- Фиксация молекулярного азота

Содержание
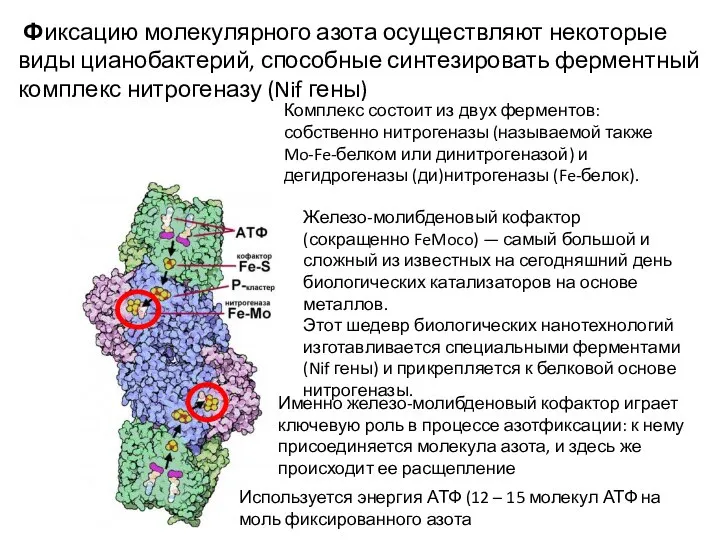
- 2. Комплекс состоит из двух ферментов: собственно нитрогеназы (называемой также Mo-Fe-белком или динитрогеназой) и дегидрогеназы (ди)нитрогеназы (Fe-белок).
- 3. Новые данные (на основе молекулярно-генетических методов): Процесс азотфиксации «стартовал» по крайней мере 3–3,5 млрд лет назад,
- 4. Гетероцисты – результат клеточной дифференциации. Дезактивация ФС2: хл-белковые комплексы и компоненты ФС2 разрушаются (активность ФС1 сохраняется);
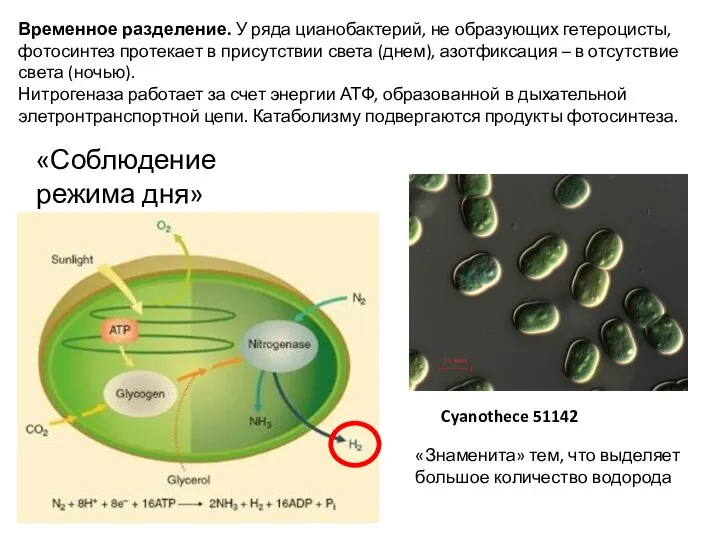
- 5. Cyanothece 51142 Временное разделение. У ряда цианобактерий, не образующих гетероцисты, фотосинтез протекает в присутствии света (днем),
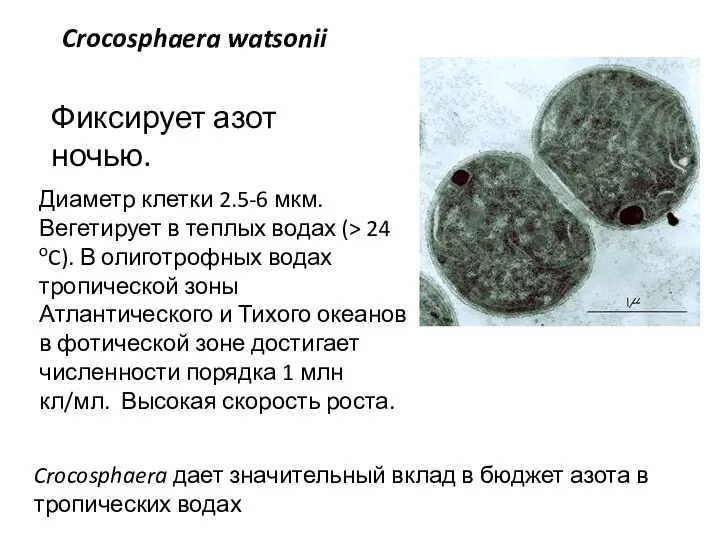
- 6. Диаметр клетки 2.5-6 мкм. Вегетирует в теплых водах (> 24 оC). В олиготрофных водах тропической зоны
- 7. Trichodesmium spp. Цианобактерии Trichodesmium фиксируют азот ТОЛЬКО на свету и при этом у них НЕТ гетероцист
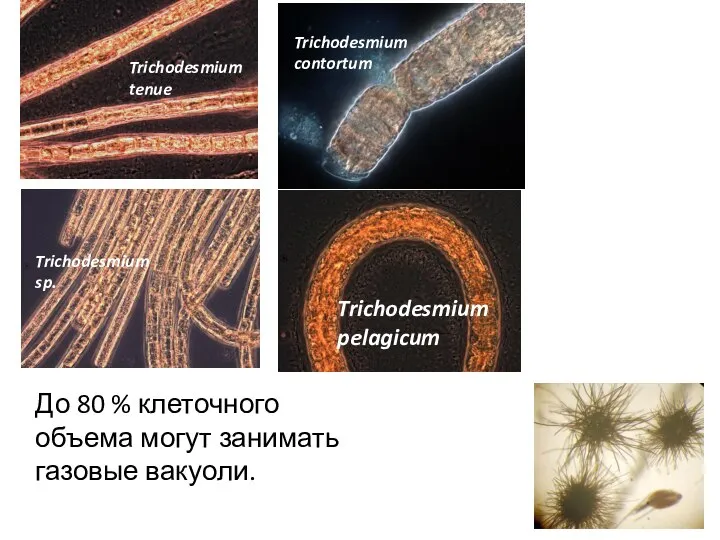
- 8. Trichodesmium tenue Trichodesmium sp. Trichodesmium contortum Trichodesmium pelagicum До 80 % клеточного объема могут занимать газовые
- 9. Как же осуществляется защита нитрогеназ от фотосинтетически выделяемого кислорода у Trichodesmium? Как экспрессия Nif генов, так
