- Белки и ферменты

Содержание
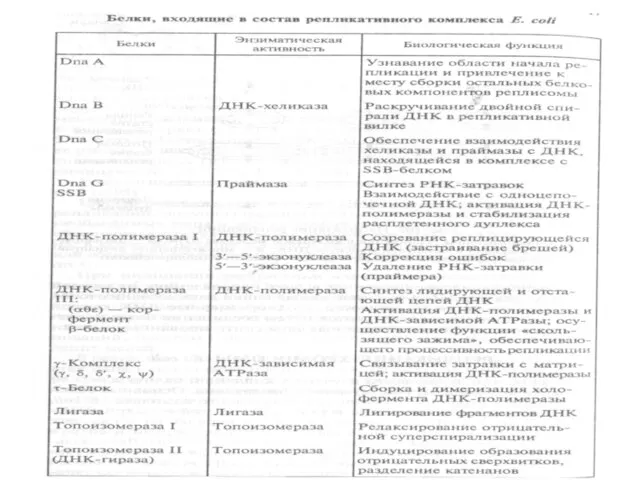
- 2. ДНК-полимеразы присутствуют во всех клетках про- и эукариот. Некоторые прокариотические и эукариотические ДНК-полимеразы выделены в чистом
- 4. ДНК-пoлимераза I E. coli Выделена А. Корнбергом и сотр. в 1958 г. Изучена наиболее полно. Представляет
- 5. Схема удвоения молекулы ДНК при репликации
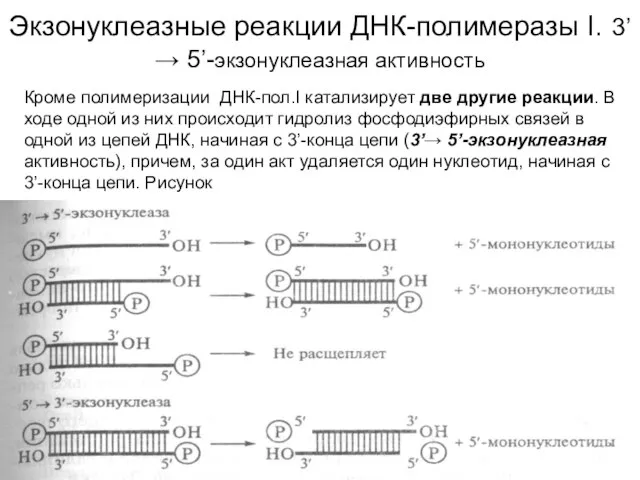
- 6. Экзонуклеазные реакции ДНК-полимеразы I. 3’→ 5’-экзонуклеазная активность Кроме полимеризации ДНК-пол.I катализирует две другие реакции. В ходе
- 7. 5’→ 3’-экзонуклеазная активность ДНК-пол.I Вторая реакция также заключается в отщеплении нуклеотидов, но гидролиз идет с 5’-конца
- 8. Ник-трансляция •ДНК-пол. I способна удлинять 3’-конец одной из цепей ДНК в месте разрывов и одновременно удалять
- 9. ДНК-полимераза II Две другие ДНК-полимеразы присутствуют в клетках E. сoli в меньших количествах. ДНК-пол. II (мол.масса
- 10. ДНК-полимераза III-холофермент Ключевой фермент, ответственный за репликацию ДНК E. coli. Кор- фермент ДНК-пол. III состоит из
- 11. ДНК-полимераза III-холофермент ДНК-полимераза III (мол. масса 103 кДа) — играет главную роль в репли- кации ДНК
