- Генно-модифицированные продукты растительного происхождения: проблемы биоэтики и биобезопасности

Содержание
- 2. Что же такое ГМО? Генети́чески модифици́рованный органи́зм (ГМО) — живой организм, генотип которого был искусственно изменён
- 3. Глоссарий Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК,
- 4. Общая оценка состояния генной инженерии Научно-технический прогресс в жизни современного общества ознаменовался появлением новых научных и
- 5. Бурное развитие технологий и быстрое внедрение в практику научных достижений зачастую не подкреплено достаточно обоснованными оценками
- 6. Использование ГМО в медицинских целях Генно-инженерные организмы используются в прикладной медицине с 1982 года, когда был
- 7. Использование ГМО в сельском хозяйстве Генная инженерия используется для создания новых сортов растений, устойчивых к неблагоприятным
- 14. Другие направления использования ГМО Разрабатываются генно-инженерные бактерии, способные производить экологически чистое топливо. В 2003 году на
- 15. Где чаще всего встречаются генетически модифицированные организмы?
- 16. Проведем научный анализ рисков, связанных с широким использованием генетически модифицированных (трансгенных) организмов (ГМО) и продуктов их
- 17. Биотехнология и генная инженерия Основой современной биотехнологии является генная, или генетическая инженерия – совокупность приемов, методов
- 18. Биотехнология и генная инженерия Технология рекомбинантных ДНК использует следующие методы: специфическое расщепление ДНК рестрицирующими нуклеазами, ускоряющее
- 19. Биотехнология и генная инженерия Методом генной инженерии уже получен ряд препаратов медицинского назначения, в том числе
- 20. Биотехнология и генная инженерия Начиная с 1982 г. фирмы США, Японии, Великобритании и других стран производят
- 21. Лиофилизированный препарат рекомбинантного нуклеокапсидного белка вируса гепатита С обладает антигенными и иммуногенными свойствами будет использован в
- 22. Биотехнология и генная инженерия Технология рекомбинантных ДНК сделала возможным нетрадиционный подход "белок-ген", получивший название "обратная генетика".
- 23. Биотехнология и генная инженерия Если гибридную ДНК ввести в оплодотворенное яйцеклетку, могут быть получены трансгенные организмы,
- 24. Векторные системы для введения генетической информации в растительные клетки Подробнее о введении ДНК в клетки растений
- 25. Векторные системы для введения генетической информации в растительные клетки Генетическая колонизация растения A. tumefaciens: 1- агробактерии
- 26. Векторные системы для введения генетической информации в растительные клетки Ткани корончатых галлов содержат более высокие уровни
- 27. Векторные системы для введения генетической информации в растительные клетки Структура Тi-плазмид нопалинового и окто-пинового типа Подробная
- 28. Векторные системы для введения генетической информации в растительные клетки Кроме Т-ДНК в плазмидах имеются область, кодирующая
- 29. Векторные системы для введения генетической информации в растительные клетки Agrobacterium имеет очень широкий круг растений-хозяев и
- 30. Векторные системы для введения генетической информации в растительные клетки Коинтегративные и бинарные вектора Разработаны два метода
- 31. Векторные системы для введения генетической информации в растительные клетки Т-ДНК вырезают из Ti-плазмиды с помощью рестриктаз
- 32. Векторные системы для введения генетической информации в растительные клетки Второй метод основан на создании системы trans-,
- 33. Векторные системы для введения генетической информации в растительные клетки Пример плазмидного бинарного вектора. Вектор pMON10117 использован
- 34. Векторные системы для введения генетической информации в растительные клетки После клонирования, изучения и отбора нужных нам
- 35. Векторные системы для введения генетической информации в растительные клетки При разработке бинарных векторных систем использована такая
- 36. Современные направления в создании генетически модифицированных растений Первые трансгенные растения были получены с помощью технологии рекомбинантной
- 37. Современные направления в создании генетически модифицированных растений Генетическая конструкция, вводимая в растительную клетку обычно включает: белоккодирующую
- 38. Современные направления в создании генетически модифицированных растений Минимальный промотор, связывающий РНК-полимеразу, как правило, недостаточен для обеспечения
- 39. Современные направления в создании генетически модифицированных растений Помимо конститутивных, известно большое число специфических промоторов, которые активны
- 40. Современные направления в создании генетически модифицированных растений Ввести чужеродную ДНК в растения можно различными способами. Для
- 41. Современные направления в создании генетически модифицированных растений На первом этапе осуществляется выделение трансгена из геномной ДНК
