- Физиология миокарда

Содержание
- 2. 1. УЛЬТРАСТРУКТУРНАЯ ОРГАНИЗАЦИЯ МИОКАКРДА В отличие скелетной мускулатуры мышечные волокна сердца ветвятся и контактируют между собой
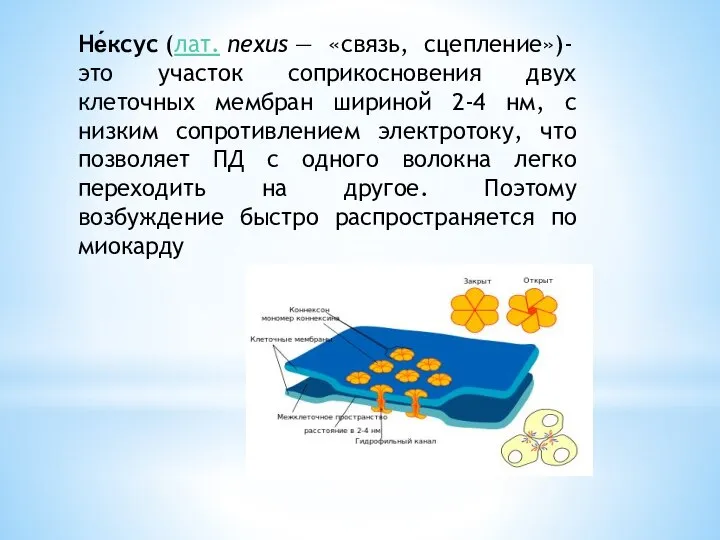
- 3. Не́ксус (лат. nexus — «связь, сцепление»)- это участок соприкосновения двух клеточных мембран шириной 2-4 нм, с
- 4. Функциональный синцитий Благодаря нексусам мышечная стенка сердца функционально ведет себя как единая большая клетка. Возбуждение по
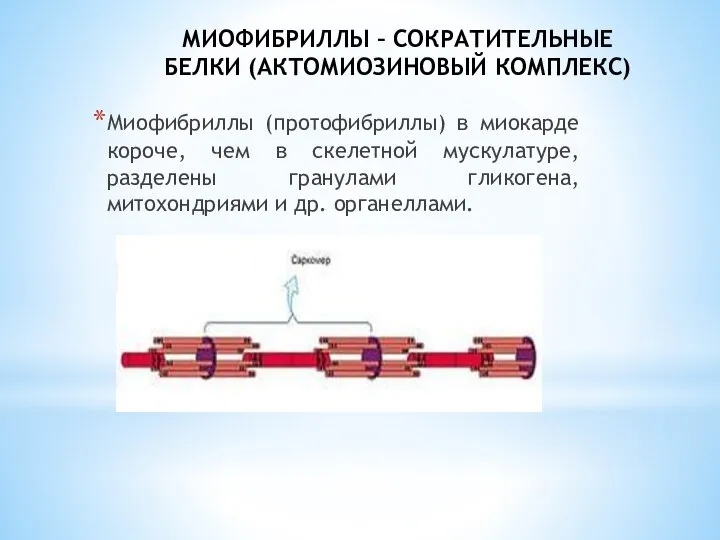
- 5. МИОФИБРИЛЛЫ – СОКРАТИТЕЛЬНЫЕ БЕЛКИ (АКТОМИОЗИНОВЫЙ КОМПЛЕКС) Миофибриллы (протофибриллы) в миокарде короче, чем в скелетной мускулатуре, разделены
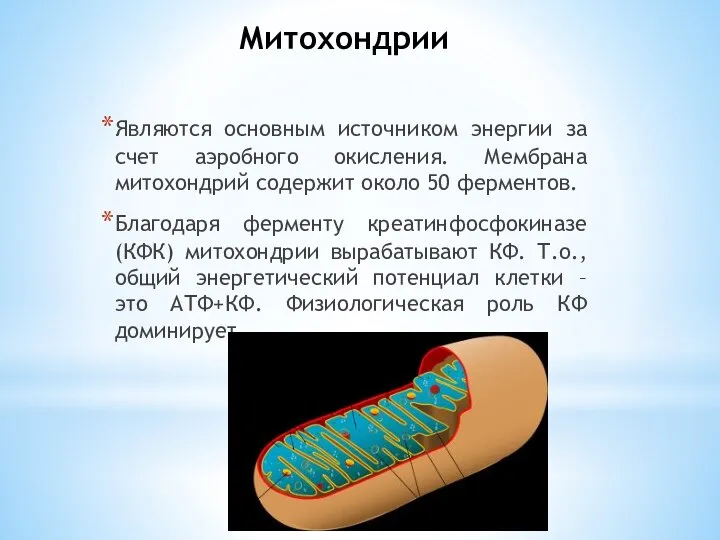
- 6. Митохондрии Являются основным источником энергии за счет аэробного окисления. Мембрана митохондрий содержит около 50 ферментов. Благодаря
- 7. ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ ВОЗБУДИМОСТЬ ПРОВОДИМОСТЬ СОКРАТИМОСТЬ АВТОМАТИЯ
- 8. ВОЗБУДИМОСТЬ Это способность генерировать ПД, если величина раздражителя является пороговой или превышает порог. Длительность ПД при
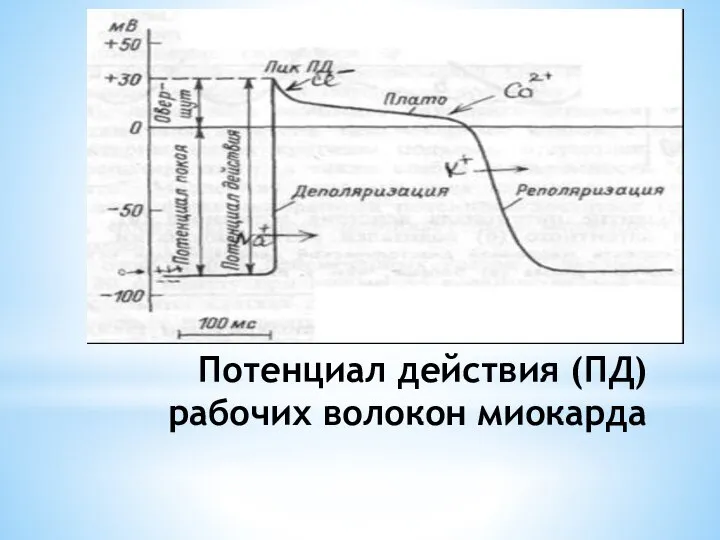
- 9. Потенциал действия (ПД) рабочих волокон миокарда
- 10. ПД рабочих волокон миокарда и атипичных миоцитов проводящей системы сердца
- 11. Фазы ПД рабочих волокон миокарда и атипичных клеток сердца 0. Фаза быстрой деполяризации 1. Фаза быстрой
- 12. ОВЕРШУТ Во время 0-фазы на пике ПД мембрана перезаряжается с – 90 мВ до +30 мВ.
- 13. Фаза «плато»( II) Замедление реполяризации имеет глубокий смысл. Дело в том, что во время этой фазы
