- Механизмы иммунологической толерантности и иммунопатологии

Содержание
- 2. Благодарю за внимание!
- 3. Иммунологическая толерантность (терпимость) 1- активное состояние иммунной системы, которое характеризуется неотвечаемостью на антигены (требуется предварительный контакт
- 4. Иммунологическая толерантность формируется естественными и искусственными путями. Естественные пути: Физиологические: -терпимость к своим молекулам поддерживается в
- 5. Пример физиологической толерантности. Толерантность матери к плоду
- 7. Клеточный микрохимеризм мать-ребенок
- 8. История идей в изучении толерантности
- 9. Американский генетик R.OWEN, впервые описавший в 1945 году эритроцитарный мозаицизм у дизиготных телят-двоен (химеризм)
- 10. 1960
- 11. Активно приобретенная толерантность Billingham RE, Brent L, Medawar P. Nature,1953;172:603-6 Взята самка СВА на 15—16-й день
- 12. Активно приобретенная толерантность к чужим клеткам при неонатальном введении живых клеток (рис из А.А.Ярилин,2010)
- 13. Виды толерантности
- 14. Виды иммунологической толерантности Антигеспецифическая Естественная Индуцированная Антиген-неспецифическая Врожденная Индуцированная
- 15. Антигеспецифическая Естественная Толерантность к молекулам своего организма (аутотолерантность) Формирование толерантности к своим молекулам в процессе эмбриогенеза
- 16. Антиген-специфическая индуцированная толерантность Введение антигена в эмбрион или в первые сутки после рождения (эксперименты, врожденные и
- 17. Антигеннеспецифическая иммунологическая врожденная толерантность Обусловлена неспособностью к иммунному ответу при: Грубых генетических дефектах в иммунной системе
- 18. Сбой толерантности при генетических нарушениях
- 19. Мутация в гамма-цепи гена рецептора ИЛ-2 приводит к тяжелому комбинированному иммунодефициту
- 21. Антигеннеспецифическая индуцированная толерантность Супрессивные воздействия на иммунную систему Инфекционные Радиационные Токсические Лекарственные и др. Удаление лимфоидных
- 22. Индукции специфической аутотолерантности предшествует контакт с антигеном Запоминание «своих» молекул в эмбриональном периоде Центральная толерантность –
- 24. Клеточный ответ Толерантность Гуморальный ответ
- 25. Механизмы аутотолерантности Т-клеток Центральный Удаление аутореактивных клонов (гены AIRE – аутоиммунные регуляторы) Периферический «Иммунологическая привилегия» органов
- 26. Центральная аутотолерантность
- 27. Двойное распознавание антигена Антиген- презенти- рующая клетка (МНС) Антиген- специфичный рецептор Т клетки (ТСR)
- 28. Негативная селекция аутореактивных В и Т клеток
- 29. Позитивная селекция для тимоцитов, несущих рецепторы, способные к связыванию собственных молекул гистосовместимости (молекул MHC), которая обозначается
- 30. Механизмы селекции Т-лимфоцитов в тимусе
- 31. Роль гена Aire(АутоИммунного РЕгулятора) в продукции аутоантигенов для негативной селекции, создающей центральную толерантность
- 32. Селекция В-лимфоцитов в костном мозге
- 33. Редактирование аутореактивного рецептора В-клеток (ВСR IgM)
- 34. Гомеостатическая толерантность
- 36. Естественная регуляторная Т-клетка созревает в тимусе
- 37. Центральное и периферическое происхождение регуляторных Т-клеток
- 38. Регуляторные Т-клетки подавляют иммунный ответ на разных стадиях
- 39. Сокращение числа эффекторных Т-клеток на 90-95% после удаления патогена происходит по следующим причинам: Большое число оставшихся
- 40. Периферическая аутотолерантность
- 41. К периферической толерантности Незаметность антигенов для иммунной системы. Она проявляется по отношению к некоторым антигенам, расположенным