- 10. Разделение азотфиксации и фотосинтеза в пространстве у Trichodesmium Окраска специфическим красителем Nif белков Фиксация азота идет
- 11. Разделение азотфиксации и фотосинтеза во времени у Trichodesmium Суточный ритм регулирует транскрипцию nif генов, синтез de
- 12. Способность к фиксации СО2 также снижается в диазоцитах Trichodesmium во время полуденной депрессии, при этом возрастает
- 13. Механизмы предотвращения блокировки нитрогеназ фотосинтетически выделяемым кислородом, ПРЕДПОЛОЖИТЕЛЬНО, также включают: тетразол Формирование в диазоцитах восстановленных зон
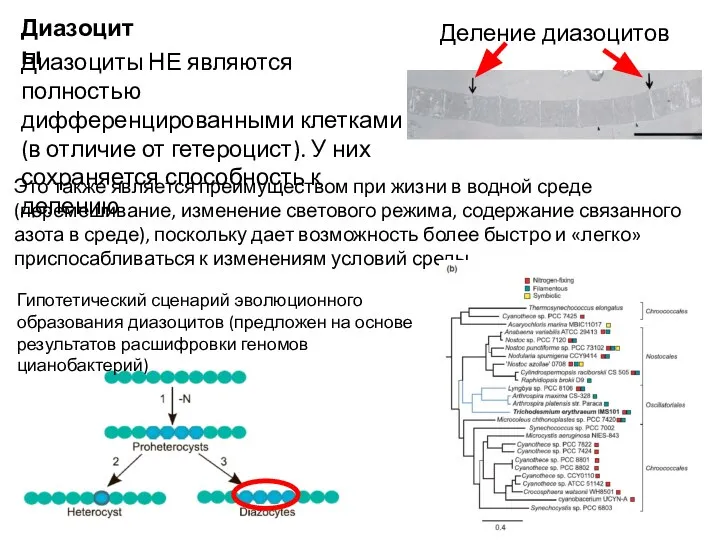
- 14. Гипотетический сценарий эволюционного образования диазоцитов (предложен на основе результатов расшифровки геномов цианобактерий) Диазоциты Диазоциты НЕ являются
- 15. Дальнейшая ассимиляция глутамата, образованного при азотфиксации, происходит в глутаминсинтетазо - глутамин оксоглутарат аминотрансферазном (ГС-ГОГАТ) метаболитном цикле
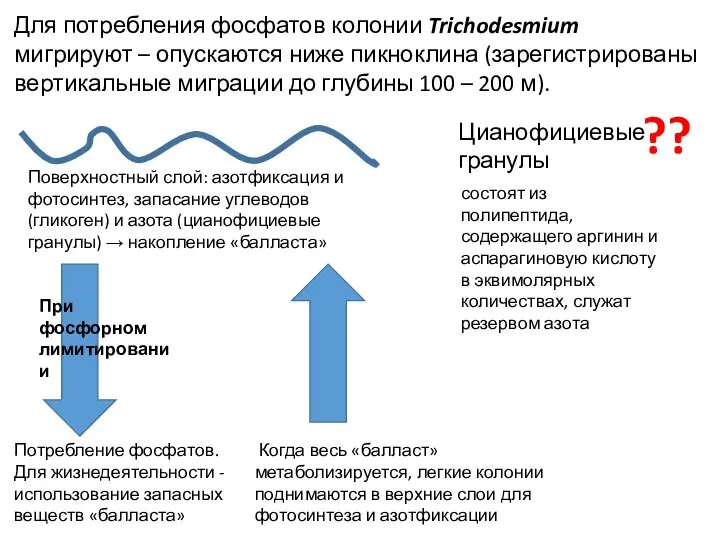
- 16. Для потребления фосфатов колонии Trichodesmium мигрируют – опускаются ниже пикноклина (зарегистрированы вертикальные миграции до глубины 100
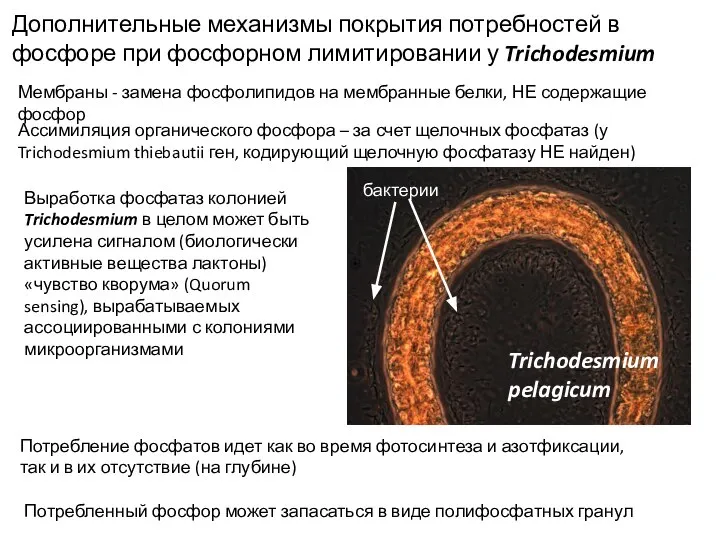