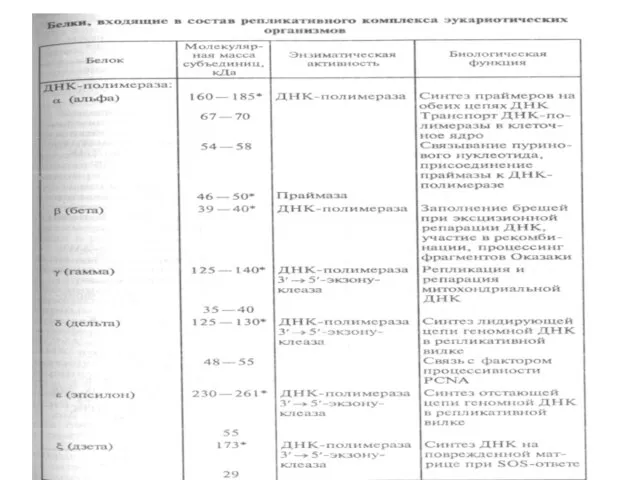
- 12. ДНК-полимеразы эукариот Механизмы репликации ДНК у эукариот менее изучены из-за их большей сложности. Основные результаты получены
- 14. ДНК-полимераза α — первая ДНК-полимераза, обнаружен- ная в клетках эукариот. Она представлена в клетке в виде
- 15. ДНК-полимераза δ — гетеродимер, состоящий из каталити- ческой субъединицы (125—130 кДа) и субъединицы 48 — 55
- 16. ДНК-полимераза ε, выделена из клеток HeLa, содержит два полипептида — каталитический 261 кДа и полипептид 55
- 17. ДНК-полимераза γ локализована в митохондриях, ее функция связана с репликацией и репарацией митохондриальной ДНК, она кодируется
- 18. Таким образом, поскольку эукариотические ДНК-полимеразы α и β лишены 3′ → 5 ′ и 5 ′
- 19. Модель работы димерной полимеразы(ДНК-пол.III); координация синтеза ДНК на комплементарных цепях Элонгация репликации. Модель полуконсерва- тивной прерывистой
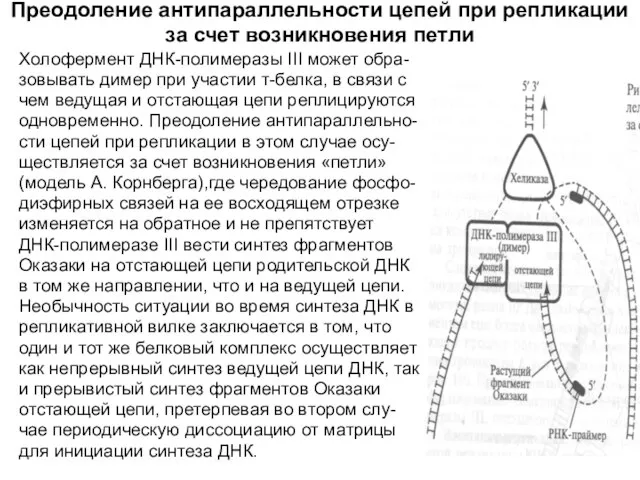
- 20. Преодоление антипараллельности цепей при репликации за счет возникновения петли Холофермент ДНК-полимеразы III может обра- зовывать димер
