- Лекция 3

Содержание
- 2. Журнальный клуб 18 марта: Jimenez-Sanchez G, Childs B, Valle D. Human disease genes. Nature. 2001 Feb
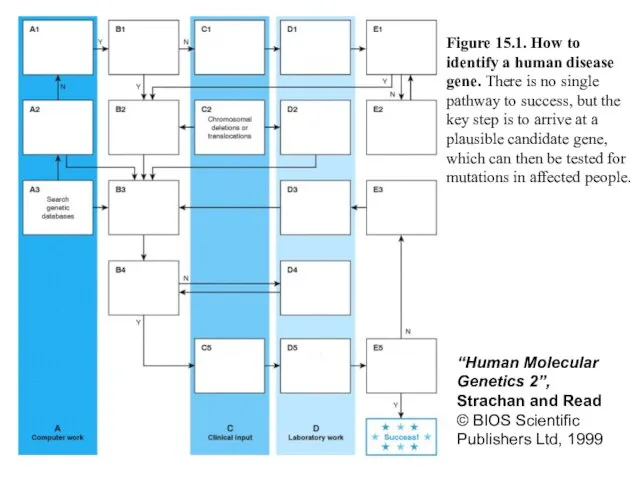
- 3. Figure 15.1. How to identify a human disease gene. There is no single pathway to success,
- 4. Стратегии поиска генов заболеваний Functional/Candidate Gene Cloning: “Either a known protein that is responsible for an
- 5. Сложные заболевания: примеры Аутизм Астма Диабет Ожирение Алкоголизм Гипертония Шизофрения
- 6. Особенности сложных заболеваний Неполная пенетрантность (Не всякая генетическая предрасположенность проявляется как болезнь) Различный возраст развития заболевания
- 8. Evolution revisited: упрощенный взгляд на болезнетворные аллели Моногенные заболевания Сильный эффект аллелей Мутационно-отборное равновесие Сложные заболевания
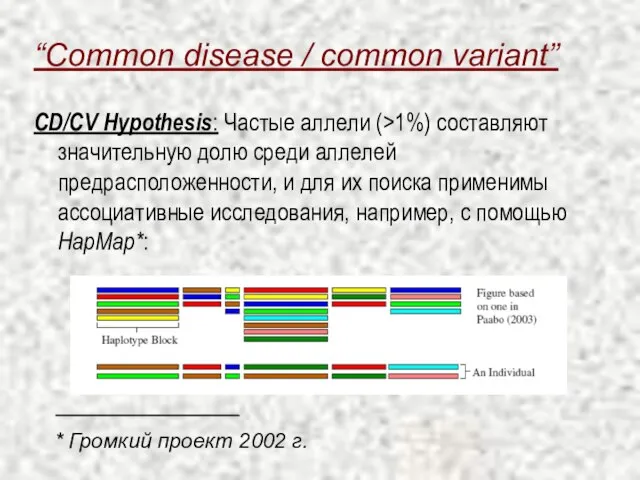
- 9. “Common disease / common variant” CD/CV Hypothesis: Частые аллели (>1%) составляют значительную долю среди аллелей предрасположенности,
- 10. “Common disease / rare variant” CD/RV Hypothesis: Редкие аллели ( * Громкий проект 2008 г.