- 42. Разделение аутореактивных циркулирующих Т-лимфоцитов и аутоантигенов Для предотвращения доступа антигенпредставляющих клеток к достаточно большим количествам своих
- 44. Для активации Т-клеток нужен двойной сигнал
- 45. Поддержание толерантности к аутоантигенам на периферии
- 46. Толерантность в привилегированных зонах- яичке и глазном яблоке Для предотвращения воспаления и последующей аутоиммунной реакции в
- 47. В-клеточная толерантность В-клеточная толерантность выражена в меньшей степени, чем толерантность Т-клеток в связи с отсутствием эквивалента
- 48. Оральная толерантность
- 49. Иммунная система кишечника
- 50. Оральная толерантность при лечении аллергии
- 51. ИММУННЫЕ МЕХАНИЗМЫ РАЗВИТИЯ ПАТОЛОГИЧЕСКИХ ПРОЦЕССОВ Клеточный ответ Гуморальный ответ
- 52. Анафилактическая гиперчувствитель-ность немедленного типа (ГНТ) В 1963 г. Кумбс и Джелл предложили классификацию гиперчувствительности, согласно которой
- 53. АБЗИМ - АНТИТЕЛО С ФЕРМЕНТАТИВНОЙ АКТИВНОСТЬЮ
- 54. МЕХАНИЗМЫ РЕАЛИЗАЦИИ ЦИТОТОКСИЧЕСКОГО ПОТЕНЦИАЛА ДНК-АБЗИМОВ
- 55. ИММУНОПАТОЛОГИЯ ПРИ САХАРНОМ ДИАБЕТЕ
- 56. К механизму развития рассеянного склероза Figure 4. The Multiple Sclerosis Plaque. Within a postcapillary venule, various
- 57. Эктопическая лимфоидная ткань У части пациентов с аутоиммунными болезнями в мишеневом органе эктопически развивается организованная лимфоидная
- 58. Инфекция и аутоиммунитет Аутоиммунные болезни связаны с вирусными инфекциями, которые часто предшествуют возникновению воспаления в мишеневом
- 59. Молекулярная мимикрия между белками патогенных микробов и хозяина.
- 60. К механизмам развития иммунопатологических процессов (1/3) К настоящему времени открыты новые механизмы индукции и развития иммунопатологических
- 61. К механизмам развития иммунопатологических процессов (2/3) 11. Антигенная мимикрия между молекулами патогенов и хозяина 12. Индукция
- 62. К механизмам развития иммунопатологических процессов (3/3) 21. Лимфопролиферативные заболевания 22. Злокачественные новообразования 23. Антиммунные воздействия патогенов
- 63. Формирование аутореактивных Т-клеток памяти при вирусной инфекции. Срыв толерантности к аутоантигенам при ослаблении Т-регуляторных клеток
- 64. Аутоиммунные болезни как следствие вакцинации
- 65. Т-клетки памяти формируются в начале ответа и после его завершения
- 67. Миграция Т-клеток памяти: центральной и эффекторов
- 69. Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней Несколько работ свидетельствует о роли Т-клеток памяти
- 70. Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней (продолжение) Изолированные T-клетки памяти с фенотипом CD4+CCR4+CD44high
- 71. Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней (продолжение) Т- клетки памяти имеют повышенную функциональную
- 72. Имеются экспериментальные доказательстива, поддерживающие гипотезу о чрезпеченочной циркуляции кишечных лимфоцитов памяти, возникших при воспалении в слизистой
- 73. Фиброз вокруг желчного протока при склерозирующем холангите
- 74. Интегрины α4β7 Т лимфоцитов соединяются с адгезивными адрессинами слизистой MAdCAM-1
- 75. Формирование провоспалительных Т клеток памяти с молекулами хоминга
- 76. МЕХАНИЗМ РАЗРУШЕНИЯ Т-КЛЕТОК ПАМЯТИ В БЛЯШКЕ ПРИ ПСОРИАЗЕ АЛЕФАЦЕПТОМ New England Journal of Medicine Volume 345:248-255
- 77. Увидел – расскажи другу!
- 78. Peripheral Mechanisms of the Induction of Tolerance. T cells that are physically separated from their specific
- 81. Медицинский Уотергейт-1974 Доктор Вильям Саммерлин был руководителем клиники кожных болезней в Стэнфордском университете в Калифорнии. Результаты
- 83. Скачать презентацию