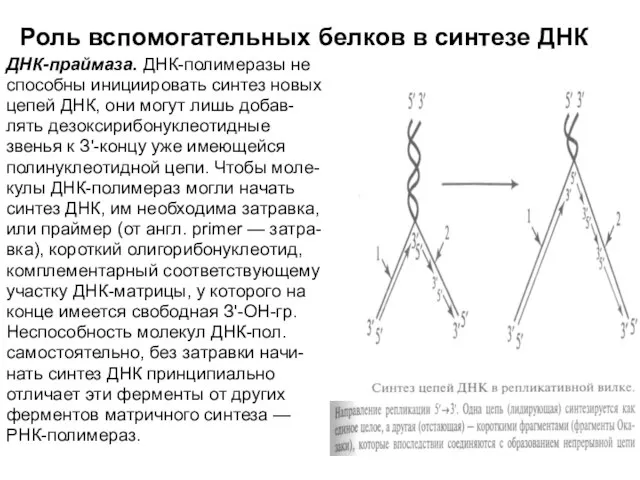
- 21. Роль вспомогательных белков в синтезе ДНК ДНК-праймаза. ДНК-полимеразы не способны инициировать синтез новых цепей ДНК, они
- 22. На стадии инициации репликации короткую РНК-затравку из рибонуклеозидтрифосфатов синтезирует фермент, называ- емый ДНК-праймазой. ДНК-праймаза может быть
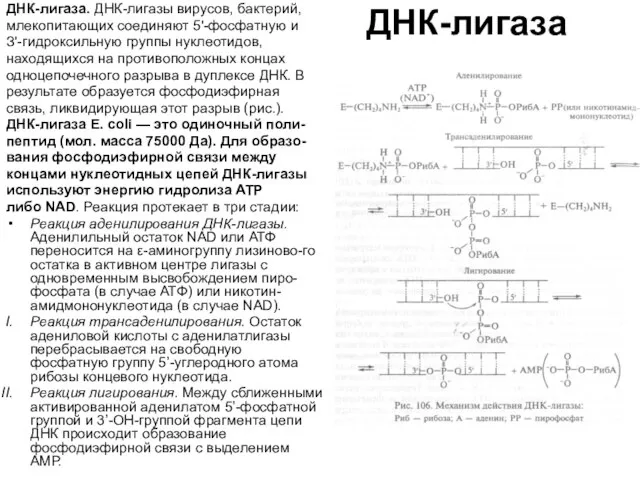
- 23. ДНК-лигаза ДНК-лигаза. ДНК-лигазы вирусов, бактерий, млекопитающих соединяют 5'-фосфатную и З'-гидроксильную группы нуклеотидов, находящихся на противоположных концах
- 24. Хеликаза Раскручивание, или расплетание, спирали происходит в локальном участке ДНК. Эту реакцию осуществляет хеликаза — ДНК-зависимая
- 25. ДСБ-белки (SSB-белки) Белки, связывающиеся с одноцепочечной ДНК. ДСБ-белки связываются с сахарофосфатным остовом одиночных цепей ДНК, не
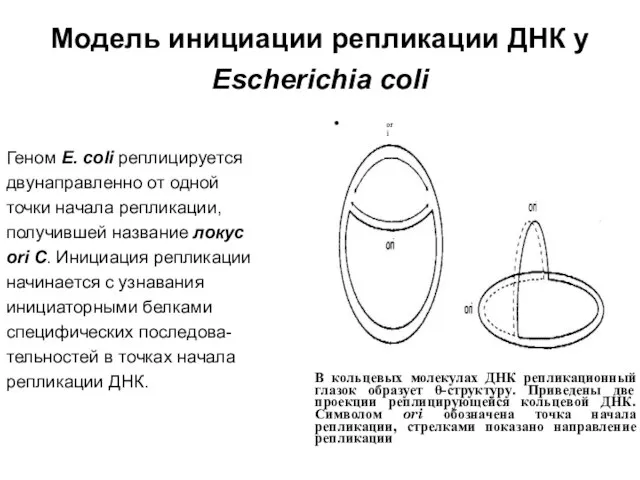
- 26. Модель инициации репликации ДНК у Escherichia coli Геном Е. coli реплицируется двунаправленно от одной точки начала
- 27. Регуляция инициации репликации у E. coli Хромосома Е. coli содержит единственную область начала репликации (oriC), размер