- 14. Фаза быстрой конечной реполяризации Эта III- я фаза ПД полностью восстанавливает уровень МП до уровня примерно
- 15. ПД атипичных миоцитов проводящей системы сердца
- 16. Функции проводящей системы сердца Проводящая система выполняет две функции: во-первых, она является внутрисердечным генератором ритма сердца,
- 17. Проводящая система сердца
- 18. Элементы проводящей системы Синоатриальный узел (САУ) Кисса-Флека (лат. nódius sinuatriális) расположен субэндокардиально в стенке правого предсердия
- 19. Градиент автоматизма САУ является водителем нормального ритма сердца (60-80 в мин). Он называется водителем ритма сердца
- 20. Скорость проведения импульсов возбуждения
- 21. В различных участках сердца скорость проведения возбуждения неодинакова. В синоатриальном узле скорость проведения возбуждения невелика -
- 22. СОКРАТИМОСТЬ Амплитуда одиночных сокращений миокарда зависит от частоты сокращений и величины растяжения волокон миокарда во время
- 23. Лестница Боудича Это закон гомеометрической саморегуляции клеток сердца (хроно-инотропная зависимость). Только во второй половине XX века
- 24. Гетерометрическая саморегуляция сокращений (закон Франка-Старлинга) Это зависимость силы сокращений от величины растяжения мышечных волокон. Чем больше
- 26. Скачать презентацию
Слайд 21. УЛЬТРАСТРУКТУРНАЯ ОРГАНИЗАЦИЯ МИОКАКРДА
В отличие скелетной мускулатуры мышечные волокна сердца ветвятся и
1. УЛЬТРАСТРУКТУРНАЯ ОРГАНИЗАЦИЯ МИОКАКРДА
В отличие скелетной мускулатуры мышечные волокна сердца ветвятся и