- 17. Дополнительные механизмы покрытия потребностей в фосфоре при фосфорном лимитировании у Trichodesmium Мембраны - замена фосфолипидов на
- 18. Обеспечение потребностей в железе у Trichodesmium В клетках Trichodesmium высокое содержание Fe. Учитывая, что массовое развитие
- 19. Лимитирование недостатком железа ведет к снижению числа диазоцитов и скорости азофиксации А вот фотосинтетическая активность продолжает
- 20. Экосистемная роль Trichodesmium Пределы температурной толерантности для роста и азотфиксации у Trichodesmium: 20 – 34 оС,
- 21. Цветение Trichodesmium. Большой барьерный риф побережья Австралии. Снимок NASA Цветение Trichodesmium. Красное море Снимок NASA Массовое
- 22. Для цветений Trichodesmium характерно как прижизненное выделение значительно количества органического вещества, так и при отмирании клеток
- 23. Это довольно НЕОЖИДАННЫЙ результат, поскольку Trichodesmium синтезирует токсины, вызывающие гибель копепод, таких как Acartia tonsa, а
- 24. Прогнозы динамики Trichodesmium при наблюдаемом климатическом тренде Есть все основания полагать, что повышение температуры и концентрации
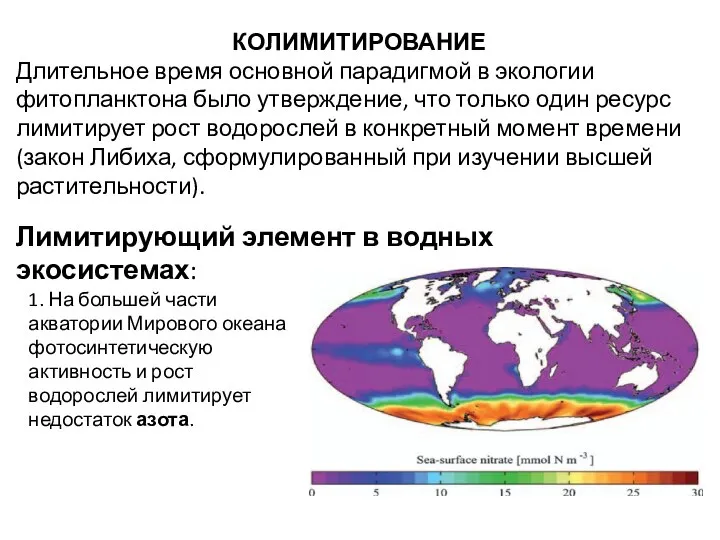
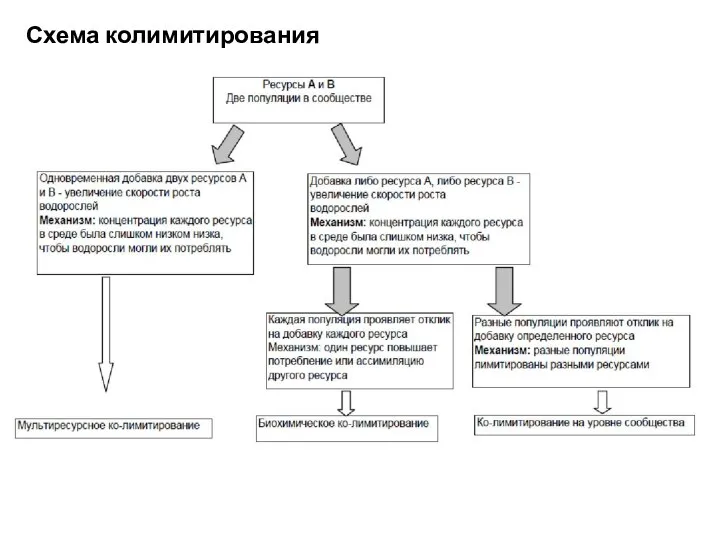
- 25. КОЛИМИТИРОВАНИЕ Длительное время основной парадигмой в экологии фитопланктона было утверждение, что только один ресурс лимитирует рост