- 42. Современные направления в создании генетически модифицированных растений Агробактериальная трансформация — наиболее эффективная технология введения трансгенов в
- 43. Современные направления в создании генетически модифицированных растений Для трансформации устойчивых ("рекальцитрантных") к агробактериям растений разработаны приемы
- 44. Современные направления в создании генетически модифицированных растений Баллистическая трансфекция основана на обстреле органов и тканей микрочастицами
- 45. Современные направления в создании генетически модифицированных растений Метод введения ДНК в клетки растений с помощью биолистики
- 46. Современные направления в создании генетически модифицированных растений В последнее время был разработан и успешно применен также
- 47. Современные направления в создании генетически модифицированных растений Возможности генной инженерии растений: Улучшение качества запасных белков, в
- 48. Социально-экономические аспекты внедрения трансгенных организмов в практику
- 49. Социально-экономические аспекты внедрения трансгенных организмов в практику
- 50. Доля сои в посевах – 55%, хлопка – 21%, рапса – 16%, кукурузы – 11%
- 51. Тенденции по использованию ГМ растений в Европе Небольшие посевные площади. Еврокомиссия одобрила посевы 17 сортов кукурузы,
- 52. Трансгенный картофель, несущий ген хитиназы Ингибирование роста патогена Fusarium oxysporum экстрактом трансгенного картофеля (В). А –
- 53. Трансгенные растения табака с введенным геном цитохрома Р450scc (CYP11A1) 1 – дикий тип, 2 – контрольное
- 54. Внешний вид контрольных и трансформированных растений Nicotiana plumbaginifolia с химерным геном апоэкворина К T0 T0 T0
- 55. Критерии и методы оценки безопасности генетически модифицированных организмов Исследование безопасности ГМО (правильнее говорить именно генно-инженерные организмы!)
- 56. Критерии и методы оценки безопасности генетически модифицированных организмов
- 57. Критерии и методы оценки безопасности генетически модифицированных организмов Риск и оценка риска. По наиболее общему определению,
- 58. Критерии и методы оценки безопасности генетически модифицированных организмов Риск генно-инженерной деятельности. Для получения экономической выгоды от
- 59. Критерии и методы оценки безопасности генетически модифицированных организмов В конечном итоге процедура оценки риска должна дать
- 60. Критерии и методы оценки безопасности генетически модифицированных организмов Принцип принятия мер предосторожности Источники появления и применения
- 61. Критерии и методы оценки безопасности генетически модифицированных организмов Принцип принятия мер предосторожности является по существу политической
- 62. Критерии и методы оценки безопасности генетически модифицированных организмов С целью прояснить порядок применения данного принципа в
- 63. Критерии и методы оценки безопасности генетически модифицированных организмов Пропорциональность соответствия. Меры по управлению риском ГИД в
- 64. Критерии и методы оценки безопасности генетически модифицированных организмов Компоненты идеальной системы оценки риска
- 65. Критерии и методы оценки безопасности генетически модифицированных организмов Применяемая в разных странах методика оценки риска ГИД
- 66. Критерии и методы оценки безопасности генетически модифицированных организмов • Оценка возможных последствий каждого неблагоприятного воздействия ГИД,
- 67. Международная и государственная регламентация биобезопасности Если экономическая выгода от использования ГИО в целом очевидна, то их
- 68. Международная и государственная регламентация биобезопасности Во избежание негативных последствий от бесконтрольного применения генетически модифицированных организмов, 29
- 69. Международная и государственная регламентация биобезопасности Наибольшее беспокойство вызывает вероятность переноса генетического материала трансгенного растения в геномы
- 70. Международная и государственная регламентация биобезопасности Так, в 2003 г. немецкие исследователи из Кельна и Эрлангена опубликовали
- 71. Международная и государственная регламентация биобезопасности Международная структура биобезопасности и структура биобезопасности отдельных государств включают в себя
- 72. Международная и государственная регламентация биобезопасности Национальный координа-ционный центр био-безопасности организован на базе Института генетики и цитологии
- 73. Международная и государственная регламентация биобезопасности
- 74. Международная и государственная регламентация биобезопасности Среди прочих информационных материалов на сайте центра представлены различные национальные и
- 75. вступил в силу 10 июля 2006 г. 27 нормативных правовых актов законодательства Изменения в законе Республики
- 77. Скачать презентацию
Слайд 2Что же такое ГМО?
Генети́чески модифици́рованный органи́зм (ГМО) — живой организм, генотип которого
Что же такое ГМО?
Генети́чески модифици́рованный органи́зм (ГМО) — живой организм, генотип которого

