- ФОРМЫ ИММУННОГО ОТВЕТА: АНТИТЕЛООБРАЗОВАНИЕ, иммунологическая память

Содержание
- 2. Формы проявления иммунного ответа: 1) антителообразование, 2) иммунный фагоцитоз, 3) опосредованный клетками киллинг, 4) реакции гиперчувствительности,
- 3. Все элементы иммунной системы реагируют на изменение гомеостаза, в зависимости от характера антигенного воздействия одни формы
- 4. Антитела — белки, специфически реагирующие с антигенами. Антитела и фагоцитоз - одна из наиболее филогенетически древних
- 5. Антитела — это γ-глобулины, вырабатываемые в ответ на введение антигена, способные специфически связываться с антигеном ,
- 6. Антитела используют для: 1. диагностики 2.профилактики 3.лечения различных заболеваний, 4.выделения и очистки биологически активных веществ. Для
- 7. Иммуноглобулины - это гликопротеиды. Они состоят из полипептидных цепей, стабилизированных сахаридными остатками. При нагревании выше 60
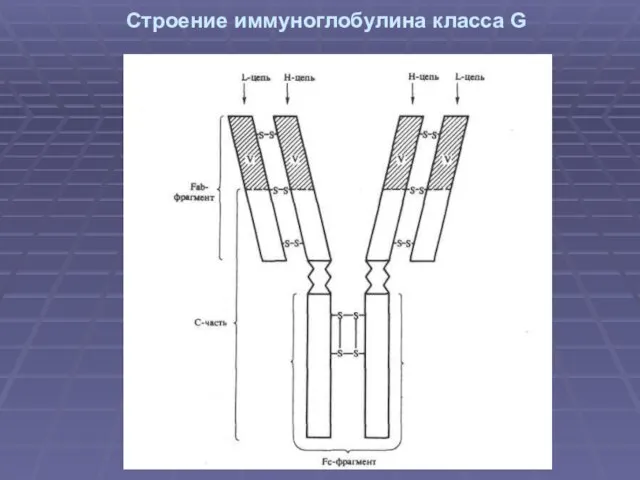
- 8. Строение иммуноглобулина класса G
- 9. Между тяжелыми цепями также есть дисульфидная связь . Такой тип межпептидного соединения позволяет менять конформацию в
- 10. Отдельные участки цепи молекулы Ig свернуты в глобулы (домены). Домены стабилизированы дисульфидной связью. В составе тяжелой
- 11. Вариабельные домены легкой и тяжелой цепи образуют участок, который специфически связывается с антигеном. Это антигенсвязывающий центр
- 12. Обработка ферментами молекулы Ig приводит к ее гидролизу. Папаин разрывает молекулу на три фрагмента. Два из
- 13. В зависимости от особенностей молекулярного строения тяжелой цепи различают 5 классов, или изотипов Ig . Молекулы,
- 14. Иммуноглобулин класса G. Изотип G составляет основную массу Ig сыворотки крови ( 70—80 %), при этом
- 15. IgGl и IgG3 связывают комплемент. IgG4, обладает цитофильностью (тропностью к тучным клеткам и базофилам) и участвует
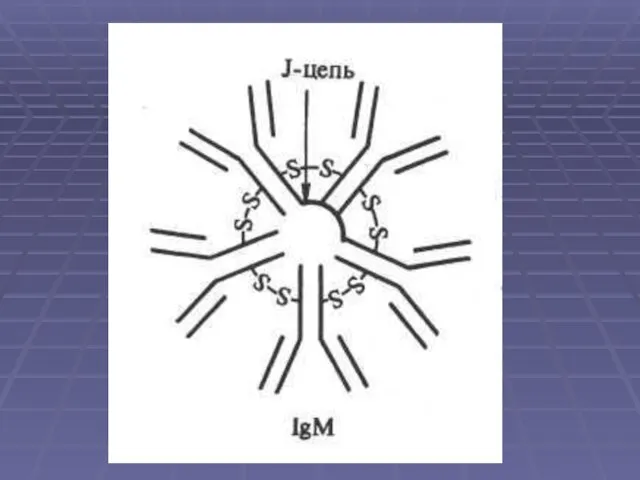
- 16. Иммуноглобулин класса М. Самая крупная молекула из всех Ig. Это пентамер, который имеет 10 антигенсвязывающих центров
- 18. IgM филогенетически наиболее древний иммуноглобулин. Синтезируется предшественниками и зрелыми В-лимфоцитами . Образуется в начале первичного иммунного
- 19. Не проходит через плаценту. Обнаружение специфических антител изотипа М в сыворотке крови новорожденного указывает на бывшую
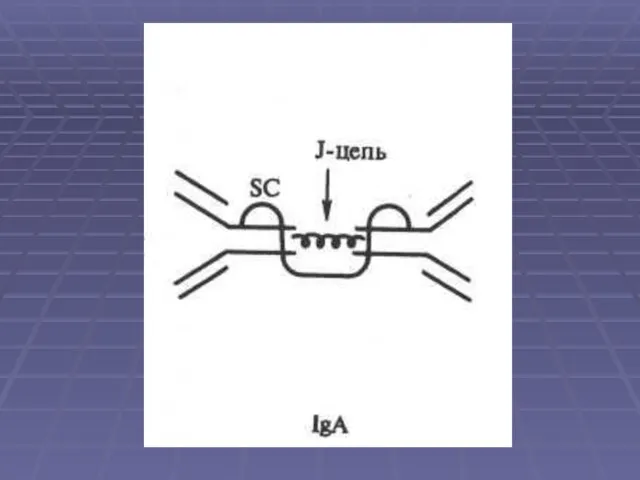
- 20. Иммуноглобулин класса А. Существует в сывороточной и секреторной формах. Около 60 % всех IgA содержится в
- 22. Может быть неполным антителом. Не связывает комплемент. Не проходит через плацентарный барьер. IgA обеспечивает нейтрализацию, опсонизацию
- 23. Молекулярная масса 350 кДа и выше, константа седиментации 13S и выше. Синтезируется В-лимфоцитами и плазматическими клетками
- 24. Секреторная форма IgA — основной фактор специфического гуморального местного иммунитета слизистых оболочек желудочно-кишечного тракта, мочеполовой системы