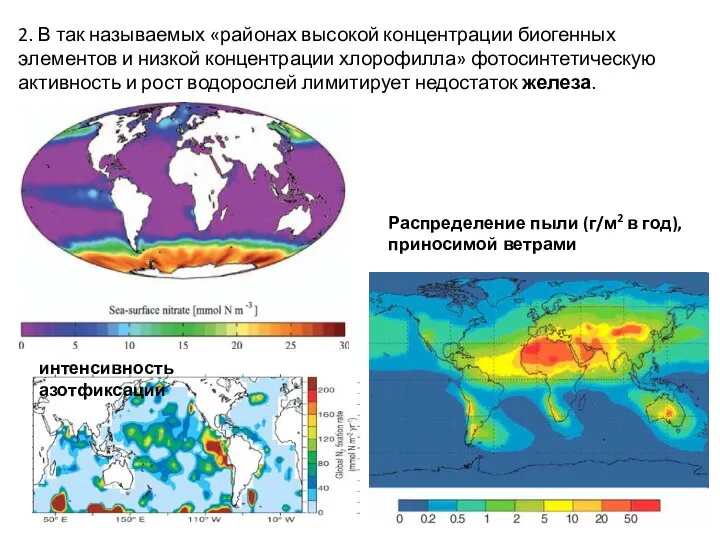
- 26. 2. В так называемых «районах высокой концентрации биогенных элементов и низкой концентрации хлорофилла» фотосинтетическую активность и
- 27. 3. В ряде районов Мирового океана (Северная Атлантика, Средиземное море, Бискайский залив, норвежские фиорды, Белое море)
- 28. Мультиресурсное ко-лимитирование. Два ресурса в такой низкой концентрации (ниже порогового уровня), что их потребление уже невозможно.
- 29. Колимитирование на уровне сообщества. Добавка ресурса А стимулирует рост одной популяции, добавка ресурса В стимулирует другую.
- 30. Схема колимитирования
- 31. На следующей лекции: Пространственно-временная изменчивость первичной продукции в Мировом океане Вопросы?
- 33. Скачать презентацию
Слайд 2Комплекс состоит из двух ферментов: собственно нитрогеназы (называемой также Mo-Fe-белком или динитрогеназой)
Комплекс состоит из двух ферментов: собственно нитрогеназы (называемой также Mo-Fe-белком или динитрогеназой)
Слайд 3Новые данные (на основе молекулярно-генетических методов):
Процесс азотфиксации «стартовал» по крайней мере 3–3,5
Новые данные (на основе молекулярно-генетических методов):
Процесс азотфиксации «стартовал» по крайней мере 3–3,5