Слайд 3Глоссарий
Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных
Глоссарий
Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных

Слайд 4Общая оценка состояния генной инженерии
Научно-технический прогресс в жизни современного общества ознаменовался
Общая оценка состояния генной инженерии
Научно-технический прогресс в жизни современного общества ознаменовался

Слайд 5Бурное развитие технологий и быстрое внедрение в практику научных достижений зачастую не
Бурное развитие технологий и быстрое внедрение в практику научных достижений зачастую не

Слайд 6Использование ГМО в медицинских целях
Генно-инженерные организмы используются в прикладной медицине с
Использование ГМО в медицинских целях
Генно-инженерные организмы используются в прикладной медицине с

Слайд 7Использование ГМО в сельском хозяйстве
Генная инженерия используется для создания новых сортов
Использование ГМО в сельском хозяйстве
Генная инженерия используется для создания новых сортов

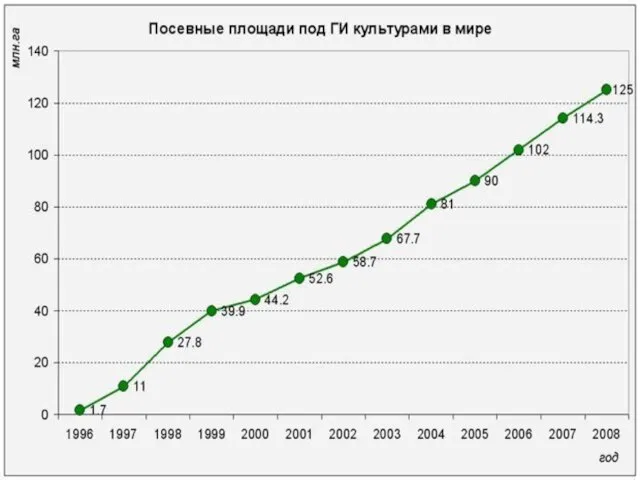
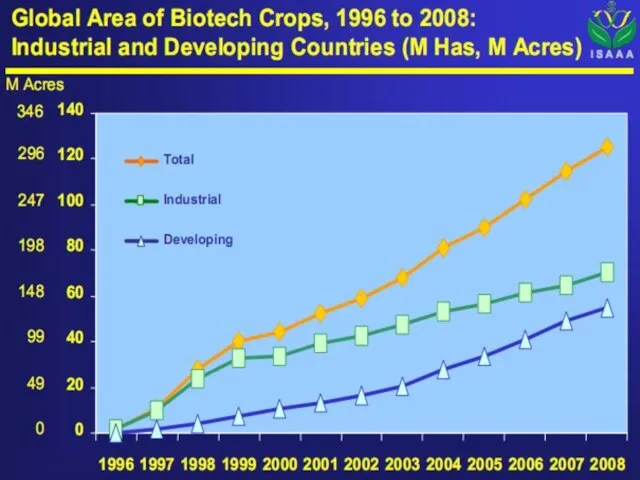
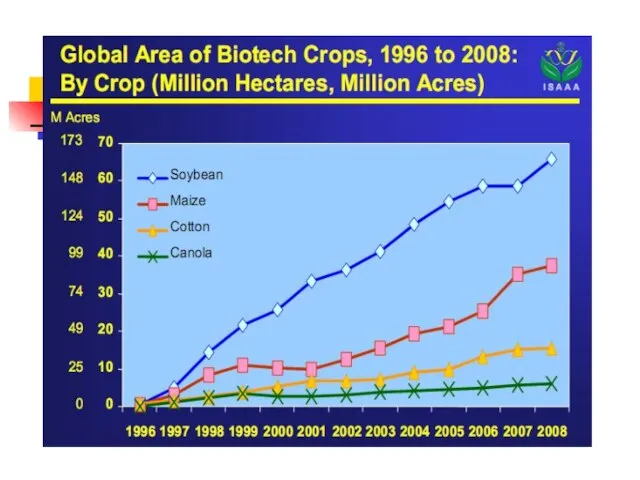
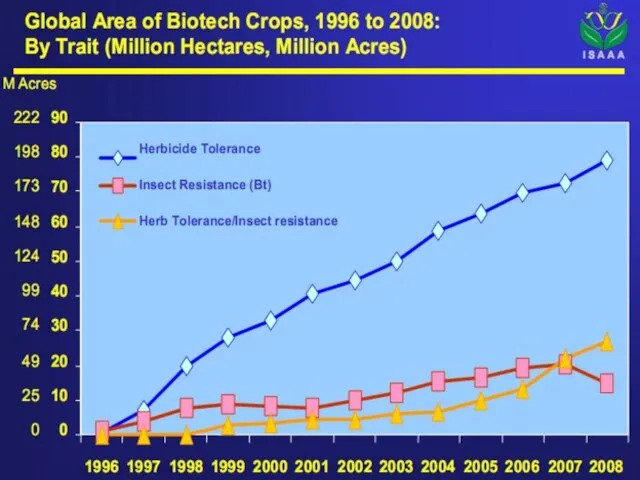
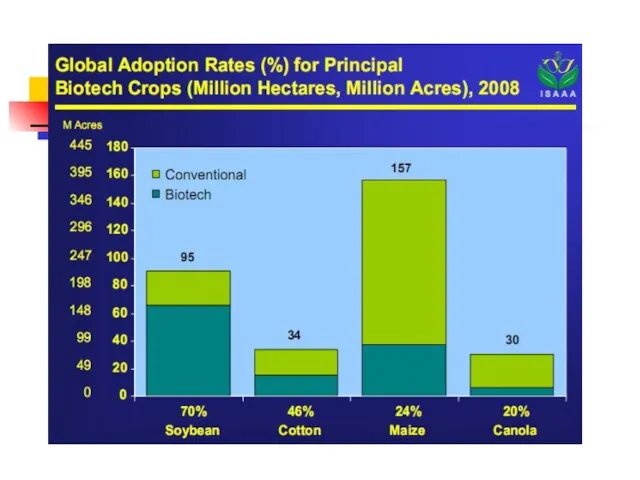
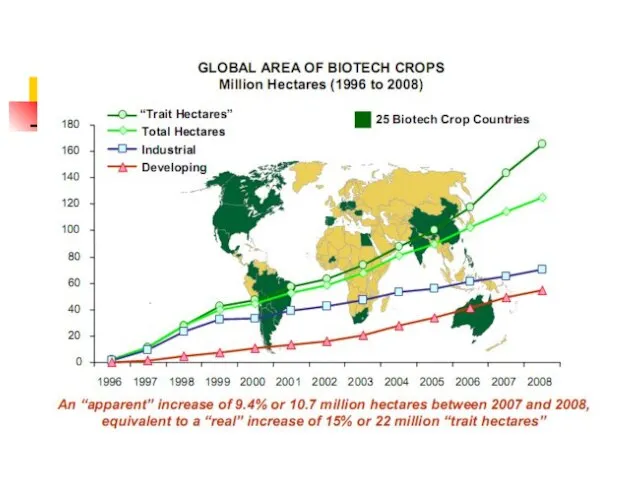
Слайд 14Другие направления
использования ГМО
Разрабатываются генно-инженерные бактерии, способные производить экологически чистое топливо.
В 2003
Другие направления
использования ГМО
Разрабатываются генно-инженерные бактерии, способные производить экологически чистое топливо.
В 2003

Слайд 15Где чаще всего встречаются генетически модифицированные организмы?
Где чаще всего встречаются генетически модифицированные организмы?