Слайд 3
Иммунологическая толерантность (терпимость)
1- активное состояние иммунной системы, которое характеризуется неотвечаемостью на
Иммунологическая толерантность (терпимость)
1- активное состояние иммунной системы, которое характеризуется неотвечаемостью на

Слайд 4Иммунологическая толерантность
формируется
естественными и искусственными путями.
Естественные пути:
Физиологические:
-терпимость к своим молекулам поддерживается
в течение
Иммунологическая толерантность
формируется
естественными и искусственными путями.
Естественные пути:
Физиологические:
-терпимость к своим молекулам поддерживается
в течение

Слайд 5
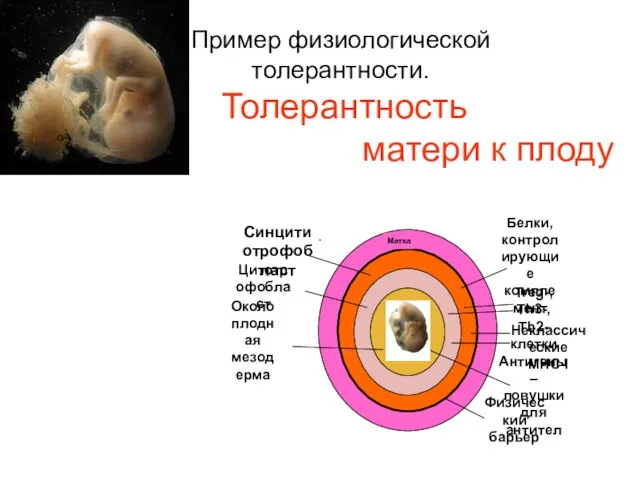
Пример физиологической
толерантности.
Толерантность матери к плоду
Пример физиологической
толерантности.
Толерантность матери к плоду

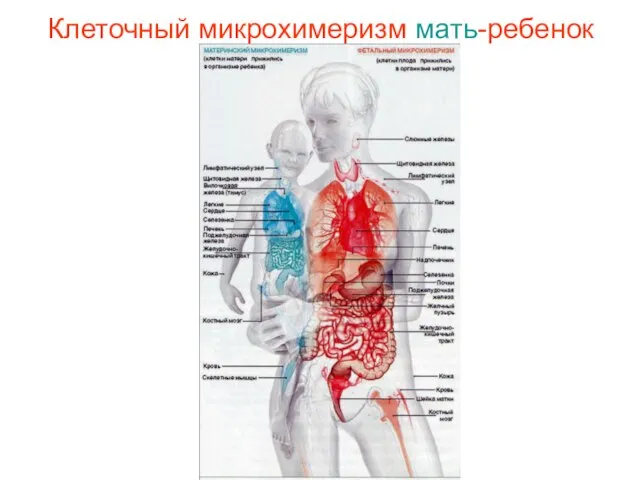
Слайд 7Клеточный микрохимеризм мать-ребенок
Клеточный микрохимеризм мать-ребенок
Слайд 8История идей
в изучении толерантности
История идей
в изучении толерантности

Слайд 9Американский генетик R.OWEN, впервые описавший в 1945 году эритроцитарный мозаицизм
у дизиготных
Американский генетик R.OWEN, впервые описавший в 1945 году эритроцитарный мозаицизм
у дизиготных

Слайд 101960
1960

Слайд 11Активно приобретенная толерантность
Billingham RE, Brent L, Medawar P. Nature,1953;172:603-6
Взята самка СВА на
Активно приобретенная толерантность
Billingham RE, Brent L, Medawar P. Nature,1953;172:603-6
Взята самка СВА на

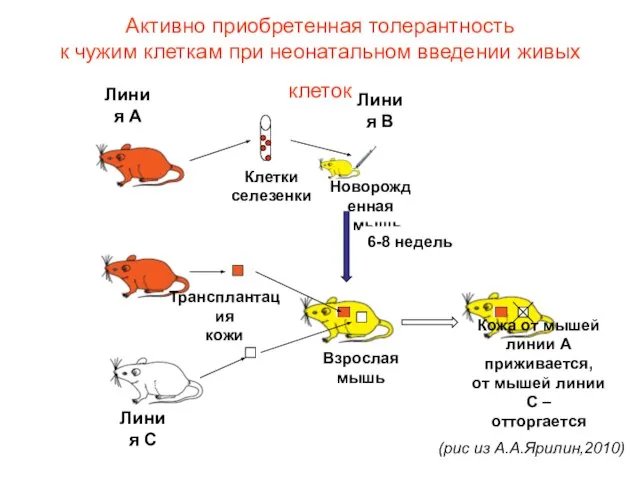
Слайд 12Активно приобретенная толерантность
к чужим клеткам при неонатальном введении живых клеток
Активно приобретенная толерантность
к чужим клеткам при неонатальном введении живых клеток
Слайд 13Виды толерантности
Виды толерантности