Слайд 4Гетероцисты – результат клеточной дифференциации.
Дезактивация ФС2: хл-белковые комплексы и компоненты ФС2 разрушаются
Гетероцисты – результат клеточной дифференциации.
Дезактивация ФС2: хл-белковые комплексы и компоненты ФС2 разрушаются

Слайд 5Cyanothece 51142
Временное разделение. У ряда цианобактерий, не образующих гетероцисты, фотосинтез протекает в
Cyanothece 51142
Временное разделение. У ряда цианобактерий, не образующих гетероцисты, фотосинтез протекает в
Слайд 6Диаметр клетки 2.5-6 мкм. Вегетирует в теплых водах (> 24 оC). В
Диаметр клетки 2.5-6 мкм. Вегетирует в теплых водах (> 24 оC). В
Слайд 7Trichodesmium spp.
Цианобактерии Trichodesmium фиксируют азот ТОЛЬКО на свету и при этом у
Trichodesmium spp.
Цианобактерии Trichodesmium фиксируют азот ТОЛЬКО на свету и при этом у

Слайд 8Trichodesmium tenue
Trichodesmium sp.
Trichodesmium contortum
Trichodesmium pelagicum
До 80 % клеточного объема могут занимать газовые
Trichodesmium tenue
Trichodesmium sp.
Trichodesmium contortum
Trichodesmium pelagicum
До 80 % клеточного объема могут занимать газовые
Слайд 9Как же осуществляется защита нитрогеназ от фотосинтетически выделяемого кислорода у Trichodesmium?
Как экспрессия
Как же осуществляется защита нитрогеназ от фотосинтетически выделяемого кислорода у Trichodesmium?
Как экспрессия