Слайд 16Проведем научный анализ рисков, связанных с широким использованием генетически модифицированных (трансгенных) организмов
Проведем научный анализ рисков, связанных с широким использованием генетически модифицированных (трансгенных) организмов

Слайд 17Биотехнология и генная инженерия
Основой современной биотехнологии является генная, или генетическая инженерия –
Биотехнология и генная инженерия
Основой современной биотехнологии является генная, или генетическая инженерия –

Слайд 18Биотехнология и генная инженерия
Технология рекомбинантных ДНК использует следующие методы:
специфическое расщепление
Биотехнология и генная инженерия
Технология рекомбинантных ДНК использует следующие методы:
специфическое расщепление

Слайд 19Биотехнология и генная инженерия
Методом генной инженерии уже получен ряд препаратов медицинского назначения,
Биотехнология и генная инженерия
Методом генной инженерии уже получен ряд препаратов медицинского назначения,

Слайд 20Биотехнология и генная инженерия
Начиная с 1982 г. фирмы США, Японии, Великобритании и
Биотехнология и генная инженерия
Начиная с 1982 г. фирмы США, Японии, Великобритании и

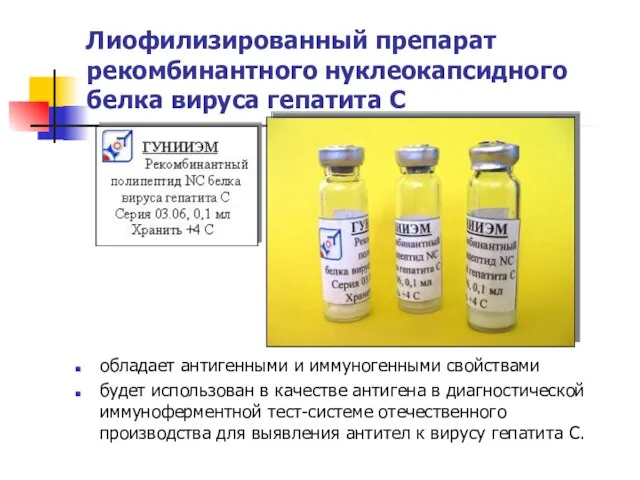
Слайд 21Лиофилизированный препарат рекомбинантного нуклеокапсидного белка вируса гепатита С
обладает антигенными и иммуногенными свойствами
будет
Лиофилизированный препарат рекомбинантного нуклеокапсидного белка вируса гепатита С
обладает антигенными и иммуногенными свойствами
будет
Слайд 22Биотехнология и генная инженерия
Технология рекомбинантных ДНК сделала возможным нетрадиционный подход "белок-ген", получивший
Биотехнология и генная инженерия
Технология рекомбинантных ДНК сделала возможным нетрадиционный подход "белок-ген", получивший

Слайд 23Биотехнология и генная инженерия
Если гибридную ДНК ввести в оплодотворенное яйцеклетку, могут быть
Биотехнология и генная инженерия
Если гибридную ДНК ввести в оплодотворенное яйцеклетку, могут быть

Слайд 24Векторные системы для введения генетической информации в растительные клетки
Подробнее о введении ДНК
Векторные системы для введения генетической информации в растительные клетки
Подробнее о введении ДНК

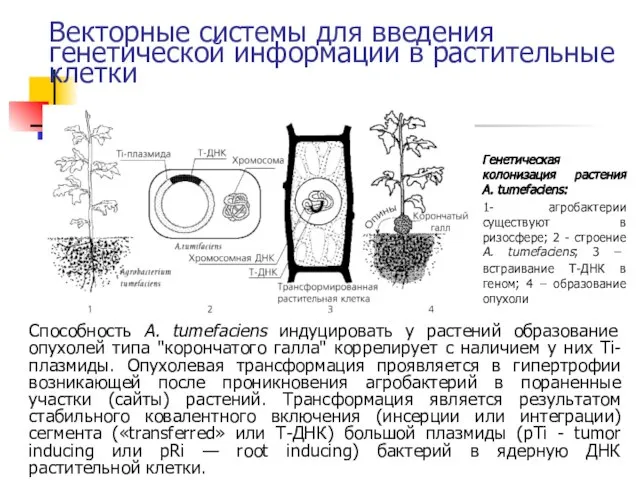
Слайд 25Векторные системы для введения генетической информации в растительные клетки
Генетическая колонизация растения A.
Векторные системы для введения генетической информации в растительные клетки
Генетическая колонизация растения A.
Слайд 26Векторные системы для введения генетической информации в растительные клетки
Ткани корончатых галлов содержат
Векторные системы для введения генетической информации в растительные клетки
Ткани корончатых галлов содержат