- 28. Белок Dna В (хеликаза) в виде гексамеров в комплексе с шестью мономерами белка Dna С, каждый
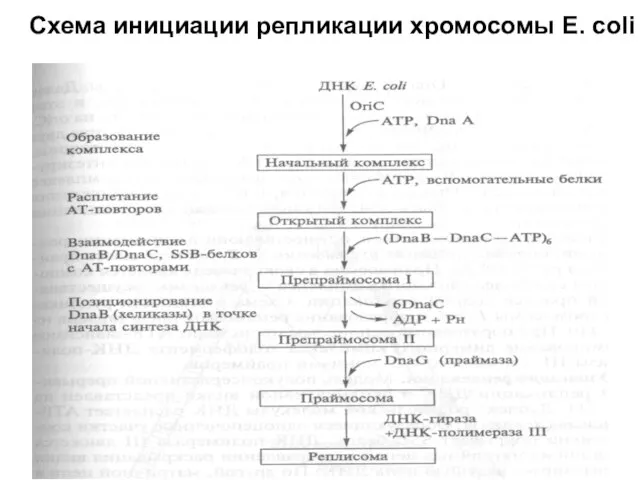
- 29. Схема инициации репликации хромосомы E. coli
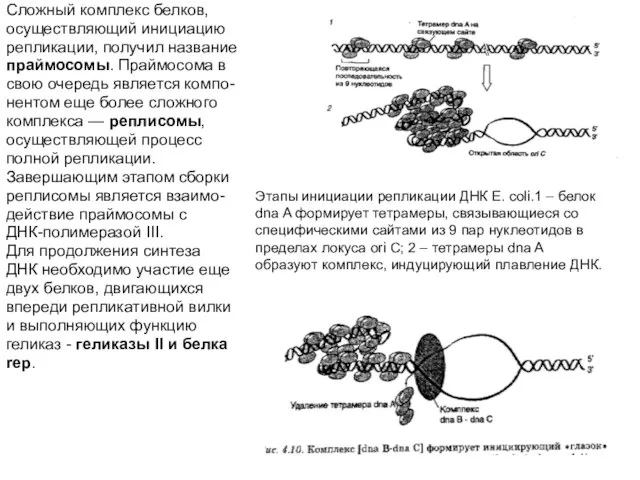
- 30. Этапы инициации репликации ДНК E. coli.1 – белок dna A формирует тетрамеры, связывающиеся со специфическими сайтами
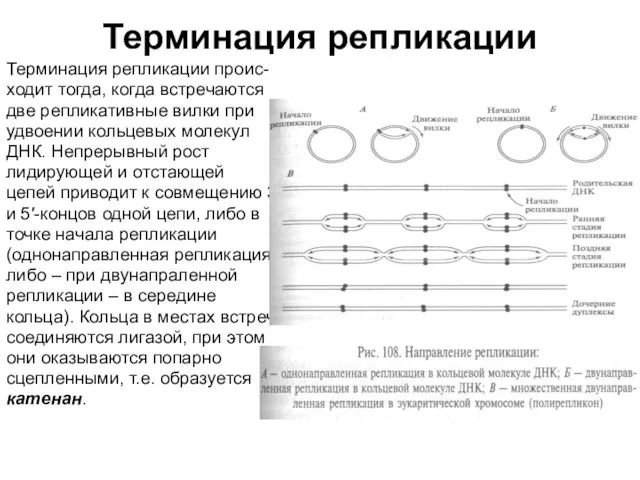
- 31. Терминация репликации Терминация репликации проис- ходит тогда, когда встречаются две репликативные вилки при удвоении кольцевых молекул
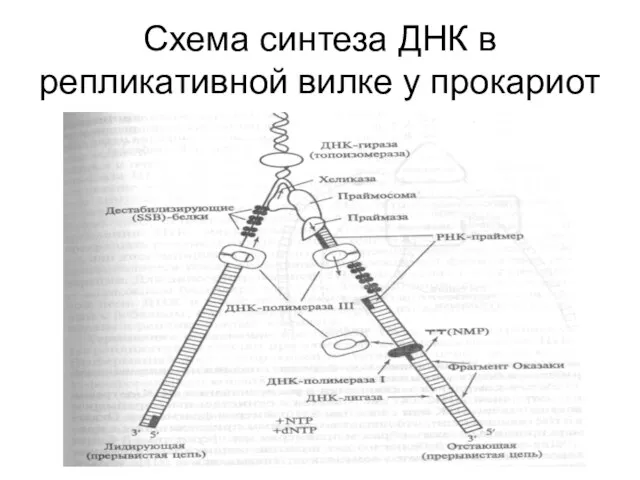
- 32. Схема синтеза ДНК в репликативной вилке у прокариот
- 33. Бокуть, с.122
- 34. Репликация ДНК у эукариот Рассмотрим особенности репликации эукариот на примере дрожжей – Saccharomyces сеrеvisiae. Точки начала
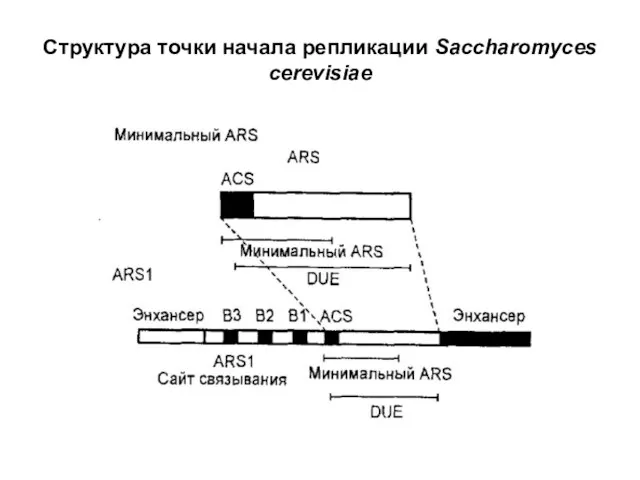
- 35. Структура точки начала репликации Saccharomyces cerevisiae
- 37. Скачать презентацию
Слайд 2ДНК-полимеразы присутствуют во всех клетках про- и
эукариот. Некоторые прокариотические и эукариотические
ДНК-полимеразы присутствуют во всех клетках про- и
эукариот. Некоторые прокариотические и эукариотические