Слайд 3Не́ксус (лат. nexus — «связь, сцепление»)- это участок соприкосновения двух клеточных мембран шириной 2-4 нм,
Не́ксус (лат. nexus — «связь, сцепление»)- это участок соприкосновения двух клеточных мембран шириной 2-4 нм,
Слайд 4Функциональный синцитий
Благодаря нексусам мышечная стенка сердца функционально ведет себя как единая большая
Функциональный синцитий
Благодаря нексусам мышечная стенка сердца функционально ведет себя как единая большая

Слайд 5МИОФИБРИЛЛЫ – СОКРАТИТЕЛЬНЫЕ БЕЛКИ (АКТОМИОЗИНОВЫЙ КОМПЛЕКС)
Миофибриллы (протофибриллы) в миокарде короче, чем в
МИОФИБРИЛЛЫ – СОКРАТИТЕЛЬНЫЕ БЕЛКИ (АКТОМИОЗИНОВЫЙ КОМПЛЕКС)
Миофибриллы (протофибриллы) в миокарде короче, чем в
Слайд 6Митохондрии
Являются основным источником энергии за счет аэробного окисления. Мембрана митохондрий содержит около
Митохондрии
Являются основным источником энергии за счет аэробного окисления. Мембрана митохондрий содержит около
Слайд 7ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ
ВОЗБУДИМОСТЬ
ПРОВОДИМОСТЬ
СОКРАТИМОСТЬ
АВТОМАТИЯ
ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ
ВОЗБУДИМОСТЬ
ПРОВОДИМОСТЬ
СОКРАТИМОСТЬ
АВТОМАТИЯ

Слайд 8ВОЗБУДИМОСТЬ
Это способность генерировать ПД, если величина раздражителя является пороговой или превышает порог.
ВОЗБУДИМОСТЬ
Это способность генерировать ПД, если величина раздражителя является пороговой или превышает порог.

Слайд 9Потенциал действия (ПД) рабочих волокон миокарда
Потенциал действия (ПД) рабочих волокон миокарда
Слайд 10ПД рабочих волокон миокарда и атипичных миоцитов проводящей системы сердца
ПД рабочих волокон миокарда и атипичных миоцитов проводящей системы сердца
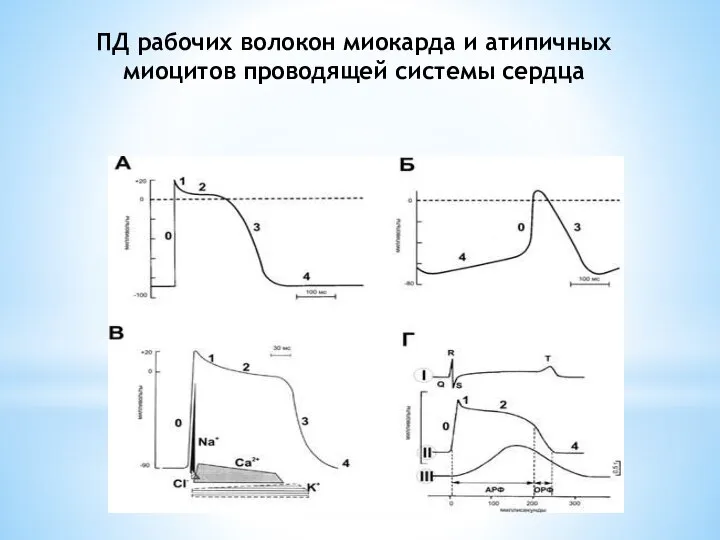
Слайд 11Фазы ПД рабочих волокон миокарда и атипичных клеток сердца
0. Фаза быстрой деполяризации
1.
Фазы ПД рабочих волокон миокарда и атипичных клеток сердца
0. Фаза быстрой деполяризации
1.