Слайд 27Векторные системы для введения генетической информации в растительные клетки
Структура Тi-плазмид нопалинового и
Векторные системы для введения генетической информации в растительные клетки
Структура Тi-плазмид нопалинового и
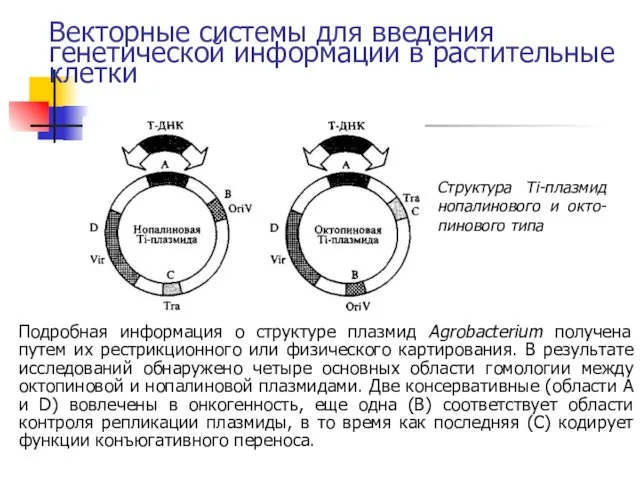
Слайд 28Векторные системы для введения генетической информации в растительные клетки
Кроме Т-ДНК в плазмидах
Векторные системы для введения генетической информации в растительные клетки
Кроме Т-ДНК в плазмидах

Слайд 29Векторные системы для введения генетической информации в растительные клетки
Agrobacterium имеет очень широкий
Векторные системы для введения генетической информации в растительные клетки
Agrobacterium имеет очень широкий

Слайд 30Векторные системы для введения генетической информации в растительные клетки
Коинтегративные и бинарные вектора
Векторные системы для введения генетической информации в растительные клетки
Коинтегративные и бинарные вектора
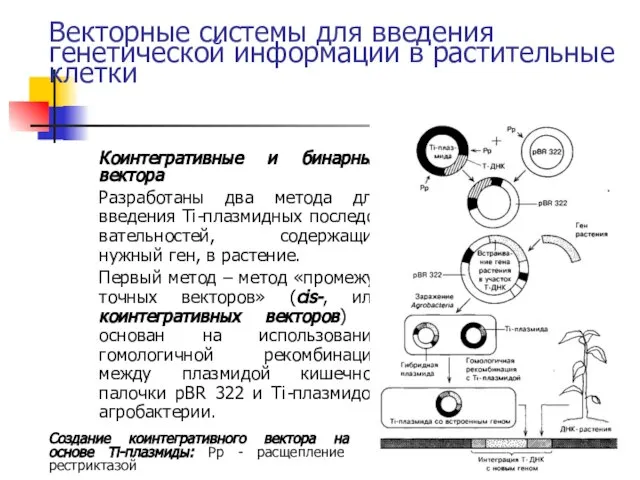
Слайд 31Векторные системы для введения генетической информации в растительные клетки
Т-ДНК вырезают из Ti-плазмиды
Векторные системы для введения генетической информации в растительные клетки
Т-ДНК вырезают из Ti-плазмиды

Слайд 32Векторные системы для введения генетической информации в растительные клетки
Второй метод основан на
Векторные системы для введения генетической информации в растительные клетки
Второй метод основан на

Слайд 33Векторные системы для введения генетической информации в растительные клетки
Пример плазмидного бинарного вектора.
Векторные системы для введения генетической информации в растительные клетки
Пример плазмидного бинарного вектора.
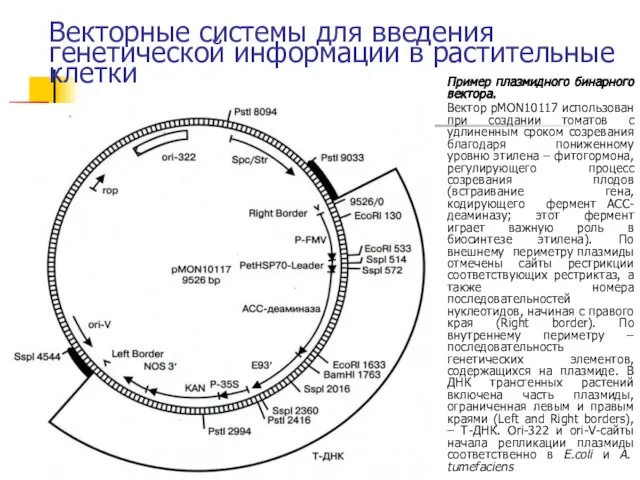
Слайд 34Векторные системы для введения генетической информации в растительные клетки
После клонирования, изучения и
Векторные системы для введения генетической информации в растительные клетки
После клонирования, изучения и

Слайд 35Векторные системы для введения генетической информации в растительные клетки
При разработке бинарных векторных
Векторные системы для введения генетической информации в растительные клетки
При разработке бинарных векторных

Слайд 36Современные направления в создании генетически модифицированных растений
Первые трансгенные растения были получены с
Современные направления в создании генетически модифицированных растений
Первые трансгенные растения были получены с

Слайд 37Современные направления в создании генетически модифицированных растений
Генетическая конструкция, вводимая в растительную клетку
Современные направления в создании генетически модифицированных растений
Генетическая конструкция, вводимая в растительную клетку

Слайд 38Современные направления в создании генетически модифицированных растений
Минимальный промотор, связывающий РНК-полимеразу, как правило,
Современные направления в создании генетически модифицированных растений
Минимальный промотор, связывающий РНК-полимеразу, как правило,

Слайд 39Современные направления в создании генетически модифицированных растений
Помимо конститутивных, известно большое число специфических
Современные направления в создании генетически модифицированных растений
Помимо конститутивных, известно большое число специфических

Слайд 40Современные направления в создании генетически модифицированных растений
Ввести чужеродную ДНК в растения можно
Современные направления в создании генетически модифицированных растений
Ввести чужеродную ДНК в растения можно

