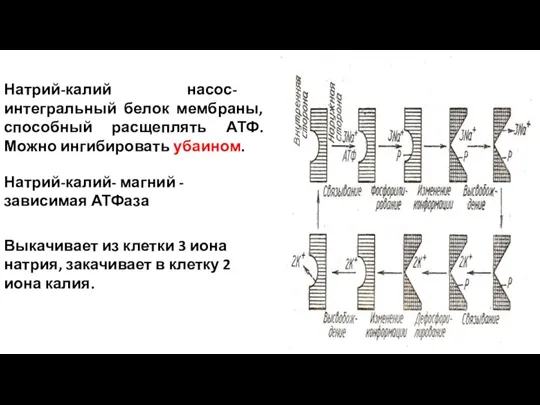
Слайд 51Натрий-калий- магний - зависимая АТФаза
Натрий-калий насос- интегральный белок мембраны, способный расщеплять АТФ.
Натрий-калий- магний - зависимая АТФаза
Натрий-калий насос- интегральный белок мембраны, способный расщеплять АТФ.
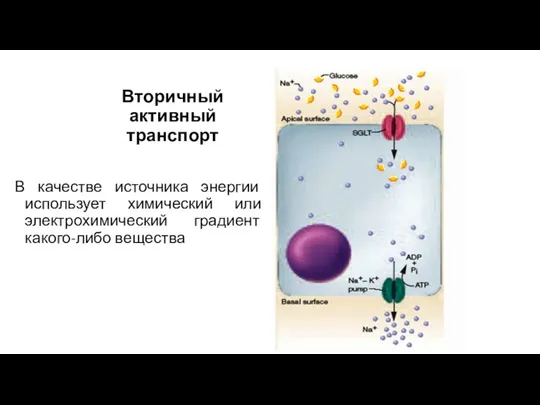
Слайд 52Вторичный активный транспорт
В качестве источника энергии использует химический или электрохимический градиент какого-либо
Вторичный активный транспорт
В качестве источника энергии использует химический или электрохимический градиент какого-либо
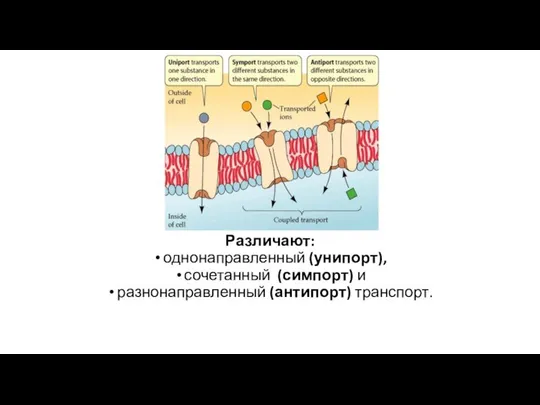
Слайд 53Различают:
однонаправленный (унипорт),
сочетанный (симпорт) и
разнонаправленный (антипорт) транспорт.
Различают:
однонаправленный (унипорт),
сочетанный (симпорт) и
разнонаправленный (антипорт) транспорт.
Слайд 54Эндоцитоз
Образуется окаймленная везикула (3),
Везикула сливается с эндосомой(4).
Вакуоль и ее содержимое претерпевают
Эндоцитоз
Образуется окаймленная везикула (3),
Везикула сливается с эндосомой(4).
Вакуоль и ее содержимое претерпевают

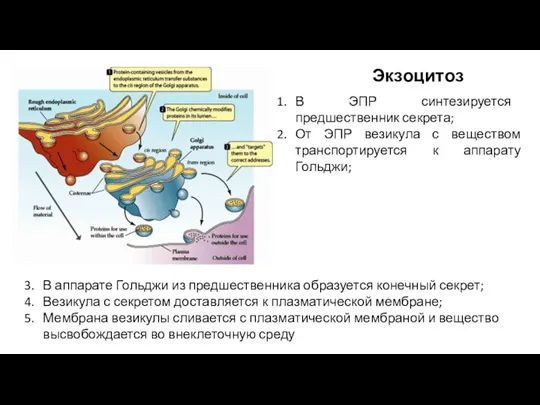
Слайд 55Экзоцитоз
В аппарате Гольджи из предшественника образуется конечный секрет;
Везикула с секретом доставляется к
Экзоцитоз
В аппарате Гольджи из предшественника образуется конечный секрет;
Везикула с секретом доставляется к
Слайд 56Рецепторная функция. Постоянные межклеточные контакты. Цитоскелет
Рецепторная функция. Постоянные межклеточные контакты. Цитоскелет

Слайд 57Этапы проведения сигнала
1) Прием сигнала (рецепция)
2) Обработка и передача сигнала
(системы сигнальных
Этапы проведения сигнала
1) Прием сигнала (рецепция)
2) Обработка и передача сигнала
(системы сигнальных

Слайд 58Стимулы
(информация, приходящая к клетке)
1) Высокоаффинные лиганды (связываются с
рецептором высокоаффинно и высокоселективно)
2)
Стимулы
(информация, приходящая к клетке)
1) Высокоаффинные лиганды (связываются с
рецептором высокоаффинно и высокоселективно)
2)

Слайд 59Ответы клетки
Влияние на пролиферацию (стимуляция деления клеток / блок клеточного цикла);
Влияние на
Ответы клетки
Влияние на пролиферацию (стимуляция деления клеток / блок клеточного цикла);
Влияние на

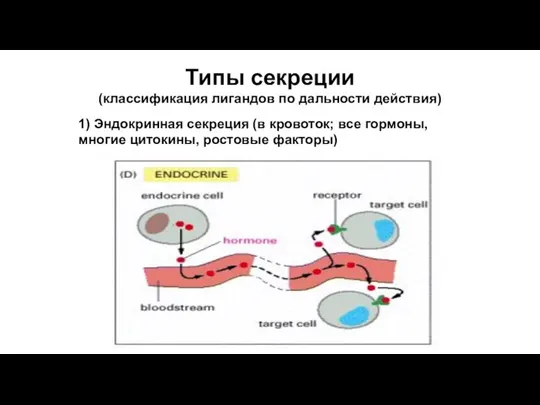
Слайд 60Типы секреции
(классификация лигандов по дальности действия)
1) Эндокринная секреция (в кровоток; все гормоны,
многие
Типы секреции
(классификация лигандов по дальности действия)
1) Эндокринная секреция (в кровоток; все гормоны,
многие
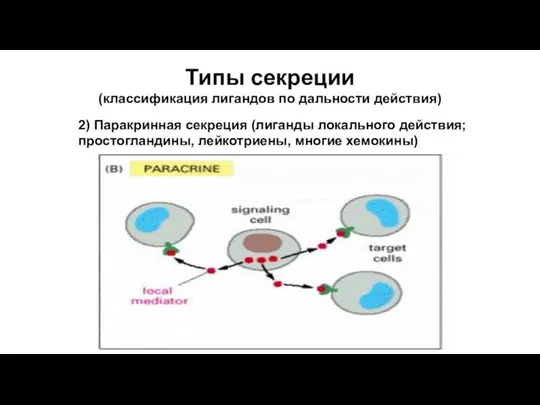
Слайд 61Типы секреции
(классификация лигандов по дальности действия)
2) Паракринная секреция (лиганды локального действия;
простогландины, лейкотриены,
Типы секреции
(классификация лигандов по дальности действия)
2) Паракринная секреция (лиганды локального действия;
простогландины, лейкотриены,
Слайд 62Типы секреции
(классификация лигандов по дальности действия)
3) Аутокринная секреция (действие на ту же
Типы секреции
(классификация лигандов по дальности действия)
3) Аутокринная секреция (действие на ту же

Слайд 63Типы секреции
(классификация лигандов по дальности действия)
4) Юкстакринная, или зависимая от контактов секреция
Типы секреции
(классификация лигандов по дальности действия)
4) Юкстакринная, или зависимая от контактов секреция

Слайд 64Типы секреции
(классификация лигандов по дальности действия)
5) Секреция компонентов внеклеточного матрикса
(нерастворимые лиганды
Типы секреции
(классификация лигандов по дальности действия)
5) Секреция компонентов внеклеточного матрикса
(нерастворимые лиганды

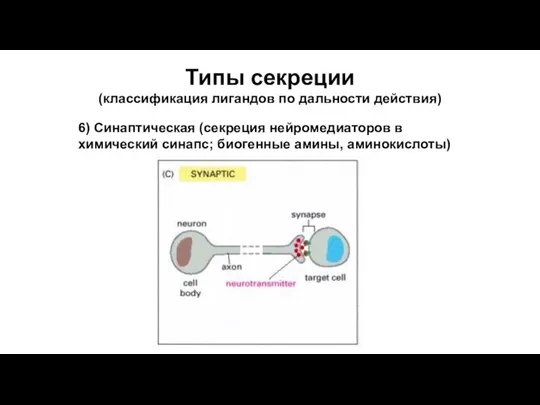
Слайд 65Типы секреции
(классификация лигандов по дальности действия)
6) Синаптическая (секреция нейромедиаторов в
химический синапс;
Типы секреции
(классификация лигандов по дальности действия)
6) Синаптическая (секреция нейромедиаторов в
химический синапс;
Слайд 66Типы секреции
(классификация лигандов по дальности действия)
7) Секреция через щелевые контакты (gap junctions;
Типы секреции
(классификация лигандов по дальности действия)
7) Секреция через щелевые контакты (gap junctions;