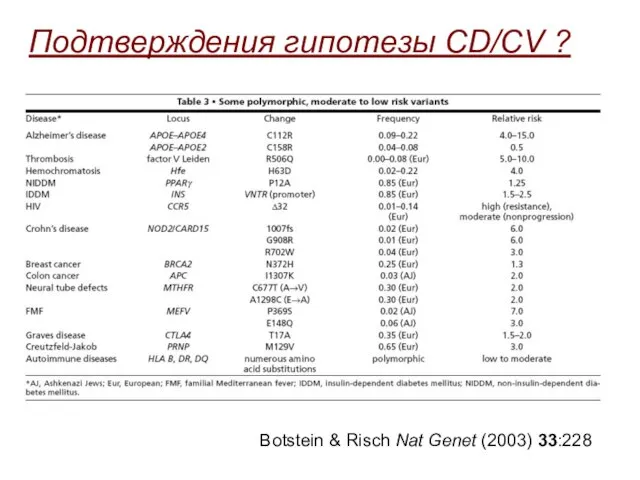
- 11. Botstein & Risch Nat Genet (2003) 33:228 Подтверждения гипотезы CD/CV ?
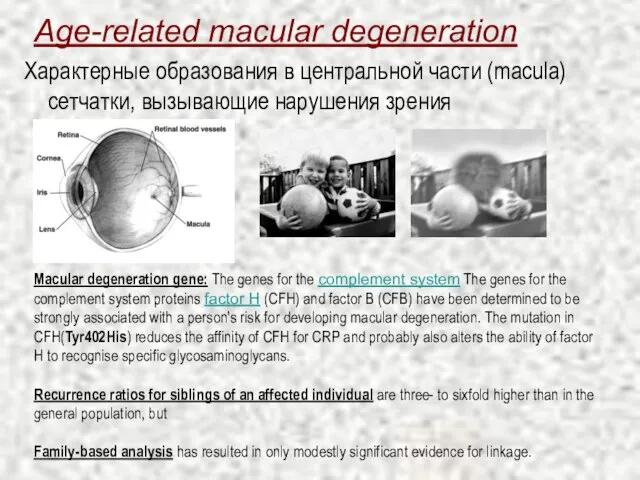
- 12. Age-related macular degeneration Характерные образования в центральной части (macula) сетчатки, вызывающие нарушения зрения Macular degeneration gene:
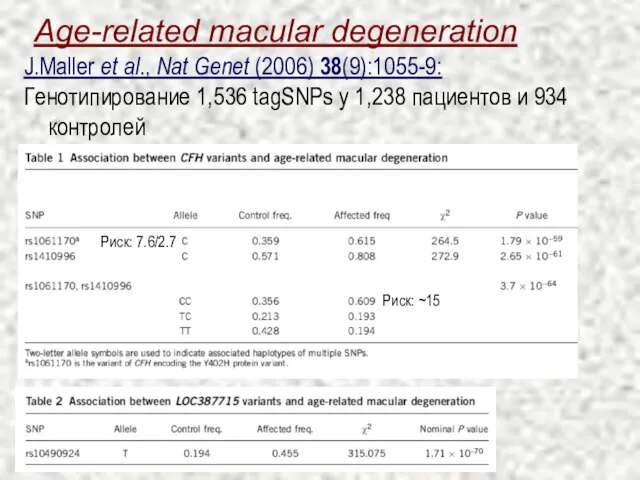
- 13. Age-related macular degeneration J.Maller et al., Nat Genet (2006) 38(9):1055-9: Генотипирование 1,536 tagSNPs у 1,238 пациентов
- 14. Age-related macular degeneration J.Maller et al., Nat Genet (2006) 38(9):1055-9: Полиморфизм в локусе, содержащем гены системы
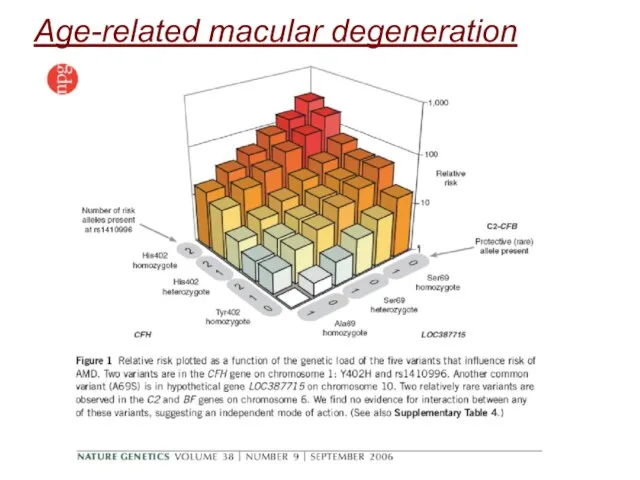
- 15. Age-related macular degeneration
- 16. Age-related macular degeneration (i) there can exist common alleles of substantial effect on common disease (in
- 17. Влияние редких аллелей на уровень холестерина высоко й плотности Низкой уровень холестерина высокой плотности (HDL-C, «хорошего»
- 18. Влияние редких аллелей на уровень холестерина высоко й плотности J.Cohen et al., Science (2004) 305:869-72 The
- 20. Скачать презентацию