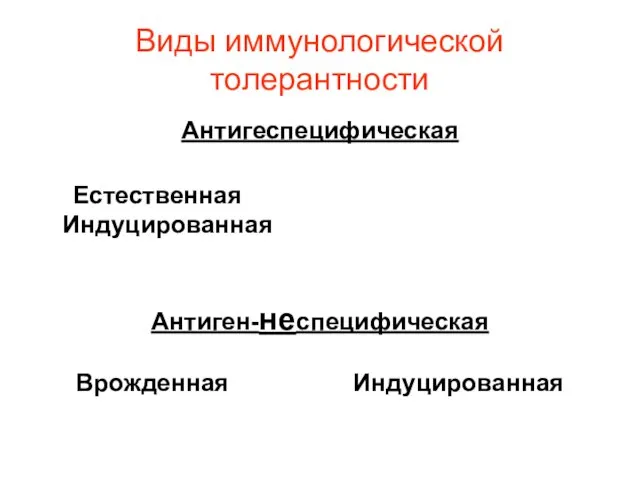
Слайд 14Виды иммунологической толерантности
Антигеспецифическая
Естественная Индуцированная
Антиген-неспецифическая
Врожденная Индуцированная
Виды иммунологической толерантности
Антигеспецифическая
Естественная Индуцированная
Антиген-неспецифическая
Врожденная Индуцированная
Слайд 15Антигеспецифическая
Естественная
Толерантность к молекулам своего организма (аутотолерантность)
Формирование толерантности к своим молекулам в процессе
Антигеспецифическая
Естественная
Толерантность к молекулам своего организма (аутотолерантность)
Формирование толерантности к своим молекулам в процессе

Слайд 16Антиген-специфическая
индуцированная толерантность
Введение антигена в эмбрион или в первые сутки после
Антиген-специфическая
индуцированная толерантность
Введение антигена в эмбрион или в первые сутки после

Слайд 17Антигеннеспецифическая иммунологическая врожденная толерантность
Обусловлена неспособностью к иммунному ответу при:
Грубых генетических дефектах в
Антигеннеспецифическая иммунологическая врожденная толерантность
Обусловлена неспособностью к иммунному ответу при:
Грубых генетических дефектах в