Слайд 67Типы секреции
(классификация лигандов по дальности действия)
6) Синаптическая (секреция нейромедиаторов в
химический синапс;
Типы секреции
(классификация лигандов по дальности действия)
6) Синаптическая (секреция нейромедиаторов в
химический синапс;
Слайд 68Клетки могут взаимодействовать друг с другом по средствам:
На расстоянии через секрецию определенных
Клетки могут взаимодействовать друг с другом по средствам:
На расстоянии через секрецию определенных

Слайд 69Рецепторы
Рецепторы
Слайд 70Внутриклеточные рецепторы
Внутриклеточные рецепторы
Слайд 71Цикл работы рецептора связанного с G-белком
Субъединица альфа умеет расщеплять ГТФ. В активированном
Цикл работы рецептора связанного с G-белком
Субъединица альфа умеет расщеплять ГТФ. В активированном

Слайд 72Мембранно-внутриклеточный механизм Вторичные посредники
цАМФ - циклический аденозинмонофосфат;
цГМФ - циклический гуанозинмонофосфат;
ИФ3 - инозитол-3-фосфат;
ДАГ
Мембранно-внутриклеточный механизм Вторичные посредники
цАМФ - циклический аденозинмонофосфат;
цГМФ - циклический гуанозинмонофосфат;
ИФ3 - инозитол-3-фосфат;
ДАГ

Слайд 73Вариант 1. (с участием аденилатциклазной системы)
Вторичный посредник - цАМФ;
Мембранно-связанный фермент - аденилатциклаза
Гормон
Вариант 1. (с участием аденилатциклазной системы)
Вторичный посредник - цАМФ;
Мембранно-связанный фермент - аденилатциклаза
Гормон

Слайд 74Передача гормонального сигнала в клетку через аденилатциклазную систему
Передача гормонального сигнала в клетку через аденилатциклазную систему
Слайд 75Прекращение влияния цАМФ на процесс активации протеинкиназы А и, следовательно, фосфорилирования белков
Прекращение влияния цАМФ на процесс активации протеинкиназы А и, следовательно, фосфорилирования белков

Слайд 76Вариант 2. (с участием инозитолфосфатной системы)
Вторичные посредники- инозитол-3-фосфат (ИФ3), ДАГ
Образуются путем гидролиза
Вариант 2. (с участием инозитолфосфатной системы)
Вторичные посредники- инозитол-3-фосфат (ИФ3), ДАГ
Образуются путем гидролиза

Слайд 77Основные этапы передачи гормонального сигнала
1.Связывание гормона с рецептором меняется конформация рецептора.
2.Изменение
Основные этапы передачи гормонального сигнала
1.Связывание гормона с рецептором меняется конформация рецептора.
2.Изменение

Слайд 78Передача сигнала в клетку через инозитолфосфатную систему
Передача сигнала в клетку через инозитолфосфатную систему
Слайд 79
Вариант 3. (с участием рецепторов с тирозинкиназной активностью)
Рецептор- фермент- протеинкиназа.
Субъединицы фермента
Вариант 3. (с участием рецепторов с тирозинкиназной активностью)
Рецептор- фермент- протеинкиназа.
Субъединицы фермента
Слайд 80Клеточные контакты
Клетка – клетка
1) Контакты простого типа:
а) адгезионные
б) интердигитация (пальцевые соединения)
2) контакты
Клеточные контакты
Клетка – клетка
1) Контакты простого типа:
а) адгезионные
б) интердигитация (пальцевые соединения)
2) контакты
Слайд 81Архитектурные типы тканей
Эпителиальные
Много клеток – мало межклеточного вещества
Межклеточные контакты
Соединительные
Много межклеточного вещества –
Архитектурные типы тканей
Эпителиальные
Много клеток – мало межклеточного вещества
Межклеточные контакты
Соединительные
Много межклеточного вещества –
Слайд 82Общая схема строения клеточных контактов
Межклеточные контакты образуются по следующей схеме:
Элемент цитоскелета
(актиновые- или
Общая схема строения клеточных контактов
Межклеточные контакты образуются по следующей схеме:
Элемент цитоскелета
(актиновые- или

Слайд 83Контакты простого типа
Адгезионные соединения
Это простое сближение плазмолемм соседних клеток на расстояние 15-20
Контакты простого типа
Адгезионные соединения
Это простое сближение плазмолемм соседних клеток на расстояние 15-20

Слайд 84Интердигитация (пальцевидное соединение) (№ 2 на рисунке) представляет собой контакт, при котором
Интердигитация (пальцевидное соединение) (№ 2 на рисунке) представляет собой контакт, при котором

Слайд 85Контакты простого типа
Встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки
Контакты простого типа
Встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки
Слайд 86Контакты сцепляющего типа
Десмосомы
Полудесмосомы
Поясок
сцепления
Контакты сцепляющего типа
Десмосомы
Полудесмосомы
Поясок
сцепления
Слайд 87Контакты сцепляющего типа
Десмосома
Десмосома представляет собой небольшое округлое образование, содержащее специфические внутри- и
Контакты сцепляющего типа
Десмосома
Десмосома представляет собой небольшое округлое образование, содержащее специфические внутри- и
Слайд 88В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены – за
В области десмосомы плазмолеммы обеих клеток с внутренней стороны утолщены – за
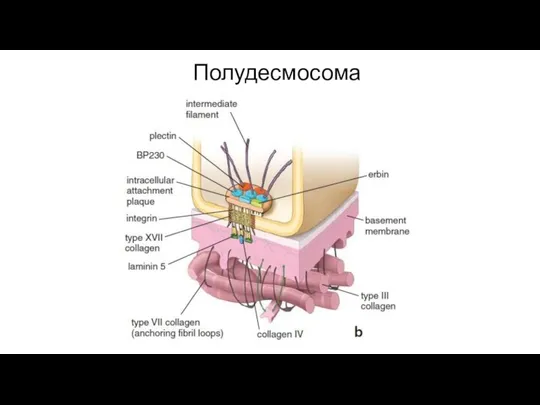
Слайд 89Полудесмосома
Полудесмосома обеспечивает контакт клеток с базальной мембраной.
По структуре гемидесмосомы напоминают десмосомы и
Полудесмосома
Полудесмосома обеспечивает контакт клеток с базальной мембраной.
По структуре гемидесмосомы напоминают десмосомы и

Слайд 90Полудесмосома
Полудесмосома
Слайд 91Поясок сцепления
Адгезивный поясок, (поясок сцепления, поясная десмосома) (zonula adherens), – парное образование
Поясок сцепления
Адгезивный поясок, (поясок сцепления, поясная десмосома) (zonula adherens), – парное образование

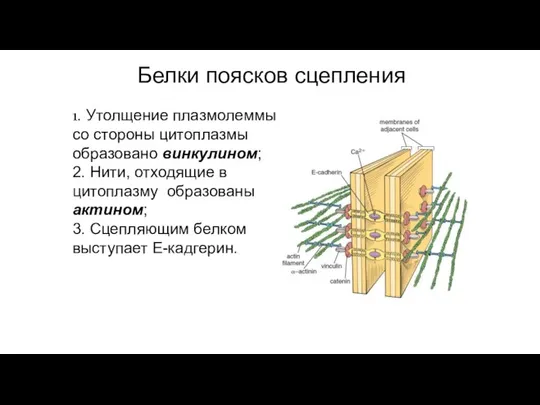
Слайд 92Белки поясков сцепления
1. Утолщение плазмолеммы со стороны цитоплазмы образовано винкулином;
2. Нити, отходящие
Белки поясков сцепления
1. Утолщение плазмолеммы со стороны цитоплазмы образовано винкулином;
2. Нити, отходящие
Слайд 93Сравнительная таблица контактов сцепляющего типа
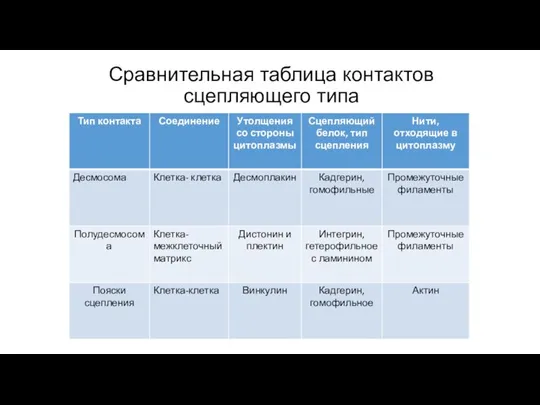
Сравнительная таблица контактов сцепляющего типа
Слайд 94Контакт запирающего типа
Плотный контакт
Плазмолеммы клеток прилегают друг к другу вплотную, сцепляясь с
Контакт запирающего типа
Плотный контакт
Плазмолеммы клеток прилегают друг к другу вплотную, сцепляясь с