Слайд 3Figure 15.1. How to identify a human disease gene. There is no
Figure 15.1. How to identify a human disease gene. There is no
Слайд 4Стратегии поиска генов заболеваний
Functional/Candidate Gene Cloning: “Either a known protein that is
Стратегии поиска генов заболеваний
Functional/Candidate Gene Cloning: “Either a known protein that is

Слайд 5Сложные заболевания: примеры
Аутизм
Астма
Диабет
Ожирение
Алкоголизм
Гипертония
Шизофрения
Сложные заболевания: примеры
Аутизм
Астма
Диабет
Ожирение
Алкоголизм
Гипертония
Шизофрения

Слайд 6Особенности сложных заболеваний
Неполная пенетрантность (Не всякая генетическая предрасположенность проявляется как болезнь)
Различный возраст
Особенности сложных заболеваний
Неполная пенетрантность (Не всякая генетическая предрасположенность проявляется как болезнь)
Различный возраст

Слайд 8Evolution revisited: упрощенный взгляд на болезнетворные аллели
Моногенные заболевания
Сильный эффект аллелей
Мутационно-отборное равновесие
Сложные заболевания
Слабый
Evolution revisited: упрощенный взгляд на болезнетворные аллели
Моногенные заболевания
Сильный эффект аллелей
Мутационно-отборное равновесие
Сложные заболевания
Слабый

Слайд 9“Common disease / common variant”
CD/CV Hypothesis: Частые аллели (>1%) составляют значительную долю
“Common disease / common variant”
CD/CV Hypothesis: Частые аллели (>1%) составляют значительную долю
Слайд 10“Common disease / rare variant”
CD/RV Hypothesis: Редкие аллели (<1%) составляют основную долю
“Common disease / rare variant”
CD/RV Hypothesis: Редкие аллели (<1%) составляют основную долю

Слайд 11Botstein & Risch Nat Genet (2003) 33:228
Подтверждения гипотезы CD/CV ?
Botstein & Risch Nat Genet (2003) 33:228
Подтверждения гипотезы CD/CV ?
Слайд 12Age-related macular degeneration
Характерные образования в центральной части (macula) сетчатки, вызывающие нарушения зрения
Macular
Age-related macular degeneration
Характерные образования в центральной части (macula) сетчатки, вызывающие нарушения зрения
Macular
Слайд 13Age-related macular degeneration
J.Maller et al., Nat Genet (2006) 38(9):1055-9:
Генотипирование 1,536 tagSNPs
Age-related macular degeneration
J.Maller et al., Nat Genet (2006) 38(9):1055-9:
Генотипирование 1,536 tagSNPs
Слайд 14Age-related macular degeneration
J.Maller et al., Nat Genet (2006) 38(9):1055-9:
Полиморфизм в локусе,
Age-related macular degeneration
J.Maller et al., Nat Genet (2006) 38(9):1055-9:
Полиморфизм в локусе,

Слайд 15Age-related macular degeneration
Age-related macular degeneration
Слайд 16Age-related macular degeneration
(i) there can exist common alleles of substantial effect on
Age-related macular degeneration
(i) there can exist common alleles of substantial effect on

Слайд 17Влияние редких аллелей на уровень холестерина высоко й плотности
Низкой уровень холестерина высокой плотности
Влияние редких аллелей на уровень холестерина высоко й плотности
Низкой уровень холестерина высокой плотности