- 25. Иммуноглобулин класса Е ( реагин). Содержание в сыворотке крови — 0,00025 г/л. Молекулярная масса — около
- 26. Иммуноглобулин класса D. Концентрация в сыворотке крови - 0,03 г/л (около 0,2 % от общего числа
- 27. Рецепторные иммуноглобулины. Рецепторные, или мембраные Ig, локализуются на цитоплазматической мембране В-лимфоцитов. Выполняют функции антигенспецифических рецепторов. Рецепторные
- 28. Нормальные антитела. В сыворотке крови человека всегда определяется базальный уровень иммуноглобулинов, которые получили название нормальных (естественных)
- 29. Моноклональные антитела. Каждый В-лимфоцит и его потомки, образовавшиеся в результате пролиферации (т. е. клон), способны синтезировать
- 30. Получение моноклональных антител возможно, если провести селекцию антителопродуцирующих клеток и их клонирование (т. е. выделение отдельных
- 31. Впервые моноклональные антитела были получены Д. Келлером и Ц. Мильштейном (1975). Они получили гибридные клетки путем
- 32. Полные и неполные антитела. По способности образовывать в реакции агглютинации или преципитации (in vitro) макромолекулярную структуру
- 33. Неполные (непреципитирующие) антитела лишены такой способности, несмотря на то что они специфически связываются с антигеном. Причиной
- 34. Другие виды антител. Различают тепловые и холодовые антитела. Первые взаимодействуют с антигеном при температуре +37 °С.
- 35. По способности активировать комплемент антитела подразделяются на комплементсвязывающие (IgM, IgGl и IgG3) и комплементнесвязывающие. Некоторые антитела
- 36. Получены белки со свойствами антител — это одноцепочечные антитела, бифункциональные антитела и иммунотоксины. Они синтезируются живыми
- 37. Иммунотоксины это гибриды иммуноглобулина и токсина. Они способны направленно доставить молекулу токсина к клетке-мишени. Иммунотоксины и
- 38. Иммуноглобулин обладает антигеностью и выраженной иммуногенностью. В молекуле Ig различают 4 типа атигенных детерминант: Видовые антигенные
- 39. Атотипические антигенные детерминанты индивидуальны. Они располагаются в легкой и тяжелой полипептидных цепях. На основании строения аллотипических
- 40. В процессе взаимодействия с антигеном принимает участие ограниченный участок Ig — антигенсвязывающии центр, или паратоп. Для
- 41. Связь антигена с антителом осуществляется за счет слабых взаимодействий (ван-дер-ваальсовы силы, водородные связи, электростатические взаимодействия) .
- 42. Особенности антитела - аффинность и авидность. Аффинность — сила специфического взаимодействия антитела с антигеном . Зависит
- 43. Авидность - это прочность связывания антитела и антигена. Эта характеристика определяется аффинностью Ig и числом антигенсвязывающих
- 45. Скачать презентацию
Слайд 2
Формы проявления иммунного ответа:
1) антителообразование,
2) иммунный фагоцитоз,
3) опосредованный клетками киллинг,
Формы проявления иммунного ответа:
1) антителообразование,
2) иммунный фагоцитоз,
3) опосредованный клетками киллинг,