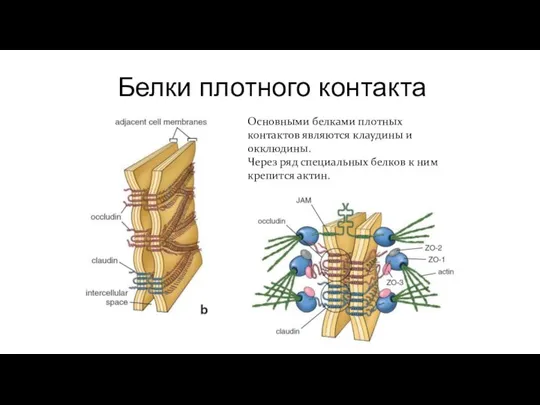
Слайд 95Белки плотного контакта
Основными белками плотных контактов являются клаудины и окклюдины.
Через ряд специальных
Белки плотного контакта
Основными белками плотных контактов являются клаудины и окклюдины.
Через ряд специальных
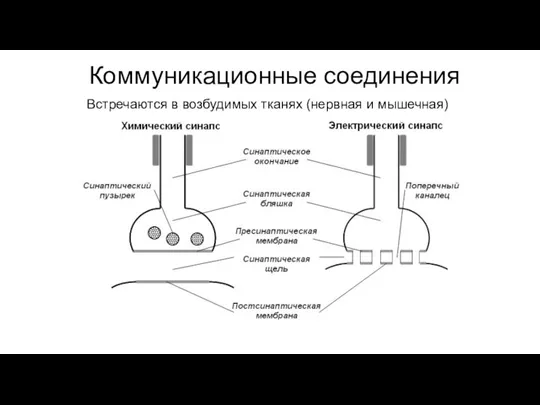
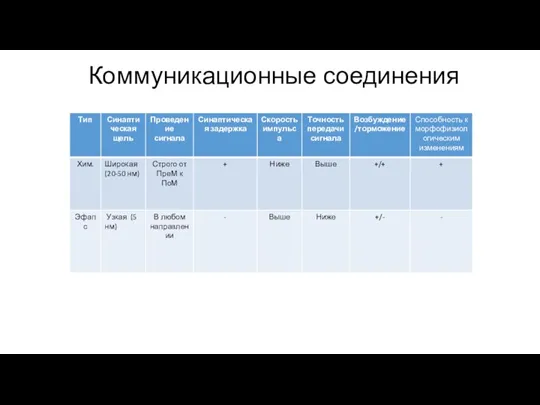
Слайд 96Контакты коммуникационного типа
Щелевидные соединения (нексусы, электрические синапсы, эфапсы)
Нексус имеет форму круга диаметром
Контакты коммуникационного типа
Щелевидные соединения (нексусы, электрические синапсы, эфапсы)
Нексус имеет форму круга диаметром

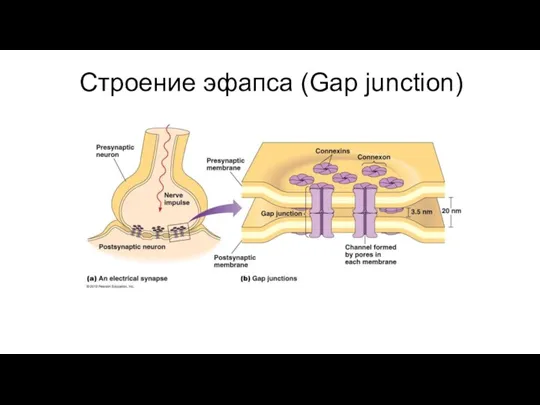
Слайд 97Строение эфапса (Gap junction)
Строение эфапса (Gap junction)
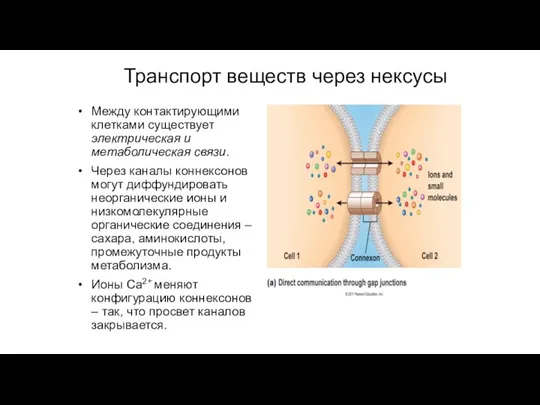
Слайд 98Между контактирующими клетками существует электрическая и метаболическая связи.
Через каналы коннексонов могут
Между контактирующими клетками существует электрическая и метаболическая связи.
Через каналы коннексонов могут
Слайд 99Контакты коммуникационного типа
Синапсы
Синапсы служат для передачи сигнала от одних возбудимых клеток к
Контакты коммуникационного типа
Синапсы
Синапсы служат для передачи сигнала от одних возбудимых клеток к

Слайд 100Коммуникационные соединения
Встречаются в возбудимых тканях (нервная и мышечная)
Коммуникационные соединения
Встречаются в возбудимых тканях (нервная и мышечная)
Слайд 101Коммуникационные соединения
Коммуникационные соединения
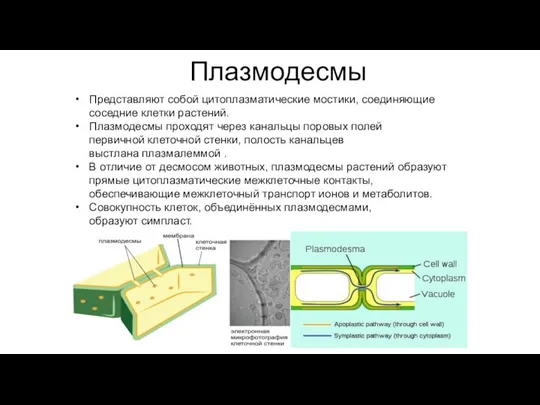
Слайд 102Плазмодесмы
Представляют собой цитоплазматические мостики, соединяющие соседние клетки растений.
Плазмодесмы проходят через канальцы
Плазмодесмы
Представляют собой цитоплазматические мостики, соединяющие соседние клетки растений.
Плазмодесмы проходят через канальцы
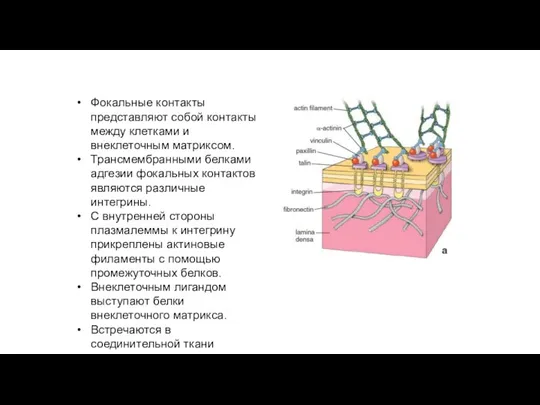
Слайд 103Фокальные контакты клеток
Фокальные контакты представляют собой контакты между клетками и внеклеточным матриксом.
Трансмембранными
Фокальные контакты клеток
Фокальные контакты представляют собой контакты между клетками и внеклеточным матриксом.
Трансмембранными
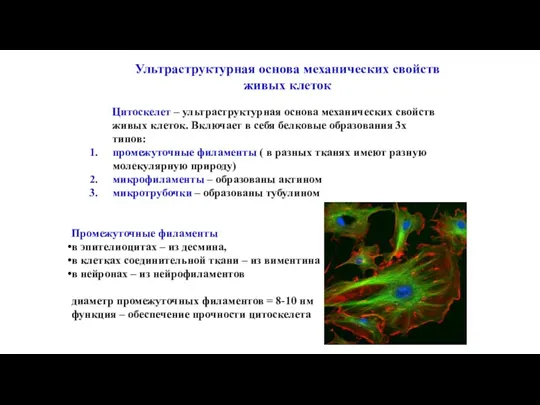
Слайд 104Ультраструктурная основа механических свойств живых клеток
Цитоскелет – ультраструктурная основа механических свойств
Ультраструктурная основа механических свойств живых клеток
Цитоскелет – ультраструктурная основа механических свойств
Слайд 106Саркомер – элементарная надмолекулярная сократительная единица мышечного волокна. Соседние саркомеры соединяются друг
Саркомер – элементарная надмолекулярная сократительная единица мышечного волокна. Соседние саркомеры соединяются друг

Слайд 107Саркомер состоит из тонких (актиновых) и толстых (миозиновых) филаментов, которые называют протофибриллами.
Саркомер состоит из тонких (актиновых) и толстых (миозиновых) филаментов, которые называют протофибриллами.