Слайд 41Современные направления в создании генетически модифицированных растений
На первом этапе осуществляется выделение трансгена
Современные направления в создании генетически модифицированных растений
На первом этапе осуществляется выделение трансгена

Слайд 42Современные направления в создании генетически модифицированных растений
Агробактериальная трансформация — наиболее эффективная технология
Современные направления в создании генетически модифицированных растений
Агробактериальная трансформация — наиболее эффективная технология

Слайд 43Современные направления в создании генетически модифицированных растений
Для трансформации устойчивых ("рекальцитрантных") к агробактериям
Современные направления в создании генетически модифицированных растений
Для трансформации устойчивых ("рекальцитрантных") к агробактериям

Слайд 44Современные направления в создании генетически модифицированных растений
Баллистическая трансфекция основана на обстреле органов
Современные направления в создании генетически модифицированных растений
Баллистическая трансфекция основана на обстреле органов

Слайд 45Современные направления в создании генетически модифицированных растений
Метод введения ДНК в клетки растений
Современные направления в создании генетически модифицированных растений
Метод введения ДНК в клетки растений
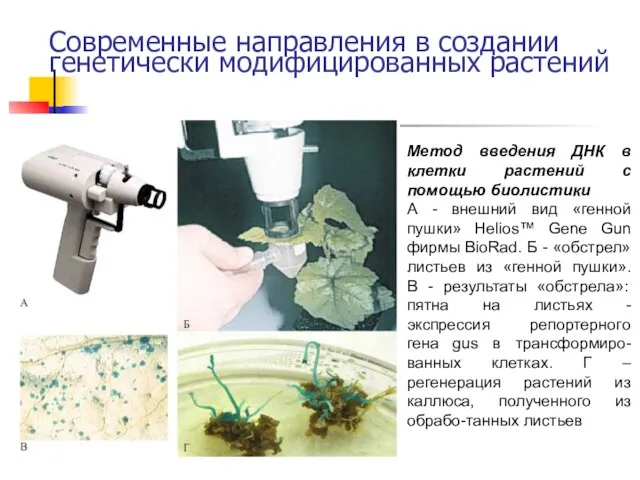
Слайд 46Современные направления в создании генетически модифицированных растений
В последнее время был разработан и
Современные направления в создании генетически модифицированных растений
В последнее время был разработан и

Слайд 47Современные направления в создании генетически модифицированных растений
Возможности генной инженерии растений:
Улучшение качества запасных
Современные направления в создании генетически модифицированных растений
Возможности генной инженерии растений:
Улучшение качества запасных

Слайд 48Социально-экономические аспекты внедрения трансгенных организмов в практику
Социально-экономические аспекты внедрения трансгенных организмов в практику
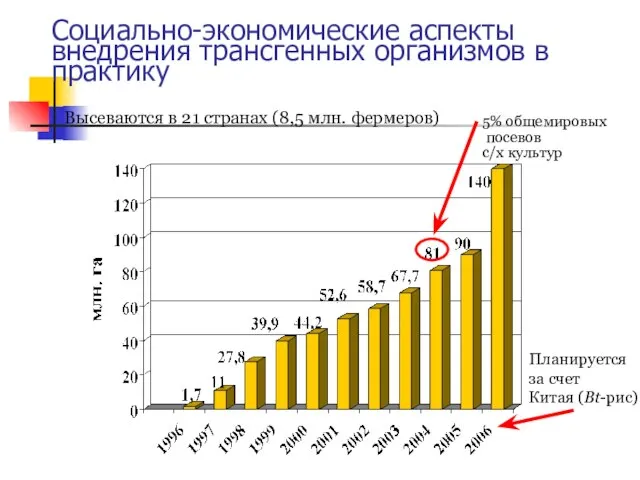
Слайд 49Социально-экономические аспекты внедрения трансгенных организмов в практику
Социально-экономические аспекты внедрения трансгенных организмов в практику

Слайд 50Доля сои в посевах – 55%, хлопка – 21%, рапса – 16%,
Доля сои в посевах – 55%, хлопка – 21%, рапса – 16%,
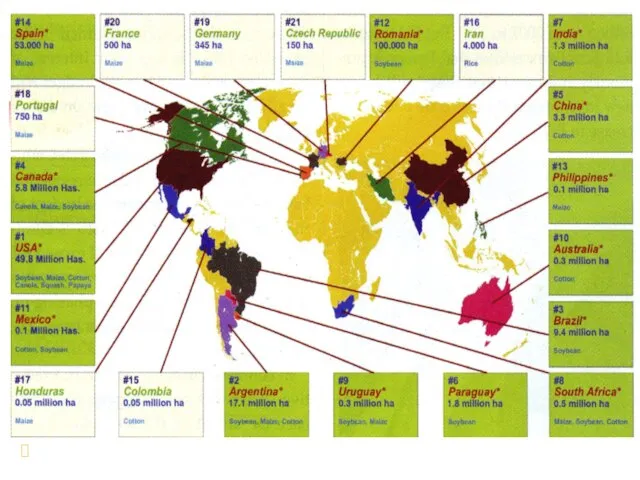
Слайд 51Тенденции по использованию ГМ растений в Европе
Небольшие посевные площади.
Еврокомиссия одобрила
Тенденции по использованию ГМ растений в Европе
Небольшие посевные площади.
Еврокомиссия одобрила