Слайд 3 Все элементы иммунной системы реагируют на изменение гомеостаза, в зависимости от характера
Все элементы иммунной системы реагируют на изменение гомеостаза, в зависимости от характера

Слайд 4 Антитела — белки, специфически реагирующие с антигенами.
Антитела и фагоцитоз - одна
Антитела — белки, специфически реагирующие с антигенами.
Антитела и фагоцитоз - одна

Слайд 5 Антитела — это γ-глобулины, вырабатываемые в ответ на введение антигена, способные специфически
Антитела — это γ-глобулины, вырабатываемые в ответ на введение антигена, способные специфически

Слайд 6 Антитела используют для:
1. диагностики
2.профилактики
3.лечения различных заболеваний,
4.выделения
Антитела используют для:
1. диагностики
2.профилактики
3.лечения различных заболеваний,
4.выделения

Слайд 7Иммуноглобулины - это гликопротеиды. Они состоят из полипептидных цепей, стабилизированных сахаридными остатками.
Иммуноглобулины - это гликопротеиды. Они состоят из полипептидных цепей, стабилизированных сахаридными остатками.

Слайд 8Строение иммуноглобулина класса G
Строение иммуноглобулина класса G
Слайд 9Между тяжелыми цепями также есть дисульфидная связь . Такой тип межпептидного соединения
Между тяжелыми цепями также есть дисульфидная связь . Такой тип межпептидного соединения

Слайд 10 Отдельные участки цепи молекулы Ig свернуты в глобулы (домены). Домены стабилизированы
Отдельные участки цепи молекулы Ig свернуты в глобулы (домены). Домены стабилизированы

Слайд 11 Вариабельные домены легкой и тяжелой цепи образуют участок, который специфически связывается с
Вариабельные домены легкой и тяжелой цепи образуют участок, который специфически связывается с

Слайд 12 Обработка ферментами молекулы Ig приводит к ее гидролизу.
Папаин разрывает молекулу на
Обработка ферментами молекулы Ig приводит к ее гидролизу.
Папаин разрывает молекулу на

Слайд 13В зависимости от особенностей молекулярного строения тяжелой цепи различают 5 классов, или
В зависимости от особенностей молекулярного строения тяжелой цепи различают 5 классов, или

Слайд 14Иммуноглобулин класса G. Изотип G составляет основную массу Ig сыворотки крови
(
Иммуноглобулин класса G. Изотип G составляет основную массу Ig сыворотки крови
(

Слайд 15 IgGl и IgG3 связывают комплемент. IgG4, обладает цитофильностью (тропностью к тучным
IgGl и IgG3 связывают комплемент. IgG4, обладает цитофильностью (тропностью к тучным

Слайд 16Иммуноглобулин класса М. Самая крупная молекула из всех Ig. Это пентамер, который
Иммуноглобулин класса М. Самая крупная молекула из всех Ig. Это пентамер, который