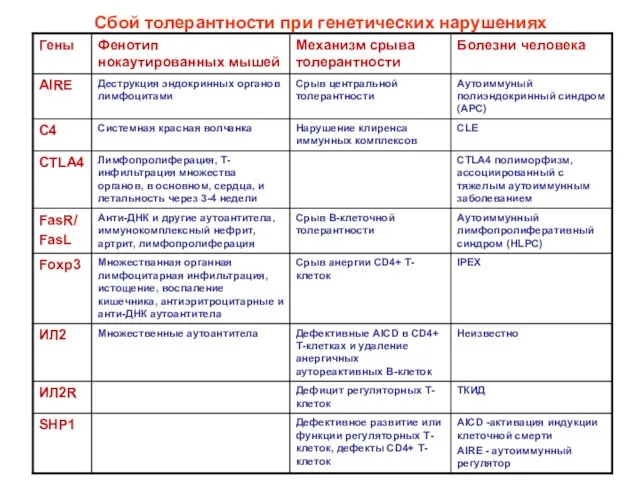
Слайд 18Сбой толерантности при генетических нарушениях
Сбой толерантности при генетических нарушениях
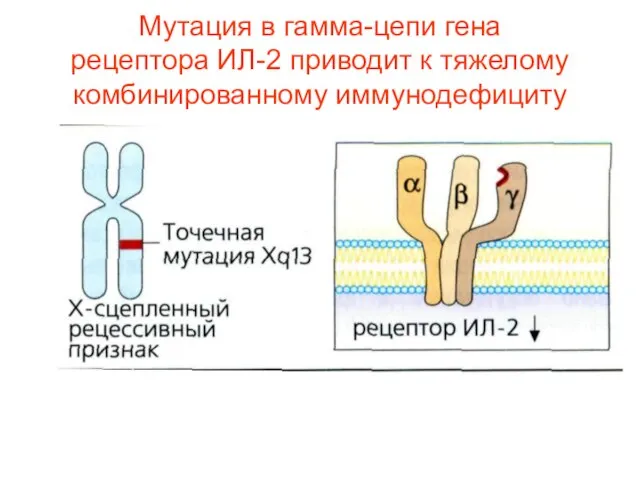
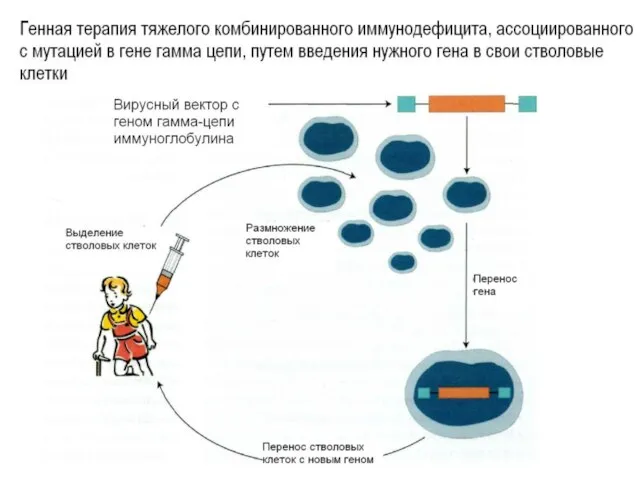
Слайд 19Мутация в гамма-цепи гена
рецептора ИЛ-2 приводит к тяжелому комбинированному иммунодефициту
Мутация в гамма-цепи гена
рецептора ИЛ-2 приводит к тяжелому комбинированному иммунодефициту
Слайд 21Антигеннеспецифическая индуцированная толерантность
Супрессивные воздействия на иммунную систему
Инфекционные
Радиационные
Токсические
Антигеннеспецифическая индуцированная толерантность
Супрессивные воздействия на иммунную систему
Инфекционные
Радиационные
Токсические

Слайд 22Индукции
специфической аутотолерантности предшествует контакт с антигеном
Запоминание «своих» молекул в эмбриональном периоде
Индукции
специфической аутотолерантности предшествует контакт с антигеном
Запоминание «своих» молекул в эмбриональном периоде

Слайд 24
Клеточный ответ Толерантность Гуморальный ответ
Клеточный ответ Толерантность Гуморальный ответ

Слайд 25Механизмы аутотолерантности Т-клеток
Центральный
Удаление аутореактивных клонов (гены AIRE – аутоиммунные регуляторы)
Периферический
«Иммунологическая привилегия» органов
Анергия
Механизмы аутотолерантности Т-клеток
Центральный
Удаление аутореактивных клонов (гены AIRE – аутоиммунные регуляторы)
Периферический
«Иммунологическая привилегия» органов
Анергия