Слайд 4ДНК-пoлимераза I E. coli
Выделена А. Корнбергом и сотр. в 1958 г.
ДНК-пoлимераза I E. coli
Выделена А. Корнбергом и сотр. в 1958 г.

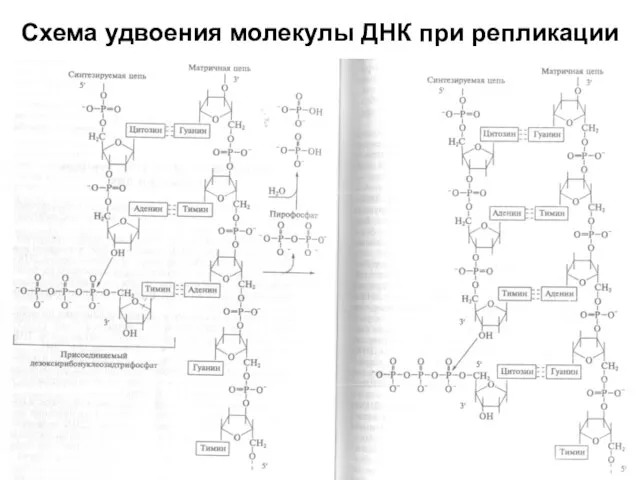
Слайд 5Схема удвоения молекулы ДНК при репликации
Схема удвоения молекулы ДНК при репликации
Слайд 6Экзонуклеазные реакции ДНК-полимеразы I. 3’→ 5’-экзонуклеазная активность
Кроме полимеризации ДНК-пол.I катализирует две другие
Экзонуклеазные реакции ДНК-полимеразы I. 3’→ 5’-экзонуклеазная активность
Кроме полимеризации ДНК-пол.I катализирует две другие
Слайд 75’→ 3’-экзонуклеазная активность ДНК-пол.I
Вторая реакция также заключается в отщеплении нуклеотидов, но
гидролиз
5’→ 3’-экзонуклеазная активность ДНК-пол.I
Вторая реакция также заключается в отщеплении нуклеотидов, но
гидролиз

Слайд 8Ник-трансляция
•ДНК-пол. I способна удлинять 3’-конец одной из цепей ДНК в
месте разрывов
Ник-трансляция
•ДНК-пол. I способна удлинять 3’-конец одной из цепей ДНК в
месте разрывов

Слайд 9ДНК-полимераза II
Две другие ДНК-полимеразы присутствуют в клетках E. сoli в
меньших количествах.
ДНК-полимераза II
Две другие ДНК-полимеразы присутствуют в клетках E. сoli в
меньших количествах.

Слайд 10ДНК-полимераза III-холофермент
Ключевой фермент, ответственный за репликацию ДНК E. coli. Кор-
фермент ДНК-пол. III
ДНК-полимераза III-холофермент
Ключевой фермент, ответственный за репликацию ДНК E. coli. Кор-
фермент ДНК-пол. III

Слайд 11ДНК-полимераза III-холофермент
ДНК-полимераза III (мол. масса 103 кДа) — играет главную роль в
ДНК-полимераза III-холофермент
ДНК-полимераза III (мол. масса 103 кДа) — играет главную роль в

Слайд 12ДНК-полимеразы эукариот
Механизмы репликации ДНК у эукариот менее изучены из-за их большей
сложности.
ДНК-полимеразы эукариот
Механизмы репликации ДНК у эукариот менее изучены из-за их большей
сложности.