Слайд 108На поперечном срезе саркомера нити актина и миозина образуют гексагональную структуру, в
На поперечном срезе саркомера нити актина и миозина образуют гексагональную структуру, в

Слайд 109Молекула тропомиозина занимает спиральный желобок между 2-я нитями F- актина, ге расположены
Молекула тропомиозина занимает спиральный желобок между 2-я нитями F- актина, ге расположены

Слайд 110Механизм мышечного сокращения
После разблокирования активного цинтра актина с помощью ионов Ca2+ миозиновые
Механизм мышечного сокращения
После разблокирования активного цинтра актина с помощью ионов Ca2+ миозиновые

Слайд 111Максимальная сила сокращения пропорциональна мышцы пропорциональна взаимного перекрытия актиновых и миозиновых нитей.
Максимальная сила сокращения пропорциональна мышцы пропорциональна взаимного перекрытия актиновых и миозиновых нитей.

Слайд 112При повышении физической нагрузки происходит гипертрофия миоцитов, т.к. это дифференцированные клетки, не
При повышении физической нагрузки происходит гипертрофия миоцитов, т.к. это дифференцированные клетки, не

Слайд 1131 – мышца в покое, 2 – сокращение, 3 - расслабление
При
1 – мышца в покое, 2 – сокращение, 3 - расслабление
При
Слайд 114нервные импульсы, поступающие их спинного
или головного мозга к к нервно-мышечному синапсу
Схема
нервные импульсы, поступающие их спинного
или головного мозга к к нервно-мышечному синапсу
Схема

Слайд 115Метаболический аппарат клетки
Метаболический аппарат клетки

Слайд 116Метаболизм – это совокупность химических реакций, протекающих в живых клетках и обеспечивающих
Метаболизм – это совокупность химических реакций, протекающих в живых клетках и обеспечивающих

Слайд 117Структуры клетки участвующие в метаболизме
Органоиды, участвующие в анаболизме:
ШЭПС
ГлЭПС
Аппарат Гольджи
Рибосомы
Хлоропласты (у растений)
Катаболизм осуществляется
Структуры клетки участвующие в метаболизме
Органоиды, участвующие в анаболизме:
ШЭПС
ГлЭПС
Аппарат Гольджи
Рибосомы
Хлоропласты (у растений)
Катаболизм осуществляется

Слайд 119Эндоплазматический ретиккулум
Эндоплазматический ретикулум (ЭПР) - это замкнутая система мембранных трубочек внутри клетки,
Эндоплазматический ретиккулум
Эндоплазматический ретикулум (ЭПР) - это замкнутая система мембранных трубочек внутри клетки,

Слайд 121Шероховатый ЭПС
На рибосомах ШЭПС синтезируются следующие группы белков:
Белки предназначенные для секреции-выведение через
Шероховатый ЭПС
На рибосомах ШЭПС синтезируются следующие группы белков:
Белки предназначенные для секреции-выведение через

Слайд 122Сборка белка. Этап прикрепления к ШЭПР
Сборка любого белка начинается на рибосомах в
Сборка белка. Этап прикрепления к ШЭПР
Сборка любого белка начинается на рибосомах в

Слайд 123Связывание SRP С SRP-рецептором.
Связывание SRP С SRP-рецептором.
Слайд 124Синтез растворимых белков в ЭПР
Полипептидная цепь (ППЦ) растет в полость канала. Гидрофобный
Синтез растворимых белков в ЭПР
Полипептидная цепь (ППЦ) растет в полость канала. Гидрофобный
Слайд 125Синтез мембранных белков
В синтезируемой ППЦ существует так называемая стоп-последовательность (STS – stop
Синтез мембранных белков
В синтезируемой ППЦ существует так называемая стоп-последовательность (STS – stop

Слайд 129Образование политопных (то есть закрепленных в мембране несколько раз) белков. Процесс формирования
Образование политопных (то есть закрепленных в мембране несколько раз) белков. Процесс формирования
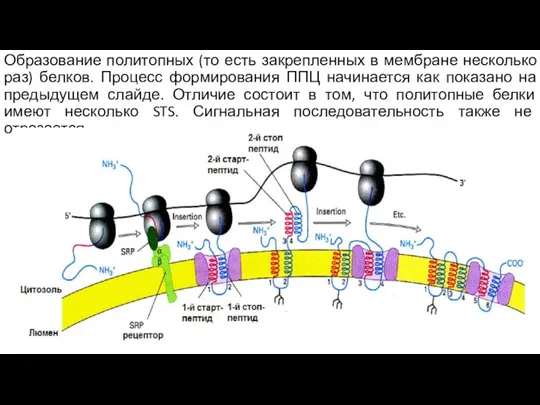
Слайд 130Другие функции ШЭПР
Большинство белков синтезированных в ШЭПР-гликопротеиды. По мере роста белковой цепи
Другие функции ШЭПР
Большинство белков синтезированных в ШЭПР-гликопротеиды. По мере роста белковой цепи

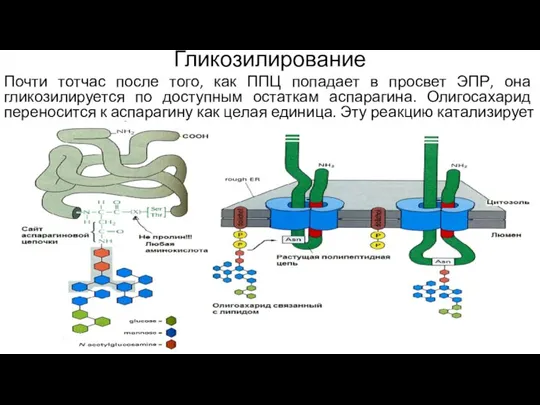
Слайд 131Гликозилирование
Почти тотчас после того, как ППЦ попадает в просвет ЭПР, она гликозилируется
Гликозилирование
Почти тотчас после того, как ППЦ попадает в просвет ЭПР, она гликозилируется
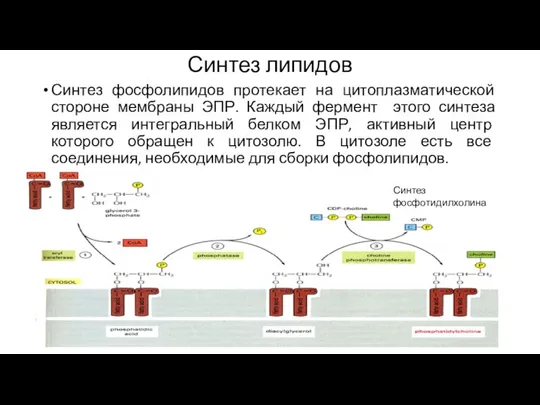
Слайд 132Синтез липидов
Синтез фосфолипидов протекает на цитоплазматической стороне мембраны ЭПР. Каждый фермент этого
Синтез липидов
Синтез фосфолипидов протекает на цитоплазматической стороне мембраны ЭПР. Каждый фермент этого
Слайд 133Гладкий ЭПР
Очень лабилен и способен к перестройкам в зависимости от внешних условий.
Гладкий ЭПР
Очень лабилен и способен к перестройкам в зависимости от внешних условий.

Слайд 134Функции ГлЭПР
Функции ГлЭПР
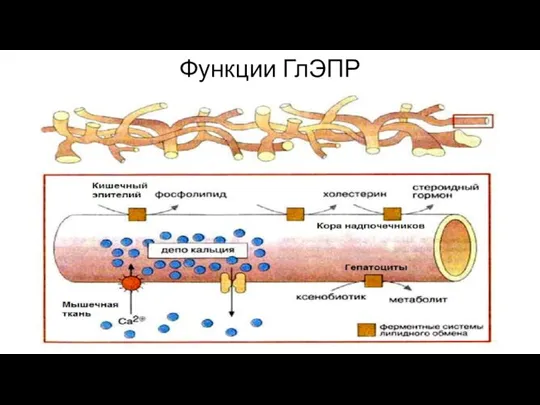
Слайд 135Связь ЭПР с другими компартментами
Связь ЭПР с другими компартментами
Слайд 136Аппарат Гольджи
Аппарат Гольджи- интегрирующая часть метаболической системы эукариотической клетки. В нем происходит
Аппарат Гольджи
Аппарат Гольджи- интегрирующая часть метаболической системы эукариотической клетки. В нем происходит

Слайд 137Tubular compartment
ER-to-Golgi intermediate compartment (ERGIC)
Везикулы из ЭПР не сразу направляются к транс-цистерне,
Tubular compartment
ER-to-Golgi intermediate compartment (ERGIC)
Везикулы из ЭПР не сразу направляются к транс-цистерне,
Слайд 138МОДЕЛЬ ТРАНСПОРТА БЕЛКОВ.
Alberts B. et al «Molecular Biology of the Cell», 2002
МОДЕЛЬ ТРАНСПОРТА БЕЛКОВ.
Alberts B. et al «Molecular Biology of the Cell», 2002
Слайд 139Синтез и модификация веществ
Каждая цистерна аппарата Гольджи имеет свой набор ферментов.
Доработка гликопротеинов;
Создание
Синтез и модификация веществ
Каждая цистерна аппарата Гольджи имеет свой набор ферментов.
Доработка гликопротеинов;
Создание

Слайд 140Сортировка
Сортировка белков происходит в транс-сети аппарата Гольджи. Результатом является разделение белков и
Сортировка
Сортировка белков происходит в транс-сети аппарата Гольджи. Результатом является разделение белков и