Слайд 52Трансгенный картофель, несущий ген хитиназы
Ингибирование роста патогена Fusarium oxysporum экстрактом трансгенного картофеля
Трансгенный картофель, несущий ген хитиназы
Ингибирование роста патогена Fusarium oxysporum экстрактом трансгенного картофеля

Слайд 53Трансгенные растения табака с введенным геном
цитохрома Р450scc (CYP11A1)
1 – дикий тип,
Трансгенные растения табака с введенным геном
цитохрома Р450scc (CYP11A1)
1 – дикий тип,

Слайд 54Внешний вид контрольных и трансформированных растений Nicotiana plumbaginifolia с химерным геном апоэкворина
К
T0
T0
T0
Внешний вид контрольных и трансформированных растений Nicotiana plumbaginifolia с химерным геном апоэкворина
К
T0
T0
T0

Слайд 55Критерии и методы оценки безопасности генетически модифицированных организмов
Исследование безопасности ГМО (правильнее говорить
Критерии и методы оценки безопасности генетически модифицированных организмов
Исследование безопасности ГМО (правильнее говорить

Слайд 56Критерии и методы оценки безопасности генетически модифицированных организмов
Критерии и методы оценки безопасности генетически модифицированных организмов

Слайд 57Критерии и методы оценки безопасности генетически модифицированных организмов
Риск и оценка риска. По
Критерии и методы оценки безопасности генетически модифицированных организмов
Риск и оценка риска. По

Слайд 58Критерии и методы оценки безопасности генетически модифицированных организмов
Риск генно-инженерной деятельности. Для получения
Критерии и методы оценки безопасности генетически модифицированных организмов
Риск генно-инженерной деятельности. Для получения

Слайд 59Критерии и методы оценки безопасности генетически модифицированных организмов
В конечном итоге процедура оценки
Критерии и методы оценки безопасности генетически модифицированных организмов
В конечном итоге процедура оценки

Слайд 60Критерии и методы оценки безопасности генетически модифицированных организмов
Принцип принятия мер предосторожности
Источники появления
Критерии и методы оценки безопасности генетически модифицированных организмов
Принцип принятия мер предосторожности
Источники появления

Слайд 61Критерии и методы оценки безопасности генетически модифицированных организмов
Принцип принятия мер предосторожности является
Критерии и методы оценки безопасности генетически модифицированных организмов
Принцип принятия мер предосторожности является

Слайд 62Критерии и методы оценки безопасности генетически модифицированных организмов
С целью прояснить порядок применения
Критерии и методы оценки безопасности генетически модифицированных организмов
С целью прояснить порядок применения

Слайд 63Критерии и методы оценки безопасности генетически модифицированных организмов
Пропорциональность соответствия. Меры по управлению
Критерии и методы оценки безопасности генетически модифицированных организмов
Пропорциональность соответствия. Меры по управлению

Слайд 64Критерии и методы оценки безопасности генетически модифицированных организмов
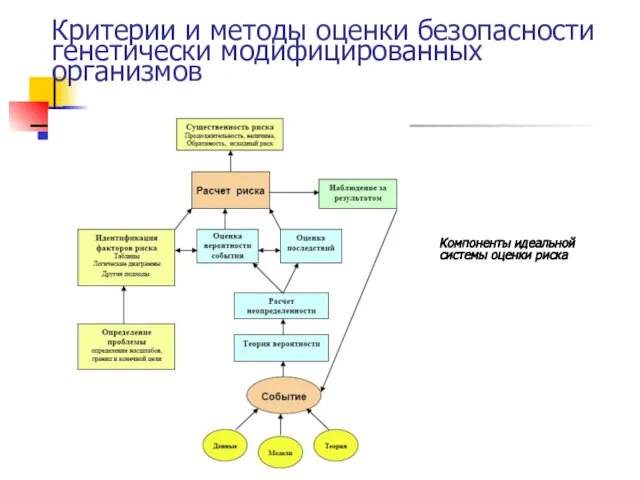
Компоненты идеальной системы оценки риска
Критерии и методы оценки безопасности генетически модифицированных организмов
Компоненты идеальной системы оценки риска
Слайд 65Критерии и методы оценки безопасности генетически модифицированных организмов
Применяемая в разных странах методика
Критерии и методы оценки безопасности генетически модифицированных организмов
Применяемая в разных странах методика

Слайд 66Критерии и методы оценки безопасности генетически модифицированных организмов
• Оценка возможных последствий каждого
Критерии и методы оценки безопасности генетически модифицированных организмов
• Оценка возможных последствий каждого

Слайд 67Международная и государственная регламентация биобезопасности
Если экономическая выгода от использования ГИО в
Международная и государственная регламентация биобезопасности
Если экономическая выгода от использования ГИО в

Слайд 68Международная и государственная регламентация биобезопасности
Во избежание негативных последствий от бесконтрольного применения
Международная и государственная регламентация биобезопасности
Во избежание негативных последствий от бесконтрольного применения

Слайд 69Международная и государственная регламентация биобезопасности
Наибольшее беспокойство вызывает вероятность переноса генетического материала
Международная и государственная регламентация биобезопасности
Наибольшее беспокойство вызывает вероятность переноса генетического материала

Слайд 70Международная и государственная регламентация биобезопасности
Так, в 2003 г. немецкие исследователи из
Международная и государственная регламентация биобезопасности
Так, в 2003 г. немецкие исследователи из