Слайд 14ДНК-полимераза α — первая ДНК-полимераза, обнаружен-
ная в клетках эукариот. Она представлена в
ДНК-полимераза α — первая ДНК-полимераза, обнаружен-
ная в клетках эукариот. Она представлена в

Слайд 15ДНК-полимераза δ — гетеродимер, состоящий из каталити-
ческой субъединицы (125—130 кДа) и субъединицы
ДНК-полимераза δ — гетеродимер, состоящий из каталити-
ческой субъединицы (125—130 кДа) и субъединицы

Слайд 16ДНК-полимераза ε, выделена из клеток HeLa, содержит два
полипептида — каталитический 261
ДНК-полимераза ε, выделена из клеток HeLa, содержит два
полипептида — каталитический 261

Слайд 17ДНК-полимераза γ локализована в митохондриях, ее функция связана с
репликацией и репарацией
ДНК-полимераза γ локализована в митохондриях, ее функция связана с
репликацией и репарацией

Слайд 18Таким образом, поскольку эукариотические
ДНК-полимеразы α и β лишены 3′ → 5
Таким образом, поскольку эукариотические
ДНК-полимеразы α и β лишены 3′ → 5

Слайд 19Модель работы димерной полимеразы(ДНК-пол.III); координация синтеза ДНК на комплементарных цепях
Элонгация репликации. Модель
Модель работы димерной полимеразы(ДНК-пол.III); координация синтеза ДНК на комплементарных цепях
Элонгация репликации. Модель

Слайд 20Преодоление антипараллельности цепей при репликации за счет возникновения петли
Холофермент ДНК-полимеразы III может
Преодоление антипараллельности цепей при репликации за счет возникновения петли
Холофермент ДНК-полимеразы III может
Слайд 21Роль вспомогательных белков в синтезе ДНК
ДНК-праймаза. ДНК-полимеразы не
способны инициировать синтез новых
Роль вспомогательных белков в синтезе ДНК
ДНК-праймаза. ДНК-полимеразы не
способны инициировать синтез новых
Слайд 22На стадии инициации репликации короткую РНК-затравку из
рибонуклеозидтрифосфатов синтезирует фермент, называ-
емый ДНК-праймазой.
На стадии инициации репликации короткую РНК-затравку из
рибонуклеозидтрифосфатов синтезирует фермент, называ-
емый ДНК-праймазой.

Слайд 23ДНК-лигаза
ДНК-лигаза. ДНК-лигазы вирусов, бактерий,
млекопитающих соединяют 5'-фосфатную и
З'-гидроксильную группы нуклеотидов,
находящихся
ДНК-лигаза
ДНК-лигаза. ДНК-лигазы вирусов, бактерий,
млекопитающих соединяют 5'-фосфатную и
З'-гидроксильную группы нуклеотидов,
находящихся
Слайд 24Хеликаза
Раскручивание, или расплетание, спирали происходит в локальном
участке ДНК. Эту реакцию осуществляет
Хеликаза
Раскручивание, или расплетание, спирали происходит в локальном
участке ДНК. Эту реакцию осуществляет

Слайд 25ДСБ-белки (SSB-белки)
Белки, связывающиеся с одноцепочечной ДНК. ДСБ-белки
связываются с сахарофосфатным остовом одиночных
ДСБ-белки (SSB-белки)
Белки, связывающиеся с одноцепочечной ДНК. ДСБ-белки
связываются с сахарофосфатным остовом одиночных

Слайд 26Модель инициации репликации ДНК у Escherichia coli
Геном Е. coli реплицируется
двунаправленно
Модель инициации репликации ДНК у Escherichia coli
Геном Е. coli реплицируется
двунаправленно
Слайд 27Регуляция инициации репликации у E. coli
Хромосома Е. coli содержит единственную область начала
Регуляция инициации репликации у E. coli
Хромосома Е. coli содержит единственную область начала