Слайд 18IgM филогенетически наиболее древний иммуноглобулин. Синтезируется предшественниками и зрелыми В-лимфоцитами . Образуется
IgM филогенетически наиболее древний иммуноглобулин. Синтезируется предшественниками и зрелыми В-лимфоцитами . Образуется

Слайд 19Не проходит через плаценту. Обнаружение специфических антител изотипа М в сыворотке крови
Не проходит через плаценту. Обнаружение специфических антител изотипа М в сыворотке крови

Слайд 20Иммуноглобулин класса А. Существует в сывороточной и секреторной формах. Около 60 %
Иммуноглобулин класса А. Существует в сывороточной и секреторной формах. Около 60 %

Слайд 22 Может быть неполным антителом. Не связывает комплемент. Не проходит через плацентарный
Может быть неполным антителом. Не связывает комплемент. Не проходит через плацентарный

Слайд 23 Молекулярная масса 350 кДа и выше, константа седиментации 13S и выше.
Синтезируется В-лимфоцитами
Молекулярная масса 350 кДа и выше, константа седиментации 13S и выше.
Синтезируется В-лимфоцитами

Слайд 24
Секреторная форма IgA — основной фактор специфического гуморального местного иммунитета слизистых оболочек
Секреторная форма IgA — основной фактор специфического гуморального местного иммунитета слизистых оболочек

Слайд 25 Иммуноглобулин класса Е ( реагин). Содержание в сыворотке крови — 0,00025 г/л.
Иммуноглобулин класса Е ( реагин). Содержание в сыворотке крови — 0,00025 г/л.

Слайд 26Иммуноглобулин класса D. Концентрация в сыворотке крови - 0,03 г/л (около 0,2
Иммуноглобулин класса D. Концентрация в сыворотке крови - 0,03 г/л (около 0,2

Слайд 27 Рецепторные иммуноглобулины. Рецепторные, или мембраные Ig, локализуются на цитоплазматической мембране В-лимфоцитов. Выполняют
Рецепторные иммуноглобулины. Рецепторные, или мембраные Ig, локализуются на цитоплазматической мембране В-лимфоцитов. Выполняют

Слайд 28 Нормальные антитела. В сыворотке крови человека всегда определяется базальный уровень иммуноглобулинов, которые
Нормальные антитела. В сыворотке крови человека всегда определяется базальный уровень иммуноглобулинов, которые

Слайд 29 Моноклональные антитела. Каждый В-лимфоцит и его потомки, образовавшиеся в результате пролиферации (т.
Моноклональные антитела. Каждый В-лимфоцит и его потомки, образовавшиеся в результате пролиферации (т.

Слайд 30 Получение моноклональных антител возможно, если провести селекцию антителопродуцирующих клеток и их клонирование
Получение моноклональных антител возможно, если провести селекцию антителопродуцирующих клеток и их клонирование

Слайд 31Впервые моноклональные антитела были получены Д. Келлером и Ц. Мильштейном (1975). Они
Впервые моноклональные антитела были получены Д. Келлером и Ц. Мильштейном (1975). Они

Слайд 32 Полные и неполные антитела.
По способности образовывать в реакции агглютинации или преципитации
Полные и неполные антитела.
По способности образовывать в реакции агглютинации или преципитации

Слайд 33 Неполные (непреципитирующие) антитела лишены такой способности, несмотря на то что они специфически
Неполные (непреципитирующие) антитела лишены такой способности, несмотря на то что они специфически

Слайд 34Другие виды антител. Различают тепловые и холодовые антитела. Первые взаимодействуют с антигеном
Другие виды антител. Различают тепловые и холодовые антитела. Первые взаимодействуют с антигеном

Слайд 35По способности активировать комплемент антитела подразделяются на комплементсвязывающие (IgM, IgGl и IgG3)
По способности активировать комплемент антитела подразделяются на комплементсвязывающие (IgM, IgGl и IgG3)

Слайд 36Получены белки со свойствами антител — это одноцепочечные антитела, бифункциональные антитела и
Получены белки со свойствами антител — это одноцепочечные антитела, бифункциональные антитела и

Слайд 37 Иммунотоксины это гибриды иммуноглобулина и токсина. Они способны направленно доставить молекулу токсина
Иммунотоксины это гибриды иммуноглобулина и токсина. Они способны направленно доставить молекулу токсина

Слайд 38 Иммуноглобулин обладает антигеностью и выраженной иммуногенностью. В молекуле Ig различают 4 типа
Иммуноглобулин обладает антигеностью и выраженной иммуногенностью. В молекуле Ig различают 4 типа

Слайд 39Атотипические антигенные детерминанты индивидуальны. Они располагаются в легкой и тяжелой полипептидных цепях.
Атотипические антигенные детерминанты индивидуальны. Они располагаются в легкой и тяжелой полипептидных цепях.