Слайд 142Секреция
Вещества выводятся из клетки одним из двух путей: в результате конструктивной (нерегулируемой)
Секреция
Вещества выводятся из клетки одним из двух путей: в результате конструктивной (нерегулируемой)

Слайд 144Функции обобщенно
1.Транспорт - через АГ проходят две группы белков: белки, предназначенные на
Функции обобщенно
1.Транспорт - через АГ проходят две группы белков: белки, предназначенные на

Слайд 145Лизосомы
Лизосомы представляют собой гетерогенную (разнородную) группу цитоплазматических вакуолеподобных структур размером 1-3 мкм,
Лизосомы
Лизосомы представляют собой гетерогенную (разнородную) группу цитоплазматических вакуолеподобных структур размером 1-3 мкм,

Слайд 147В первом случае расщепляемый материал - белки, полинуклеотиды или полисахариды - попадает
В первом случае расщепляемый материал - белки, полинуклеотиды или полисахариды - попадает

Слайд 148Второй путь формирования лизосом называется аутофагией. В процессе аутофагии происходит разрушение отработанных
Второй путь формирования лизосом называется аутофагией. В процессе аутофагии происходит разрушение отработанных

Слайд 150Третий путь формирования лизосом имеется только у клеток, специализированных для фагоцитоза больших
Третий путь формирования лизосом имеется только у клеток, специализированных для фагоцитоза больших

Слайд 151Эндосомы, аутофагосомы и фагосомы часто называют общим термином - прелизосомы, а эндолизосомы,
Эндосомы, аутофагосомы и фагосомы часто называют общим термином - прелизосомы, а эндолизосомы,

Слайд 152Пероксисомы
Мембранные пузырьки с однородным или гранулярным матриксом. В центре-сердцевина (нуклеоид) из кристаллических
Пероксисомы
Мембранные пузырьки с однородным или гранулярным матриксом. В центре-сердцевина (нуклеоид) из кристаллических

Слайд 154Пероксисомы обнаружены во всех эукариотических клетках. Они являются главным центром утилизации кислорода
Пероксисомы обнаружены во всех эукариотических клетках. Они являются главным центром утилизации кислорода

Слайд 155Митохондрии
Митохондрии, как органеллы синтеза АТФ характерны для всех эукариотических клеток как автотрофных
Митохондрии
Митохондрии, как органеллы синтеза АТФ характерны для всех эукариотических клеток как автотрофных

Слайд 156Локализация
Митохондрии скапливаются в энергозависимых участках клетки. В скелетных мышцах они располагаются между
Локализация
Митохондрии скапливаются в энергозависимых участках клетки. В скелетных мышцах они располагаются между

Слайд 157Строение
Имеющиеся методы позволяют выделить из митохондрий все четыре компонента: наружную мембрану,содержимое межмембранного
Строение
Имеющиеся методы позволяют выделить из митохондрий все четыре компонента: наружную мембрану,содержимое межмембранного

Слайд 160Наружная мембрана.
В состав наружной мембраны входит много молекул белка порина. Особенность его
Наружная мембрана.
В состав наружной мембраны входит много молекул белка порина. Особенность его

Слайд 161Межмембранное пространство
Состав вещества межмембранного пространства близок к цитозолю.
Одним из белков, содержащихся в межмембранном пространстве, является цитохром c один из компонентов дыхатеьной цепи митохондрий.
Межмембранное пространство
Состав вещества межмембранного пространства близок к цитозолю.
Одним из белков, содержащихся в межмембранном пространстве, является цитохром c один из компонентов дыхатеьной цепи митохондрий.
Слайд 162Внутренняя мембрана
Внутренняя мембрана ограничивает основное рабочее пространство митохондрии. Она высокоспецифична, содержит большое
Внутренняя мембрана
Внутренняя мембрана ограничивает основное рабочее пространство митохондрии. Она высокоспецифична, содержит большое

Слайд 163Матрикс
Матрикс содержит высококонцентрированную смесь сотен различных ферментов, необходимых для окисления пирувата, жирных
Матрикс
Матрикс содержит высококонцентрированную смесь сотен различных ферментов, необходимых для окисления пирувата, жирных

Слайд 164Генетический аппарат
Кольцевая молекула ДНК. Редко-линейная;
Может синтезировать белок как с метохондриального иРНК, так
Генетический аппарат
Кольцевая молекула ДНК. Редко-линейная;
Может синтезировать белок как с метохондриального иРНК, так

Слайд 165Функции
Синтез АТФ;
Специфический синтез-стероидные гормоны, некоторые липиды;
Накопление ионов, особенно Ca2+;
Накопление продуктов экскреции в
Функции
Синтез АТФ;
Специфический синтез-стероидные гормоны, некоторые липиды;
Накопление ионов, особенно Ca2+;
Накопление продуктов экскреции в

Слайд 166Этапы
энергетического обмена:
1. Подготовительный
2. Гликолиз
(бескислородное окисление)
3. Дыхание
(кислородное окисление)
Этапы
энергетического обмена:
1. Подготовительный
2. Гликолиз
(бескислородное окисление)
3. Дыхание
(кислородное окисление)

Слайд 167 Первый этап
Подготовительный
ферментативное расщепление сложных органических веществ
до простых
Первый этап
Подготовительный
ферментативное расщепление сложных органических веществ
до простых

Слайд 168Где происходит:
Пищеварительная система
Лизосомы в клетках
Где происходит:
Пищеварительная система
Лизосомы в клетках
Слайд 169Субстрат
Углеводы = глюкоза + Е (1г = 17,6 кДж)
Липиды = глицерин
Субстрат
Углеводы = глюкоза + Е (1г = 17,6 кДж)
Липиды = глицерин

Слайд 170Результат этапа
Энергия не запасается, а выделяется только в тепловой форме
Результат этапа
Энергия не запасается, а выделяется только в тепловой форме

Слайд 171Гликолиз
- неполное расщепление
- анаэробное дыхание
- брожение
Второй этап
Бескислородное окисление
Гликолиз
- неполное расщепление
- анаэробное дыхание
- брожение
Второй этап
Бескислородное окисление

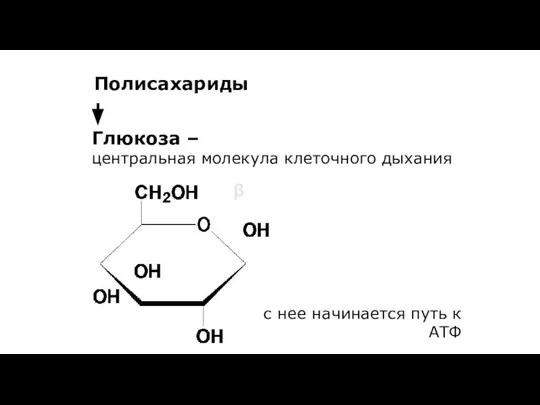
Слайд 172Глюкоза –
центральная молекула клеточного дыхания
Полисахариды
β
с нее начинается путь к АТФ
Глюкоза –
центральная молекула клеточного дыхания
Полисахариды
β
с нее начинается путь к АТФ
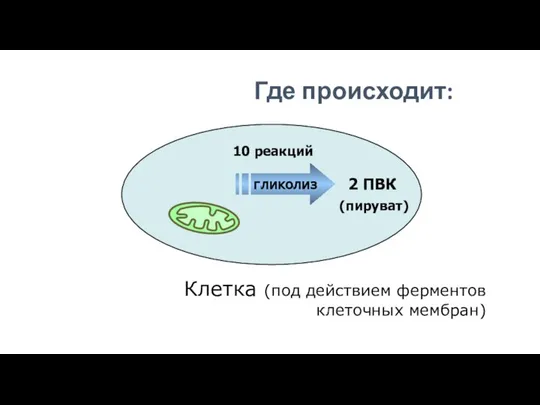
Слайд 1732 ПВК
Клетка (под действием ферментов клеточных мембран)
10 реакций
(пируват)
гликолиз
Где происходит:
2 ПВК
Клетка (под действием ферментов клеточных мембран)
10 реакций
(пируват)
гликолиз
Где происходит:
Слайд 174Субстрат
С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ →
глюкоза
2С3Н4О3 + 2АТФ
Субстрат
С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ →
глюкоза
2С3Н4О3 + 2АТФ

Слайд 17560% выделяется в виде тепла
40%
идет на синтез
АТФ
Энергия
Результат этапа:
из одной молекулы
60% выделяется в виде тепла
40%
идет на синтез
АТФ
Энергия
Результат этапа: из одной молекулы