Слайд 71Международная и государственная регламентация биобезопасности
Международная структура биобезопасности и структура биобезопасности отдельных
Международная и государственная регламентация биобезопасности
Международная структура биобезопасности и структура биобезопасности отдельных

Слайд 72Международная и государственная регламентация биобезопасности
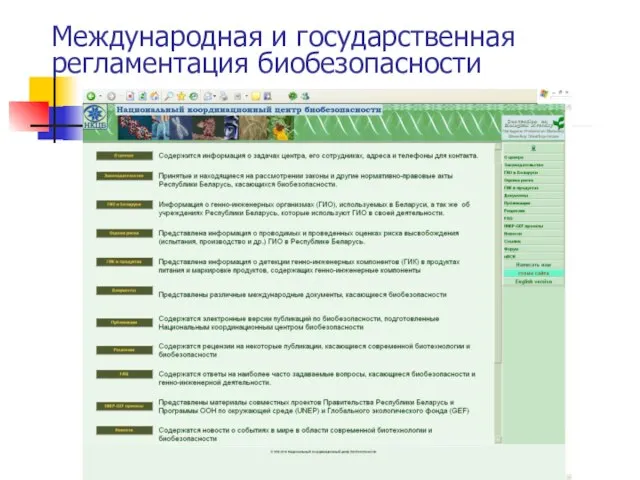
Национальный координа-ционный центр био-безопасности организован на базе
Международная и государственная регламентация биобезопасности
Национальный координа-ционный центр био-безопасности организован на базе
Слайд 73Международная и государственная регламентация биобезопасности
Международная и государственная регламентация биобезопасности
Слайд 74Международная и государственная регламентация биобезопасности
Среди прочих информационных материалов на сайте центра
Международная и государственная регламентация биобезопасности
Среди прочих информационных материалов на сайте центра

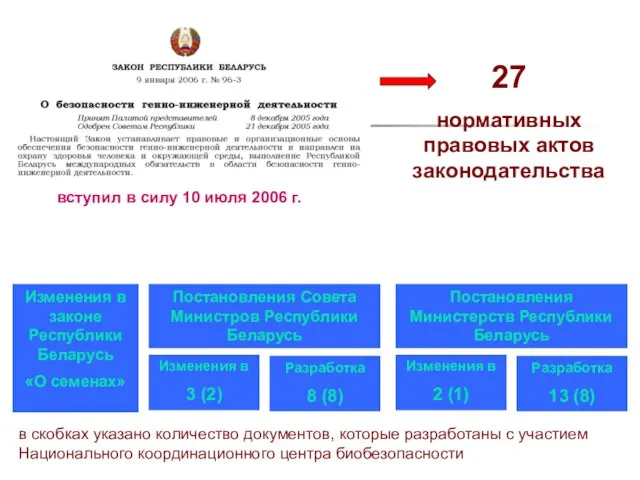
Слайд 75вступил в силу 10 июля 2006 г.
27
нормативных правовых актов законодательства
Изменения в
вступил в силу 10 июля 2006 г.
27
нормативных правовых актов законодательства
Изменения в
 Машиностроительный комплекс России
Машиностроительный комплекс России Уход в облака на практике
Уход в облака на практике Дорогие сердцу места
Дорогие сердцу места Презентация на тему Нумерации разных народов и их возникновение
Презентация на тему Нумерации разных народов и их возникновение  Экстремальное программированиеОпыт внедрения
Экстремальное программированиеОпыт внедрения Бизнес и молодежь. Заработок
Бизнес и молодежь. Заработок Определение понятий предприятие, организация, фирма
Определение понятий предприятие, организация, фирма Презентация на тему Кодирование графической информации
Презентация на тему Кодирование графической информации Путешествие на планету положительных и отрицательных чисел
Путешествие на планету положительных и отрицательных чисел Имущественные риски
Имущественные риски Налоговые системы Российской Федерации и Федеративной Республики Германии. Сравнительный анализ.
Налоговые системы Российской Федерации и Федеративной Республики Германии. Сравнительный анализ. Стон блокадного Ленинграда
Стон блокадного Ленинграда Нашествие белых медведей на человеческое жилье
Нашествие белых медведей на человеческое жилье Любимое блюдо моей семьи
Любимое блюдо моей семьи Упражнения на развитие ловкости, гибкости. Акробатические элементы
Упражнения на развитие ловкости, гибкости. Акробатические элементы Мастер – класс по русскому языку (технология Ю.А.Поташкиной)
Мастер – класс по русскому языку (технология Ю.А.Поташкиной) Презентация1
Презентация1 Заонежские узоры
Заонежские узоры Aragon. Электрооборудование
Aragon. Электрооборудование Разработка устройства виртуальной реальности для обучения сотрудников промышленных предприятий
Разработка устройства виртуальной реальности для обучения сотрудников промышленных предприятий ковер+сон
ковер+сон Анестезияв специальных разделах хирургии:эндовидеохирургические вмешательства
Анестезияв специальных разделах хирургии:эндовидеохирургические вмешательства В мире новых профессий
В мире новых профессий CAD/CAM/CAPP ADEM
CAD/CAM/CAPP ADEM Холодильная техника BEKO Россия, Киржач2006
Холодильная техника BEKO Россия, Киржач2006 «Портфолио» учителя математики Матвеевой Светланы Викторовны
«Портфолио» учителя математики Матвеевой Светланы Викторовны Бурение скважин
Бурение скважин Презентация на тему Шляпочные
Презентация на тему Шляпочные