Слайд 12ОВЕРШУТ
Во время 0-фазы на пике ПД мембрана перезаряжается с – 90
ОВЕРШУТ
Во время 0-фазы на пике ПД мембрана перезаряжается с – 90

Слайд 13Фаза «плато»( II)
Замедление реполяризации имеет глубокий смысл. Дело в том, что во
Фаза «плато»( II)
Замедление реполяризации имеет глубокий смысл. Дело в том, что во

Слайд 14Фаза быстрой конечной реполяризации
Эта III- я фаза ПД полностью восстанавливает уровень
Фаза быстрой конечной реполяризации
Эта III- я фаза ПД полностью восстанавливает уровень

Слайд 15ПД атипичных миоцитов проводящей системы сердца
ПД атипичных миоцитов проводящей системы сердца
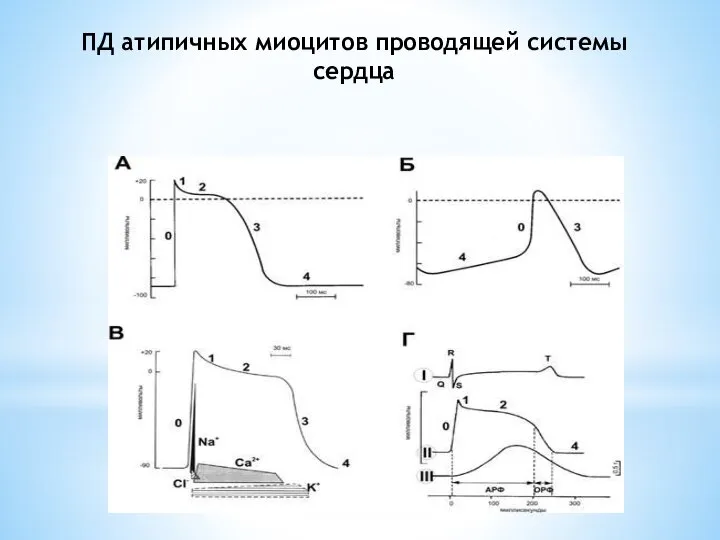
Слайд 16Функции проводящей системы сердца
Проводящая система выполняет две функции: во-первых, она является внутрисердечным
Функции проводящей системы сердца
Проводящая система выполняет две функции: во-первых, она является внутрисердечным

Слайд 17Проводящая система сердца
Проводящая система сердца
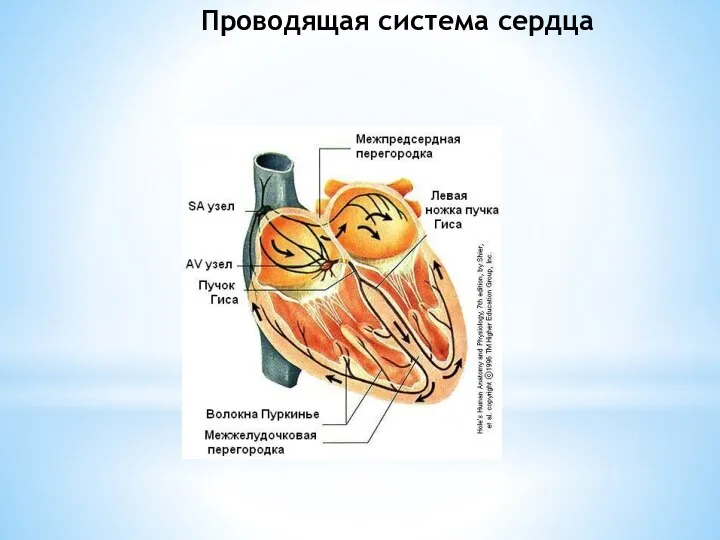
Слайд 18Элементы проводящей системы
Синоатриальный узел (САУ) Кисса-Флека (лат. nódius sinuatriális) расположен субэндокардиально в стенке
Элементы проводящей системы
Синоатриальный узел (САУ) Кисса-Флека (лат. nódius sinuatriális) расположен субэндокардиально в стенке

Слайд 19Градиент автоматизма
САУ является водителем нормального ритма сердца (60-80 в мин). Он называется
Градиент автоматизма
САУ является водителем нормального ритма сердца (60-80 в мин). Он называется

Слайд 20Скорость проведения импульсов возбуждения
Скорость проведения импульсов возбуждения
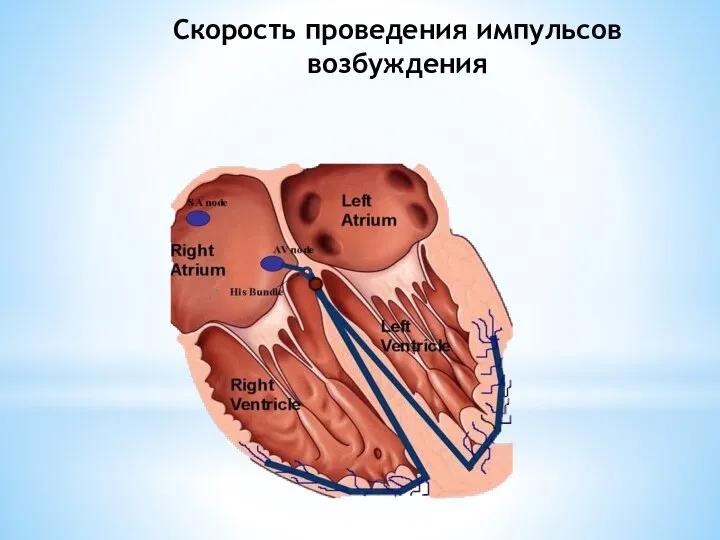
Слайд 21В различных участках сердца скорость проведения возбуждения неодинакова.
В синоатриальном узле скорость
В различных участках сердца скорость проведения возбуждения неодинакова.
В синоатриальном узле скорость