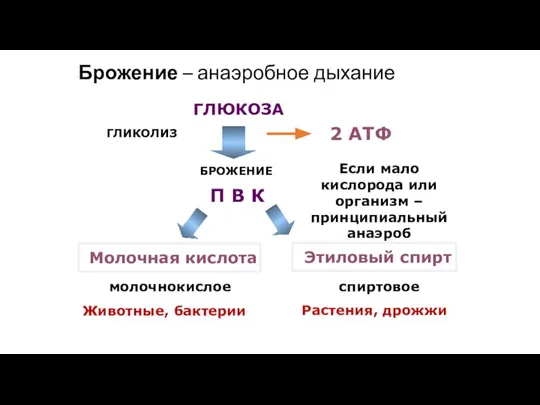
Слайд 176ГЛЮКОЗА
П В К
2 АТФ
Брожение – анаэробное дыхание
Если мало кислорода или организм –
ГЛЮКОЗА
П В К
2 АТФ
Брожение – анаэробное дыхание
Если мало кислорода или организм –
Слайд 177Выводы:
Синтез АТФ в процессе гликолиза не нуждается в мембранах. Он идёт
Выводы:
Синтез АТФ в процессе гликолиза не нуждается в мембранах. Он идёт

Слайд 178Третий этап
Кислородное расщепление:
полное расщепление пировиноградной кислоты, происходит при обязательном присутствии кислорода
Третий этап
Кислородное расщепление:
полное расщепление пировиноградной кислоты, происходит при обязательном присутствии кислорода

Слайд 179О2
Митохондрия: под действием ферментов митохондриальных мембран (необходимое условие – целостность мембран)
ПВК
СО2 и
О2
Митохондрия: под действием ферментов митохондриальных мембран (необходимое условие – целостность мембран)
ПВК
СО2 и
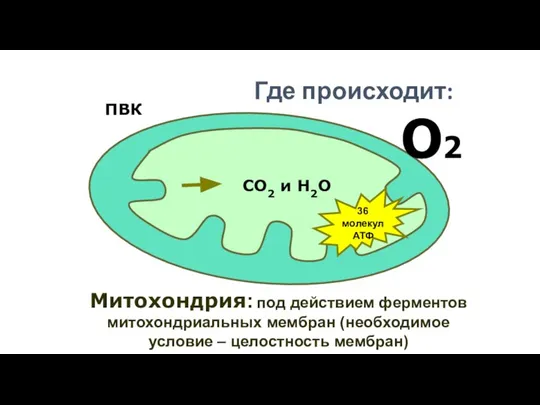
Слайд 180Стадии аэробного дыхания:
1) Окислительное декарбоксилирование
2) Цикл Кребса
3) Электронтранспортная цепь (окислительное фосфолирирование)
Стадии аэробного дыхания:
1) Окислительное декарбоксилирование
2) Цикл Кребса
3) Электронтранспортная цепь (окислительное фосфолирирование)

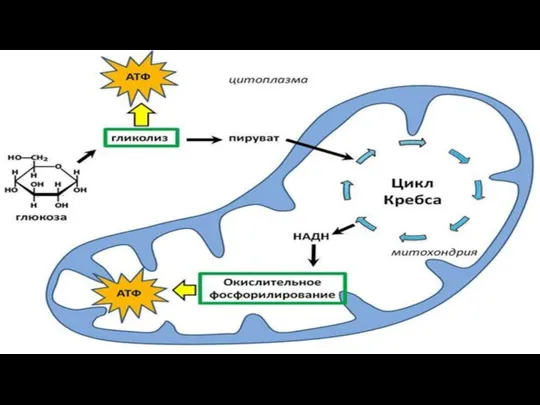
Слайд 182Электронтранспортная цепь
Электронтранспортная цепь
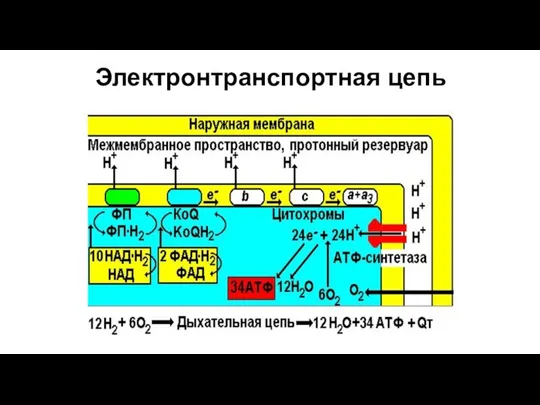
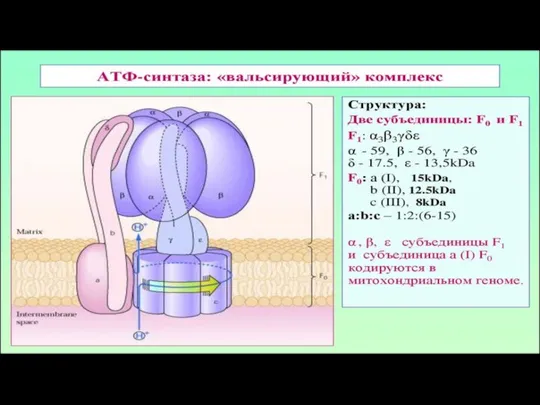
Слайд 183АТФ-ситаза
Принимает электроны от цитохрома с и передает их на кислород с образованием
АТФ-ситаза
Принимает электроны от цитохрома с и передает их на кислород с образованием

Слайд 185Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из
Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из

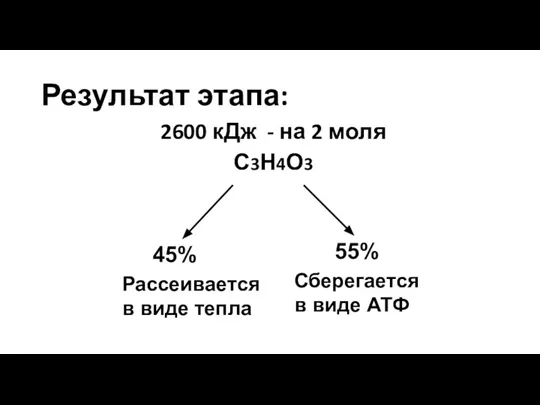
Слайд 186Результат этапа:
2600 кДж - на 2 моля
С3Н4О3
45%
Рассеивается
в виде тепла
Сберегается
в виде
Результат этапа:
2600 кДж - на 2 моля
С3Н4О3
45%
Рассеивается
в виде тепла
Сберегается
в виде
Слайд 187Выводы:
Для осуществления кислородного процесса необходимо наличие неповреждённых митохондриальных мембран
Выводы:
Для осуществления кислородного процесса необходимо наличие неповреждённых митохондриальных мембран

Слайд 188Выводы:
Расщепление в клетке
1 молекулы глюкозы до СО2
и Н2О обеспечивает синтез
Выводы:
Расщепление в клетке
1 молекулы глюкозы до СО2
и Н2О обеспечивает синтез

Слайд 189Ядерный аппарат
Ядерный аппарат

Слайд 190Состав ядерного аппарата:
Поверхностный аппарат ядра;
Ядерный матрикс;
Кариоплазма;
Хроматин;
Ядрышко.
Состав ядерного аппарата:
Поверхностный аппарат ядра;
Ядерный матрикс;
Кариоплазма;
Хроматин;
Ядрышко.

Слайд 1911 — ядерная оболочка (две мембраны - внутренняя и внешняя, и перинуклеарное
1 — ядерная оболочка (две мембраны - внутренняя и внешняя, и перинуклеарное

Слайд 192Поверхностный аппарат
Ядерная оболочка с порами:
Разделена на внешнюю и внутреннюю ядерные оболочки. Пространство
Поверхностный аппарат
Ядерная оболочка с порами:
Разделена на внешнюю и внутреннюю ядерные оболочки. Пространство

Слайд 193Поровый комплекс
Поровый комплекс
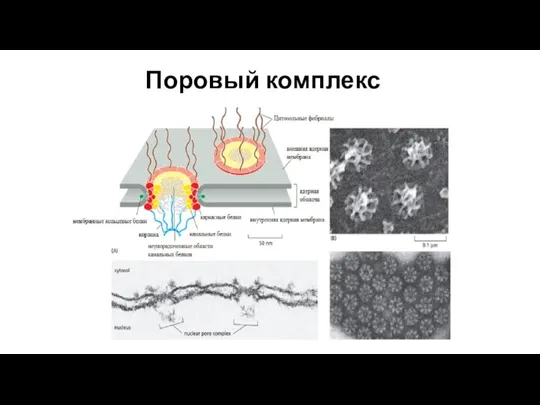
Слайд 194РНК должны попасть из ядра в цитоплазму. Для этого нужно преодолеть ядерную
РНК должны попасть из ядра в цитоплазму. Для этого нужно преодолеть ядерную

Слайд 195Функции поверхностного аппарата
Отграничение ядра от цитоплазмы. В результате этого происходит разделение транскрипции
Функции поверхностного аппарата
Отграничение ядра от цитоплазмы. В результате этого происходит разделение транскрипции

Слайд 196Ядерный матрикс
Ядерный матрикс-это скелет ядра. Белки связывают определенные последовательности ДНК, образующие основание
Ядерный матрикс
Ядерный матрикс-это скелет ядра. Белки связывают определенные последовательности ДНК, образующие основание

Слайд 197Ядрышко
Ядрышко – высокоорганизованная шаровидная структура ядра. Содержит большие петли ДНК, выступающие из
Ядрышко
Ядрышко – высокоорганизованная шаровидная структура ядра. Содержит большие петли ДНК, выступающие из