Слайд 28Белок Dna В (хеликаза) в виде гексамеров в комплексе с шестью
мономерами
Белок Dna В (хеликаза) в виде гексамеров в комплексе с шестью
мономерами

Слайд 29Схема инициации репликации хромосомы E. coli
Схема инициации репликации хромосомы E. coli
Слайд 30Этапы инициации репликации ДНК E. coli.1 – белок dna A формирует тетрамеры,
Этапы инициации репликации ДНК E. coli.1 – белок dna A формирует тетрамеры,
Слайд 31Терминация репликации
Терминация репликации проис-
ходит тогда, когда встречаются
две репликативные вилки при
удвоении
Терминация репликации
Терминация репликации проис-
ходит тогда, когда встречаются
две репликативные вилки при
удвоении
Слайд 32Схема синтеза ДНК в репликативной вилке у прокариот
Схема синтеза ДНК в репликативной вилке у прокариот
Слайд 33Бокуть, с.122
Бокуть, с.122

Слайд 34Репликация ДНК у эукариот
Рассмотрим особенности репликации эукариот на примере дрожжей –
Saccharomyces
Репликация ДНК у эукариот
Рассмотрим особенности репликации эукариот на примере дрожжей –
Saccharomyces

Слайд 35Структура точки начала репликации Saccharomyces cerevisiae
Структура точки начала репликации Saccharomyces cerevisiae
 Архитектура и функционирование персонального компьютера
Архитектура и функционирование персонального компьютера Каталог TopFranchise. Коммерческое предложение по сотрудничеству
Каталог TopFranchise. Коммерческое предложение по сотрудничеству Лот 16, г. Хабаровск, ул. Сысоева, 21, кв. 32
Лот 16, г. Хабаровск, ул. Сысоева, 21, кв. 32 Тема. Предмет исследования (шаблон)
Тема. Предмет исследования (шаблон) Воссоздающее и творческое воображение
Воссоздающее и творческое воображение Специальные звания и сроки выслуги
Специальные звания и сроки выслуги Федеральный государственный образовательный стандарт основного общего образования
Федеральный государственный образовательный стандарт основного общего образования Ректорат_26092022
Ректорат_26092022 Стиль в искусстве – это мироощущение времени.
Стиль в искусстве – это мироощущение времени. «Ах, как я зол!.. Как я зла, Как я зла!», или Превратности любви
«Ах, как я зол!.. Как я зла, Как я зла!», или Превратности любви Польза мёда
Польза мёда Анксиолитки. Тревожные расстройства
Анксиолитки. Тревожные расстройства Present Simple (Indefinite) and Present Continuous(Progressive)
Present Simple (Indefinite) and Present Continuous(Progressive) Презентация на тему Демографическая ситуация в России
Презентация на тему Демографическая ситуация в России Severe flooding in Brazil and Australia
Severe flooding in Brazil and Australia  През. Введение в предмет информатики
През. Введение в предмет информатики 18.08 план развития музея Древние курганы Салбыкской степи (1)
18.08 план развития музея Древние курганы Салбыкской степи (1) Год культурного наследия народов России
Год культурного наследия народов России Создание электронного курса
Создание электронного курса ПЛАНИРОВАНИЕ СЕБЕСТОИМОСТИ, ПРИБЫЛИ И РЕНТАБЕЛЬНОСТИ
ПЛАНИРОВАНИЕ СЕБЕСТОИМОСТИ, ПРИБЫЛИ И РЕНТАБЕЛЬНОСТИ Скульптура и рельеф Древней Греции
Скульптура и рельеф Древней Греции Отдых в Крыму – Профилакторий “Солнечный”
Отдых в Крыму – Профилакторий “Солнечный” Т.Г. Шевченко-художник
Т.Г. Шевченко-художник Личностно-ориентированное воспитание младших школьников как фактор их социализации
Личностно-ориентированное воспитание младших школьников как фактор их социализации Элементы баскетбола на уроках в начальной школе
Элементы баскетбола на уроках в начальной школе Жанры цирка
Жанры цирка Презентация на тему Традиционный японский дом
Презентация на тему Традиционный японский дом  Подарочные наборы
Подарочные наборы