Слайд 26Центральная аутотолерантность
Центральная аутотолерантность

Слайд 27Двойное распознавание антигена
Антиген-
презенти-
рующая
клетка
(МНС)
Антиген-
специфичный
рецептор
Двойное распознавание антигена
Антиген-
презенти-
рующая
клетка
(МНС)
Антиген-
специфичный
рецептор
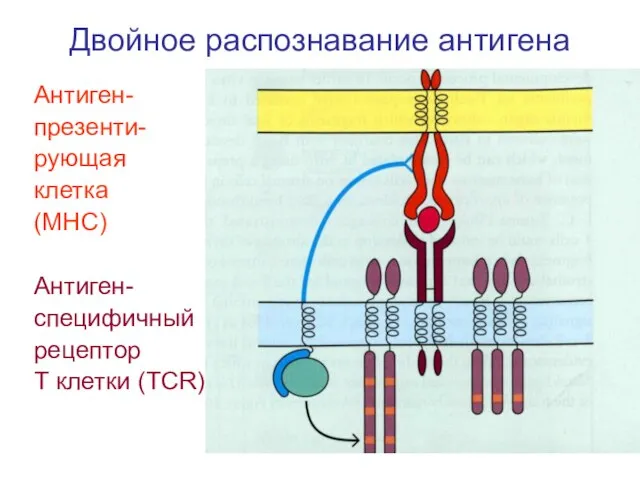
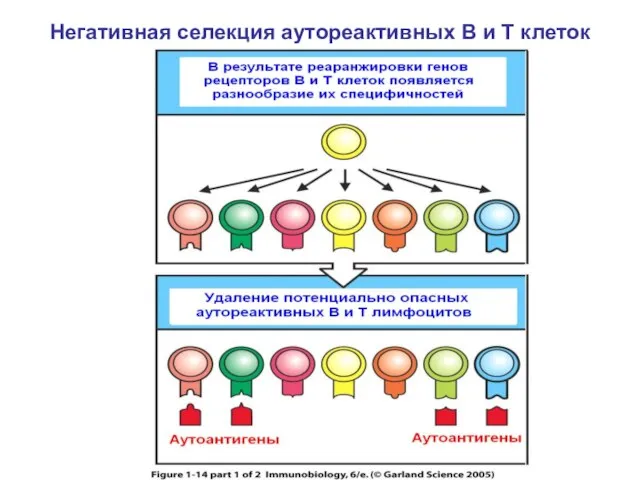
Слайд 28Негативная селекция аутореактивных В и Т клеток
Негативная селекция аутореактивных В и Т клеток
Слайд 29Позитивная селекция для тимоцитов, несущих рецепторы, способные к связыванию собственных молекул гистосовместимости
Позитивная селекция для тимоцитов, несущих рецепторы, способные к связыванию собственных молекул гистосовместимости