Слайд 198Хроматин
Хроматин: основными компонентами являютсяДНК,гистоны инегистоновые белки (ДНП). Соотношение ДНК и белка составляет
Хроматин
Хроматин: основными компонентами являютсяДНК,гистоны инегистоновые белки (ДНП). Соотношение ДНК и белка составляет

Слайд 199Урони упаковки хроматина
Урони упаковки хроматина

Слайд 200Урони упаковки хроматина
Нуклеосомный уровень-8 молекул гистонов закручивают фрагмент 140 пар оснований. Линкерный
Урони упаковки хроматина
Нуклеосомный уровень-8 молекул гистонов закручивают фрагмент 140 пар оснований. Линкерный


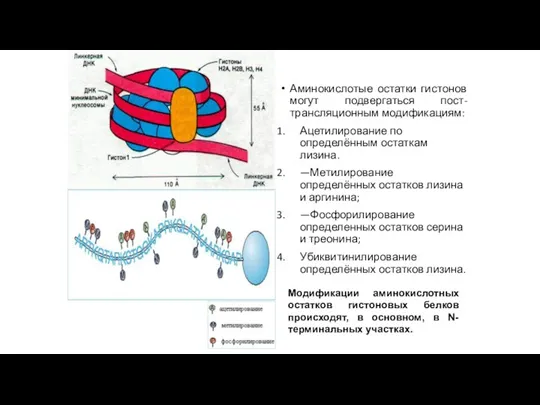
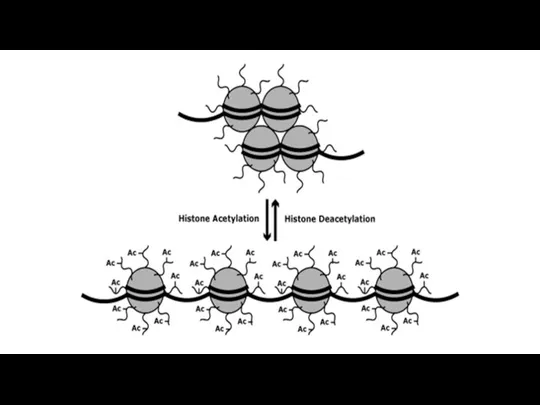
Слайд 202Аминокислотые остатки гистонов могут подвергаться пост-трансляционным модификациям:
Ацетилирование по определённым остаткам лизина.
—Метилирование определённых
Аминокислотые остатки гистонов могут подвергаться пост-трансляционным модификациям:
Ацетилирование по определённым остаткам лизина.
—Метилирование определённых
Слайд 204Метилирование ДНК
Метилирование ДНК
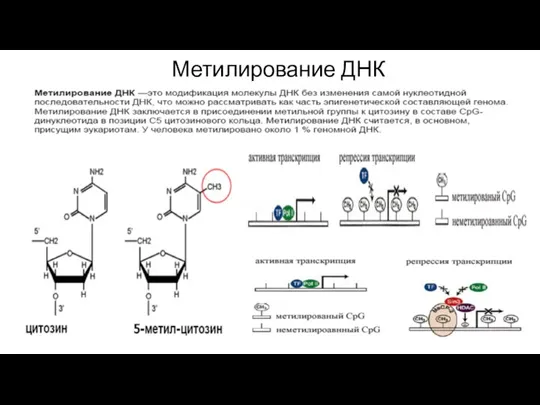

Слайд 206Что происходит?
Метильные группы нарушают ДНК-белковые взаимодействия, выступая в большую бороздку ДНК и
Что происходит?
Метильные группы нарушают ДНК-белковые взаимодействия, выступая в большую бороздку ДНК и

Слайд 207Архитектурные белки высокомобильной группы (HMG)
Были названы так потому, что обладают высокой электрофоретической
Архитектурные белки высокомобильной группы (HMG)
Были названы так потому, что обладают высокой электрофоретической

Слайд 208Хроматин-перестраивающие комплексы
В середине 90-хгодов стало известно, что хроматин содержит комплексы, перестраивающие нуклеосомы
Хроматин-перестраивающие комплексы
В середине 90-хгодов стало известно, что хроматин содержит комплексы, перестраивающие нуклеосомы

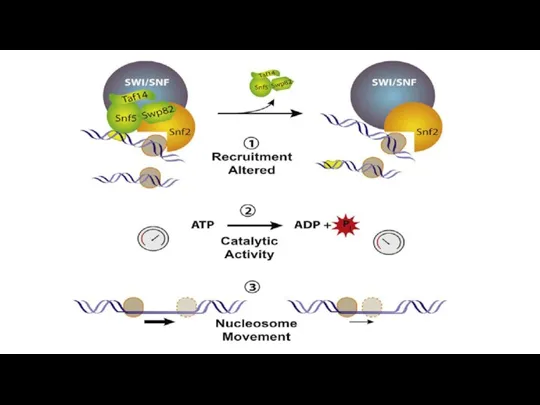
Слайд 210Функция белков хроматина
Определяют уровень компактизации ДНК;
Влияют на уровень транскрипции;
Осуществляют процесс транскрипции.
Функция белков хроматина
Определяют уровень компактизации ДНК;
Влияют на уровень транскрипции;
Осуществляют процесс транскрипции.

Слайд 211Функции ядра
Хранение и передача генетической информации.
Реализация генетической информации.
Функции ядра
Хранение и передача генетической информации.
Реализация генетической информации.

Слайд 212Основы молекулярной биологии клетки
Основы молекулярной биологии клетки

Слайд 213Что такое молекулярная биология?
Жизнь — способ существования биополимеров, из которых основными являются
Что такое молекулярная биология?
Жизнь — способ существования биополимеров, из которых основными являются

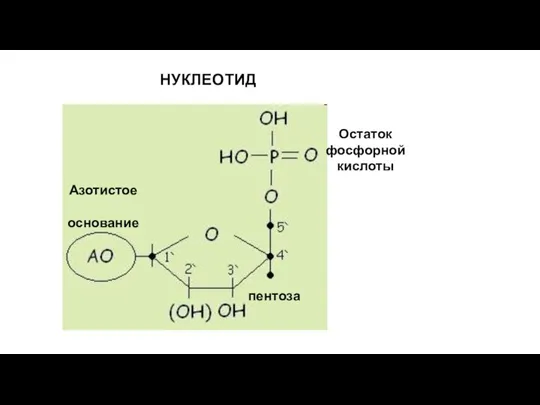
Слайд 214НУКЛЕОТИД
Остаток
фосфорной
кислоты
пентоза
Азотистое
основание
НУКЛЕОТИД
Остаток
фосфорной
кислоты
пентоза
Азотистое
основание
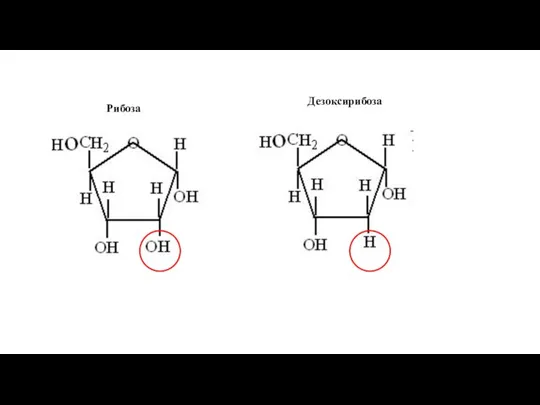
Слайд 215Рибоза
Дезоксирибоза
Рибоза
Дезоксирибоза
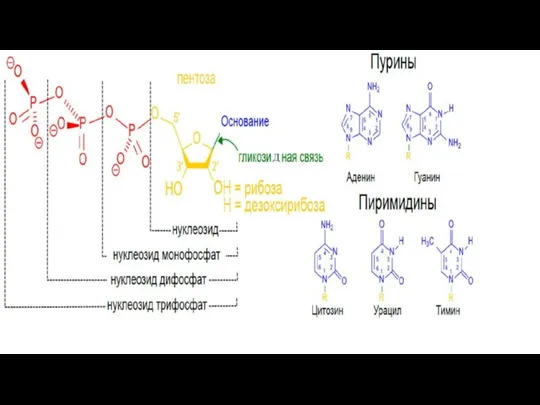
Слайд 216Нуклеиновые кислоты
Нуклеиновые кислоты выполняют генетическую функцию (ДНК) и участвуют в реализации генетической информации
Нуклеиновые кислоты
Нуклеиновые кислоты выполняют генетическую функцию (ДНК) и участвуют в реализации генетической информации

Слайд 218Связи в нуклеотидах
Гликозидная связь— это тип ковалентной связи, которая соединяет молекулу сахара
Связи в нуклеотидах
Гликозидная связь— это тип ковалентной связи, которая соединяет молекулу сахара

Слайд 219Производные нуклиотидов
Нуклиотиды, помимо того,что являются мономерами ДНК и РНК, учавствуют в большом
Производные нуклиотидов
Нуклиотиды, помимо того,что являются мономерами ДНК и РНК, учавствуют в большом