Слайд 18Влияние редких аллелей на уровень холестерина высоко й плотности
J.Cohen et al., Science (2004)
Влияние редких аллелей на уровень холестерина высоко й плотности
J.Cohen et al., Science (2004)

 Изобразительное искусство. Викторина
Изобразительное искусство. Викторина Lec 7 Marketing information and research
Lec 7 Marketing information and research Презентация на тему Творчество Чайковского
Презентация на тему Творчество Чайковского  Выдающийся соотечественник – Михаил Васильевич Ломоносов
Выдающийся соотечественник – Михаил Васильевич Ломоносов Підліткова праця. Правовий аспект
Підліткова праця. Правовий аспект Об участии в долевом строительстве многоквартирных домов и о внесении изменений в некоторые законодательные акты РФ
Об участии в долевом строительстве многоквартирных домов и о внесении изменений в некоторые законодательные акты РФ НОВЫЕ ПОДХОДЫ К ПРИМЕНЕНИЮ ЦИТОПРОТЕКТОРОВ В КОМПЛЕКСНОЙ ТЕРАПИИ ХСН С СОПУТСТВУЮЩИМ МЕТАБОЛИЧЕСКИМ СИНДРОМОМСтаценко М.Е.Волг
НОВЫЕ ПОДХОДЫ К ПРИМЕНЕНИЮ ЦИТОПРОТЕКТОРОВ В КОМПЛЕКСНОЙ ТЕРАПИИ ХСН С СОПУТСТВУЮЩИМ МЕТАБОЛИЧЕСКИМ СИНДРОМОМСтаценко М.Е.Волг Особенности труда жителей Кубани
Особенности труда жителей Кубани Моё портфолио
Моё портфолио ДИСЦИПЛИНА БАНКОВСКИЕ РИСКИ
ДИСЦИПЛИНА БАНКОВСКИЕ РИСКИ Социологическое исследование
Социологическое исследование Город Умань Черкасской области, Украина
Город Умань Черкасской области, Украина Музею имени Героя Советского Союза Е.И. Зеленко- 40 лет!
Музею имени Героя Советского Союза Е.И. Зеленко- 40 лет! Районы орошаемого земледелия в Азии
Районы орошаемого земледелия в Азии Мотивационная программа Dentsply для розницы на II полугодие 2017 года
Мотивационная программа Dentsply для розницы на II полугодие 2017 года Бизнес-план. Кафе-чебуречная Советских Времен
Бизнес-план. Кафе-чебуречная Советских Времен Презентация на тему Реформация в Европе (7 класс)
Презентация на тему Реформация в Европе (7 класс)  Жизнь Сергея Есенина
Жизнь Сергея Есенина Комитет по госзаказу Санкт-Петербурга. Смарт-контракты. Возможность применения технологии в АИС ГЗ
Комитет по госзаказу Санкт-Петербурга. Смарт-контракты. Возможность применения технологии в АИС ГЗ Российская коррупция – проявление рыночной рациональности или разрушения социума?Необходимость общественной дискуссии и роль
Российская коррупция – проявление рыночной рациональности или разрушения социума?Необходимость общественной дискуссии и роль  Муниципальное образовательное учреждение «Пежемскаяобщеобразовательная средняя школа №14»
Муниципальное образовательное учреждение «Пежемскаяобщеобразовательная средняя школа №14» Подвеска из серебра с использованием символики FIFA 2018
Подвеска из серебра с использованием символики FIFA 2018 Содержание и методика подготовки исследовательских работ обучающихся по краеведению
Содержание и методика подготовки исследовательских работ обучающихся по краеведению Основы менеджмента
Основы менеджмента Kula Pro Presentations Bundle
Kula Pro Presentations Bundle Строение растений
Строение растений Смешанные числа
Смешанные числа Здравствуйте! Меня зовут Ершова Людмила Алексеевна.
Здравствуйте! Меня зовут Ершова Людмила Алексеевна.