Слайд 30Механизмы селекции
Т-лимфоцитов в тимусе
Механизмы селекции
Т-лимфоцитов в тимусе
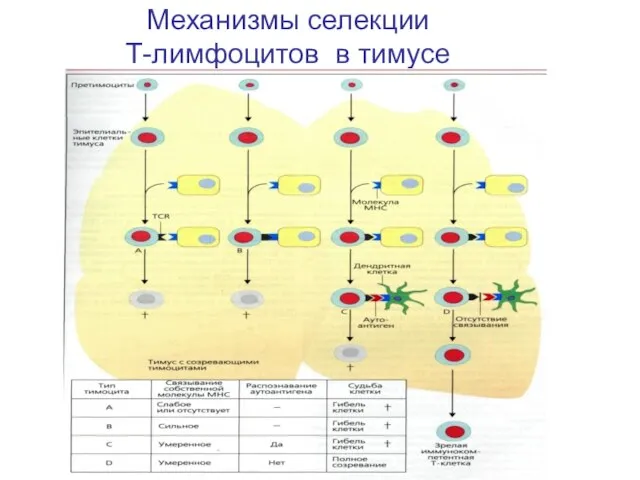
Слайд 31Роль гена Aire(АутоИммунного РЕгулятора) в продукции аутоантигенов для негативной селекции, создающей центральную
Роль гена Aire(АутоИммунного РЕгулятора) в продукции аутоантигенов для негативной селекции, создающей центральную
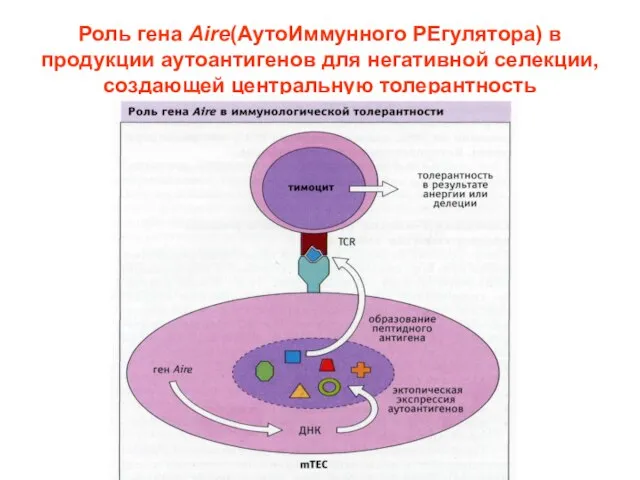
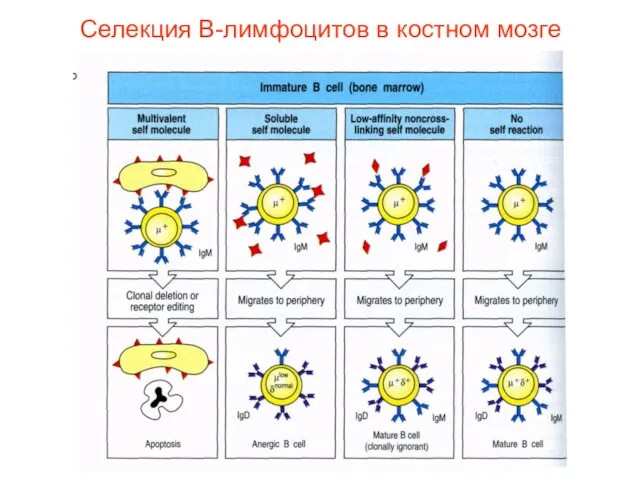
Слайд 32Селекция В-лимфоцитов в костном мозге
Селекция В-лимфоцитов в костном мозге
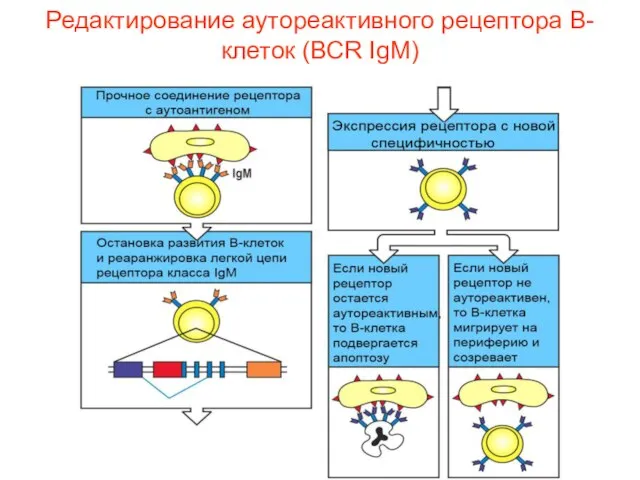
Слайд 33Редактирование аутореактивного рецептора В-клеток (ВСR IgM)
Редактирование аутореактивного рецептора В-клеток (ВСR IgM)
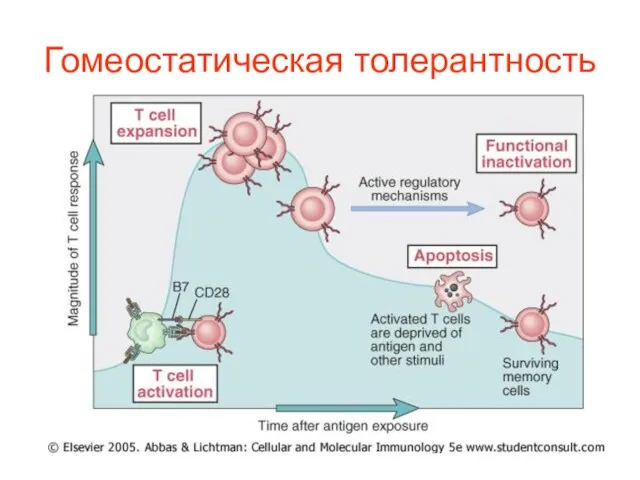
Слайд 34Гомеостатическая толерантность
Гомеостатическая толерантность
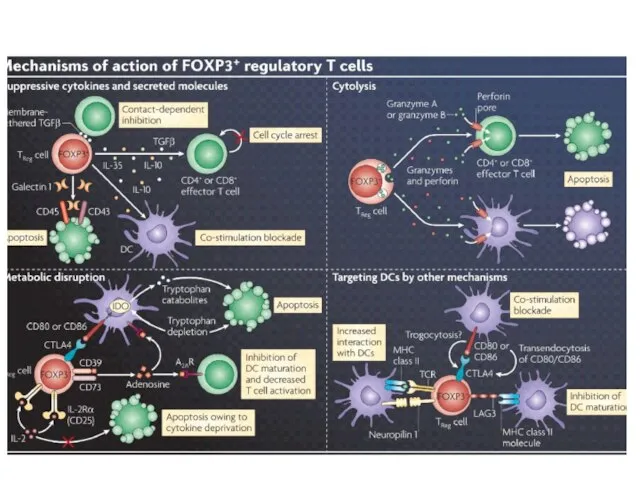
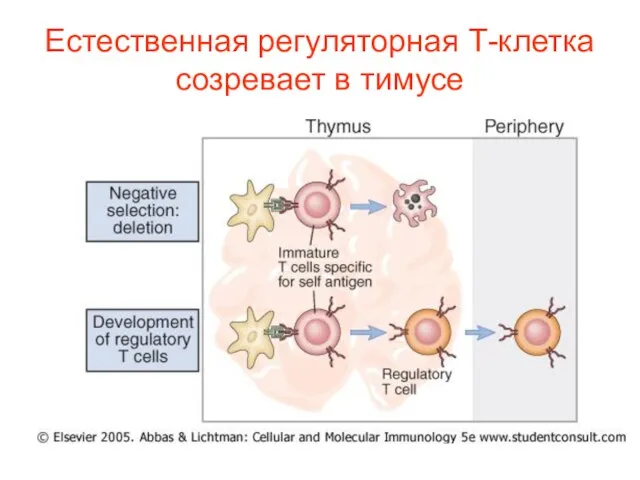
Слайд 36Естественная регуляторная Т-клетка созревает в тимусе
Естественная регуляторная Т-клетка созревает в тимусе
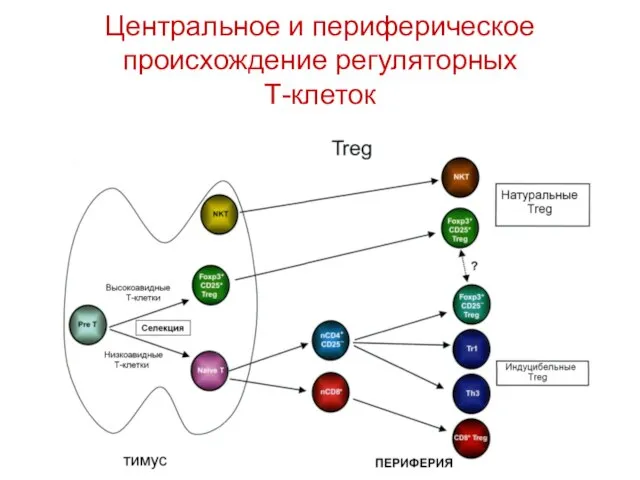