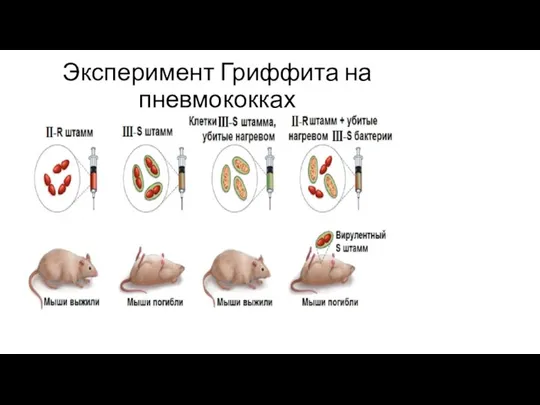
Слайд 220Эксперимент Гриффита на пневмококках
Эксперимент Гриффита на пневмококках
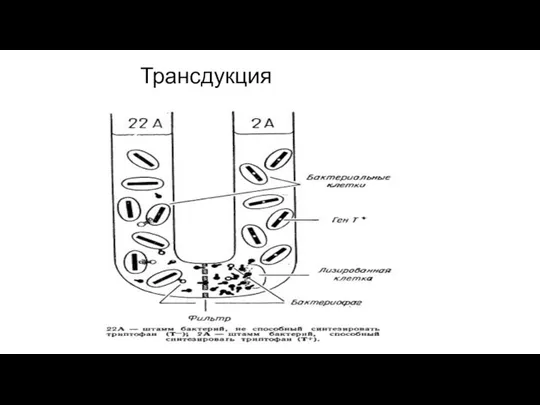
Слайд 221Трансдукция
Трансдукция
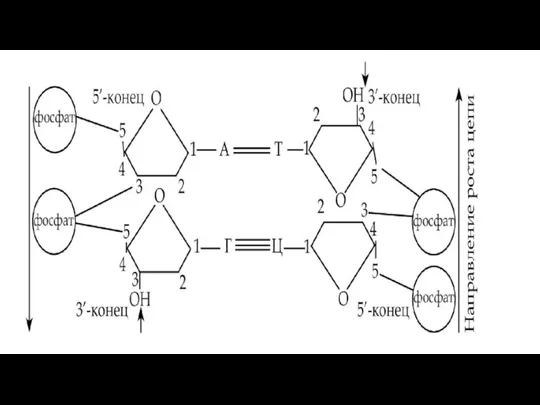
Слайд 222Строение ДНК
Нуклеотиды одной цепи соединяются связями, возникающими между остатком фосфорной кислоты и
Строение ДНК
Нуклеотиды одной цепи соединяются связями, возникающими между остатком фосфорной кислоты и

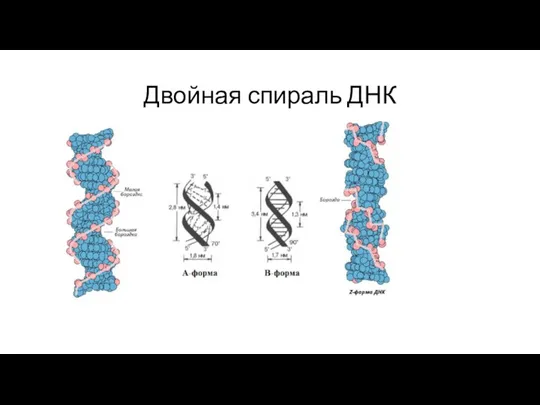
Слайд 224Двойная спираль ДНК
Двойная спираль ДНК
Слайд 225Комплементарность и правила Чаргаффа
Комплементарность - это принцип взаимного соответствия парных нуклеотидов или
Комплементарность и правила Чаргаффа
Комплементарность - это принцип взаимного соответствия парных нуклеотидов или

Слайд 226Функции ДНК
Функции ДНК

Слайд 227РНК
Молекула РНК представляет собой неразветвленный полинуклеотид, который может иметь первичную структуру –
РНК
Молекула РНК представляет собой неразветвленный полинуклеотид, который может иметь первичную структуру –

Слайд 228Виды РНК
Существует три основных класса рибонуклеиновых кислот:
Информационные (матричные) РНК — иРНК
Виды РНК
Существует три основных класса рибонуклеиновых кислот:
Информационные (матричные) РНК — иРНК
Слайд 229Информационная РНК.
Наиболее разнообразный по размерам и стабильности класс. Все они являются
Информационная РНК.
Наиболее разнообразный по размерам и стабильности класс. Все они являются

Слайд 230тРНК
Молекулы тРНК— сравнительно короткие молекулы, состоящие из 71–94 нуклеотидов.
тРНК приходится до
тРНК
Молекулы тРНК— сравнительно короткие молекулы, состоящие из 71–94 нуклеотидов.
тРНК приходится до

Слайд 231В состав тРНК входят несколько необычных азотистых оснований (минорных): инозин (Y), псевдоуридин
В состав тРНК входят несколько необычных азотистых оснований (минорных): инозин (Y), псевдоуридин
Слайд 232Рибосомы
В состав субъединиц рибосом входят разнообразные рРНК, различающиеся по константе седиментации (единица
Рибосомы
В состав субъединиц рибосом входят разнообразные рРНК, различающиеся по константе седиментации (единица

Слайд 234Репликация
Самовоспроизведение ДНК (процесс, приводящий к удвоению молекул ДНК), называется репликацией.
Репликация необходима
Репликация
Самовоспроизведение ДНК (процесс, приводящий к удвоению молекул ДНК), называется репликацией.
Репликация необходима

Слайд 235Участок молекулы ДНК, на котором идет репликация, называется репликон. У прокариот репликон

Слайд 236Начинается процесс с разрыва водородных связей между азотистыми основаниями ДНК на участке,
Начинается процесс с разрыва водородных связей между азотистыми основаниями ДНК на участке,

Слайд 237На другой половине репликона, где разъединение цепей ДНК идёт в другую сторону,
На другой половине репликона, где разъединение цепей ДНК идёт в другую сторону,

Слайд 239Повреждения ДНК
Спонтанные повреждения ДНК:
Ошибки репликации (появление некомплементарных пар нуклеотидов);
Апуринизация (отщепление азотистых оснований
Повреждения ДНК
Спонтанные повреждения ДНК:
Ошибки репликации (появление некомплементарных пар нуклеотидов);
Апуринизация (отщепление азотистых оснований

Слайд 240Фоторепарация
1. Нормальная молекула ДНК
↓
Облучение УФ-светом
↓
2. Мутантная молекула
Фоторепарация
1. Нормальная молекула ДНК
↓
Облучение УФ-светом
↓
2. Мутантная молекула


Слайд 242Эксцизионная репарация
1. Узнавание повреждения ДНК эндонуклеазой;
2. Инцизия (надрезание) цепи ДНК ферментом по
Эксцизионная репарация
1. Узнавание повреждения ДНК эндонуклеазой;
2. Инцизия (надрезание) цепи ДНК ферментом по

Слайд 243Пострепликативная репарация ДНК
Пострепликативная репарация осуществляется в тех случаях, когда повреждение доживает до
Пострепликативная репарация ДНК
Пострепликативная репарация осуществляется в тех случаях, когда повреждение доживает до

 Генетический код
Генетический код Онтогенез, жизненные формы
Онтогенез, жизненные формы Что это за листья?
Что это за листья? Опорно-двигательный аппарат человека
Опорно-двигательный аппарат человека Презентация на тему "Социальное партнерство в профориентационном воспитании при изучении биологии" - презентации по Биологи
Презентация на тему "Социальное партнерство в профориентационном воспитании при изучении биологии" - презентации по Биологи 9klass
9klass Распознавание на таблицах органов и систем органов 8 класс
Распознавание на таблицах органов и систем органов 8 класс Красная книга Красноярского края
Красная книга Красноярского края Птицы
Птицы Сингониум. Размножение
Сингониум. Размножение Красная книга
Красная книга Сон на службе у здоровья
Сон на службе у здоровья Гуморальная регуляция
Гуморальная регуляция Класс Птицы
Класс Птицы Гормоны. Номенклатура гормонов
Гормоны. Номенклатура гормонов Правила заделки семян в почву
Правила заделки семян в почву MOLEKUlyarnaya_biologia_LPZ_1
MOLEKUlyarnaya_biologia_LPZ_1 Обмен белков
Обмен белков Деление. Мейоз. Гаметогенез. 2022
Деление. Мейоз. Гаметогенез. 2022 Класс Насекомые (размножение)
Класс Насекомые (размножение) 8 советов, как сохранить срезанные цветы
8 советов, как сохранить срезанные цветы Phylogenetic disorders of skeletal system
Phylogenetic disorders of skeletal system Органы растений
Органы растений Конкурсное задание Агрономия
Конкурсное задание Агрономия Нуклеиновые кислоты
Нуклеиновые кислоты Брюхоногие моллюски
Брюхоногие моллюски Бактерии
Бактерии Гормоны периферических желез внутренней секреции
Гормоны периферических желез внутренней секреции