Слайд 40В процессе взаимодействия с антигеном принимает участие ограниченный участок Ig — антигенсвязывающии
В процессе взаимодействия с антигеном принимает участие ограниченный участок Ig — антигенсвязывающии

Слайд 41 Связь антигена с антителом осуществляется за счет слабых взаимодействий (ван-дер-ваальсовы силы, водородные
Связь антигена с антителом осуществляется за счет слабых взаимодействий (ван-дер-ваальсовы силы, водородные

Слайд 42
Особенности антитела - аффинность и авидность.
Аффинность — сила специфического взаимодействия антитела с
Особенности антитела - аффинность и авидность.
Аффинность — сила специфического взаимодействия антитела с

Слайд 43Авидность - это прочность связывания антитела и антигена. Эта характеристика определяется аффинностью
Авидность - это прочность связывания антитела и антигена. Эта характеристика определяется аффинностью

 Удалите лишнее
Удалите лишнее 45b93ad8b40a432b9fbc7c499afbbc6c (1)
45b93ad8b40a432b9fbc7c499afbbc6c (1) Цифровые приборы
Цифровые приборы Система образования в Канаде
Система образования в Канаде Путешествие в зимнюю Москву!
Путешествие в зимнюю Москву! Презентация на тему Образ святого Александра Невского в культуре и литературе
Презентация на тему Образ святого Александра Невского в культуре и литературе  Коммерческое Предложение
Коммерческое Предложение Организм человека как единая саморазвивающаяся биологическая система (Лекция 4)
Организм человека как единая саморазвивающаяся биологическая система (Лекция 4) Реклама в России.События и перспективы
Реклама в России.События и перспективы Возникновение искусства и религии
Возникновение искусства и религии Презентация на тему Питание растений
Презентация на тему Питание растений  М.Е.Салтыков-Щедрин «История одного города» Власть и мы
М.Е.Салтыков-Щедрин «История одного города» Власть и мы Международные стандарты ISO
Международные стандарты ISO География Австралии!
География Австралии! Денежные агрегаты
Денежные агрегаты Саудовской Аравии
Саудовской Аравии Муниципальное общеобразовательное учреждение «Заводская средняя общеобразовательная школа» Троицкого района
Муниципальное общеобразовательное учреждение «Заводская средняя общеобразовательная школа» Троицкого района Центр развития (www.dcenter.ru)
Центр развития (www.dcenter.ru) Механизм протекания цепной ядерной реакции
Механизм протекания цепной ядерной реакции Общая характеристика семейства Эльбрус. Разработки на базе современной отечественной микропроцессорной архитектуры
Общая характеристика семейства Эльбрус. Разработки на базе современной отечественной микропроцессорной архитектуры Сообщество «Сделаем вместе!»
Сообщество «Сделаем вместе!» Презентация на тему Малые жанры фольклора (5 класс)
Презентация на тему Малые жанры фольклора (5 класс) Уроки с использованием компьютерных технологий в учебном процессе
Уроки с использованием компьютерных технологий в учебном процессе Мотив крика, одиночества в живописи и литературе
Мотив крика, одиночества в живописи и литературе Мультимедийные урокиМАСТЕР-КЛАСС
Мультимедийные урокиМАСТЕР-КЛАСС 2CV – Rencontre mondiale
2CV – Rencontre mondiale Иоганн генрих песталоцци
Иоганн генрих песталоцци ПЛАТЕЖНЫЕ КАРТЫ:реальное состояние и нереализованные возможности
ПЛАТЕЖНЫЕ КАРТЫ:реальное состояние и нереализованные возможности