Слайд 22СОКРАТИМОСТЬ
Амплитуда одиночных сокращений миокарда зависит от частоты сокращений и величины растяжения волокон
СОКРАТИМОСТЬ
Амплитуда одиночных сокращений миокарда зависит от частоты сокращений и величины растяжения волокон

Слайд 23Лестница Боудича
Это закон гомеометрической саморегуляции клеток сердца (хроно-инотропная зависимость).
Только во второй половине
Лестница Боудича
Это закон гомеометрической саморегуляции клеток сердца (хроно-инотропная зависимость).
Только во второй половине

Слайд 24Гетерометрическая саморегуляция сокращений (закон Франка-Старлинга)
Это зависимость силы сокращений от величины растяжения мышечных
Гетерометрическая саморегуляция сокращений (закон Франка-Старлинга)
Это зависимость силы сокращений от величины растяжения мышечных

 Оперативное акушерство. Кесарево сечение
Оперативное акушерство. Кесарево сечение Срс. Антидепрессанты при воспалительных заболеваниях кишечника
Срс. Антидепрессанты при воспалительных заболеваниях кишечника Противопоказания к проведению МРТ
Противопоказания к проведению МРТ Рак желудка и предраковый состояния
Рак желудка и предраковый состояния Природа - найкращий лiкар
Природа - найкращий лiкар Коклюш
Коклюш Применение ЛС в аллергологии у беременных и в период лактации
Применение ЛС в аллергологии у беременных и в период лактации ХОБЛ
ХОБЛ Сердечно-сосудистые исходы в исследованиях ISGLT2
Сердечно-сосудистые исходы в исследованиях ISGLT2 Обезболивающие и сосудосуживающие средства
Обезболивающие и сосудосуживающие средства Стоп ВИЧ/СПИД. О проведении на территории Смоленской области мероприятий в рамках восьмой Всероссийской акции
Стоп ВИЧ/СПИД. О проведении на территории Смоленской области мероприятий в рамках восьмой Всероссийской акции Действие высоких и низких температур. Электротравма. Баротравма. (Тема 6)
Действие высоких и низких температур. Электротравма. Баротравма. (Тема 6) Заболевания сердечно-сосудистой системы. Атеросклероз
Заболевания сердечно-сосудистой системы. Атеросклероз Ишемическая болезнь сердца (ИБС). Стенокардия
Ишемическая болезнь сердца (ИБС). Стенокардия Лабораторное обследование больных с ОРВИ/гриппом
Лабораторное обследование больных с ОРВИ/гриппом Классификация психопатий
Классификация психопатий Острый и хронический пиелонефрит
Острый и хронический пиелонефрит Клинико-лабораторные особенности геморрагического синдрома при гемофилии
Клинико-лабораторные особенности геморрагического синдрома при гемофилии Маски акушерских осложнений: дифференциальная диагностика в клинической практике
Маски акушерских осложнений: дифференциальная диагностика в клинической практике Профилактика инфекционных болезней плотоядных животных
Профилактика инфекционных болезней плотоядных животных Здравоохранение будущего
Здравоохранение будущего Що таке дискордантна пара
Що таке дискордантна пара Презентация к выступлению Плюснина
Презентация к выступлению Плюснина Центральное стерилизационное отделение
Центральное стерилизационное отделение Генотерапия
Генотерапия Кровообращение
Кровообращение Интеллектуалды ойындар окушылардын кабiлеттерiн дамыту куралы ретiнде
Интеллектуалды ойындар окушылардын кабiлеттерiн дамыту куралы ретiнде Средства, нормализующие процессы обмена веществ и уплотняющие твердые ткани зуба
Средства, нормализующие процессы обмена веществ и уплотняющие твердые ткани зуба