Слайд 10Разделение азотфиксации и фотосинтеза в пространстве у Trichodesmium
Окраска специфическим красителем Nif белков
Фиксация
Разделение азотфиксации и фотосинтеза в пространстве у Trichodesmium
Окраска специфическим красителем Nif белков
Фиксация

Слайд 11Разделение азотфиксации и фотосинтеза во времени у Trichodesmium
Суточный ритм регулирует транскрипцию nif
Разделение азотфиксации и фотосинтеза во времени у Trichodesmium
Суточный ритм регулирует транскрипцию nif

Слайд 12Способность к фиксации СО2 также снижается в диазоцитах Trichodesmium во время полуденной
Способность к фиксации СО2 также снижается в диазоцитах Trichodesmium во время полуденной

Слайд 13Механизмы предотвращения блокировки нитрогеназ фотосинтетически выделяемым кислородом, ПРЕДПОЛОЖИТЕЛЬНО, также включают:
тетразол
Формирование в диазоцитах
Механизмы предотвращения блокировки нитрогеназ фотосинтетически выделяемым кислородом, ПРЕДПОЛОЖИТЕЛЬНО, также включают:
тетразол
Формирование в диазоцитах

Слайд 14Гипотетический сценарий эволюционного образования диазоцитов (предложен на основе результатов расшифровки геномов цианобактерий)
Диазоциты
Диазоциты
Гипотетический сценарий эволюционного образования диазоцитов (предложен на основе результатов расшифровки геномов цианобактерий)
Диазоциты
Диазоциты
Слайд 15Дальнейшая ассимиляция глутамата, образованного при азотфиксации, происходит в глутаминсинтетазо - глутамин оксоглутарат
Дальнейшая ассимиляция глутамата, образованного при азотфиксации, происходит в глутаминсинтетазо - глутамин оксоглутарат

Слайд 16Для потребления фосфатов колонии Trichodesmium мигрируют – опускаются ниже пикноклина (зарегистрированы вертикальные
Для потребления фосфатов колонии Trichodesmium мигрируют – опускаются ниже пикноклина (зарегистрированы вертикальные
Слайд 17Дополнительные механизмы покрытия потребностей в фосфоре при фосфорном лимитировании у Trichodesmium
Мембраны -
Дополнительные механизмы покрытия потребностей в фосфоре при фосфорном лимитировании у Trichodesmium
Мембраны -
Слайд 18Обеспечение потребностей в железе у Trichodesmium
В клетках Trichodesmium высокое содержание Fe.
Обеспечение потребностей в железе у Trichodesmium
В клетках Trichodesmium высокое содержание Fe.

Слайд 19Лимитирование недостатком железа ведет к снижению числа диазоцитов и скорости азофиксации
А вот
Лимитирование недостатком железа ведет к снижению числа диазоцитов и скорости азофиксации
А вот

Слайд 20Экосистемная роль Trichodesmium
Пределы температурной толерантности для роста и азотфиксации у Trichodesmium:
Экосистемная роль Trichodesmium
Пределы температурной толерантности для роста и азотфиксации у Trichodesmium:

Слайд 21Цветение Trichodesmium.
Большой барьерный риф побережья Австралии.
Снимок NASA
Цветение Trichodesmium.
Красное море
Снимок NASA
Массовое
Цветение Trichodesmium.
Большой барьерный риф побережья Австралии.
Снимок NASA
Цветение Trichodesmium.
Красное море
Снимок NASA
Массовое

Слайд 22Для цветений Trichodesmium характерно как прижизненное выделение значительно количества органического вещества, так
Для цветений Trichodesmium характерно как прижизненное выделение значительно количества органического вещества, так

Слайд 23Это довольно НЕОЖИДАННЫЙ результат, поскольку Trichodesmium синтезирует токсины, вызывающие гибель копепод, таких
Это довольно НЕОЖИДАННЫЙ результат, поскольку Trichodesmium синтезирует токсины, вызывающие гибель копепод, таких

Слайд 24Прогнозы динамики Trichodesmium при наблюдаемом климатическом тренде
Есть все основания полагать, что повышение
Прогнозы динамики Trichodesmium при наблюдаемом климатическом тренде
Есть все основания полагать, что повышение

Слайд 25КОЛИМИТИРОВАНИЕ
Длительное время основной парадигмой в экологии фитопланктона было утверждение, что только один
КОЛИМИТИРОВАНИЕ
Длительное время основной парадигмой в экологии фитопланктона было утверждение, что только один
Слайд 262. В так называемых «районах высокой концентрации биогенных элементов и низкой концентрации
2. В так называемых «районах высокой концентрации биогенных элементов и низкой концентрации
Слайд 273. В ряде районов Мирового океана (Северная Атлантика, Средиземное море, Бискайский залив,
3. В ряде районов Мирового океана (Северная Атлантика, Средиземное море, Бискайский залив,

Слайд 28Мультиресурсное ко-лимитирование.
Два ресурса в такой низкой концентрации (ниже порогового уровня), что
Мультиресурсное ко-лимитирование.
Два ресурса в такой низкой концентрации (ниже порогового уровня), что

Слайд 29Колимитирование на уровне сообщества. Добавка ресурса А стимулирует рост одной популяции, добавка
Колимитирование на уровне сообщества. Добавка ресурса А стимулирует рост одной популяции, добавка

Слайд 30Схема колимитирования
Схема колимитирования
Слайд 31На следующей лекции:
Пространственно-временная изменчивость первичной продукции в Мировом океане
Вопросы?
На следующей лекции:
Пространственно-временная изменчивость первичной продукции в Мировом океане
Вопросы?

 СNet Attribute
СNet Attribute Разбор демоверсии теста задания 9-18
Разбор демоверсии теста задания 9-18 Воспитательное мероприятие профориентационного характера
Воспитательное мероприятие профориентационного характера Основные звенья стратегии профессионального роста педагога-психолога
Основные звенья стратегии профессионального роста педагога-психолога Кунгурская городская управа
Кунгурская городская управа Библиотека МБОУ «Школа№17»
Библиотека МБОУ «Школа№17» Презентация на тему Техника безопасности в кабинете информатики
Презентация на тему Техника безопасности в кабинете информатики Перловая крупа и блюда из нее
Перловая крупа и блюда из нее Создание отчетов с помощью «Конструктора»
Создание отчетов с помощью «Конструктора» Презентация на тему Русская игрушка - Матрешка
Презентация на тему Русская игрушка - Матрешка Мотивация персонала
Мотивация персонала Гражданское обществ и государство
Гражданское обществ и государство Swix poles
Swix poles Лыжная подготовка
Лыжная подготовка 03_LOGITECH. B2B C&P Pres
03_LOGITECH. B2B C&P Pres Анализ проекта
Анализ проекта Образовательная система будущего
Образовательная система будущего Задачи на движение
Задачи на движение МУП «Пассажирские перевозки г.Пензы».
МУП «Пассажирские перевозки г.Пензы». ГОУ ВПО «НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ»
ГОУ ВПО «НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ» Помада абсолютный поцелуй
Помада абсолютный поцелуй Презентація. Письмо. 20.09.22
Презентація. Письмо. 20.09.22 Зрение школьника
Зрение школьника Городская неделя музыки
Городская неделя музыки Трудовой договор
Трудовой договор Экспериментальная огневая отработка жидкостных ракетных двигателей малых тяг на стенде МАИ
Экспериментальная огневая отработка жидкостных ракетных двигателей малых тяг на стенде МАИ История развития науки криминалистики
История развития науки криминалистики Протоколы глобальных сетей
Протоколы глобальных сетей