Слайд 37Центральное и периферическое происхождение регуляторных
Т-клеток
Центральное и периферическое происхождение регуляторных
Т-клеток
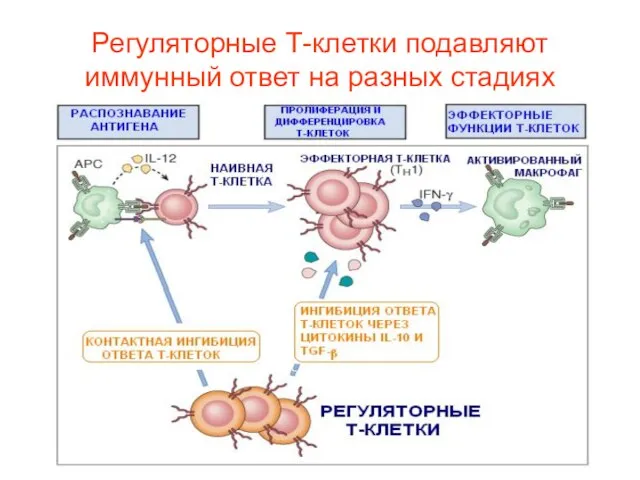
Слайд 38Регуляторные Т-клетки подавляют иммунный ответ на разных стадиях
Регуляторные Т-клетки подавляют иммунный ответ на разных стадиях
Слайд 39Сокращение числа эффекторных Т-клеток на 90-95% после удаления патогена
происходит по следующим
Сокращение числа эффекторных Т-клеток на 90-95% после удаления патогена
происходит по следующим

Слайд 40Периферическая аутотолерантность
Периферическая аутотолерантность

Слайд 41К периферической толерантности
Незаметность антигенов для иммунной системы.
Она проявляется по отношению к
К периферической толерантности
Незаметность антигенов для иммунной системы.
Она проявляется по отношению к

Слайд 42Разделение аутореактивных циркулирующих Т-лимфоцитов и аутоантигенов
Для предотвращения доступа антигенпредставляющих клеток к достаточно
Разделение аутореактивных циркулирующих Т-лимфоцитов и аутоантигенов
Для предотвращения доступа антигенпредставляющих клеток к достаточно

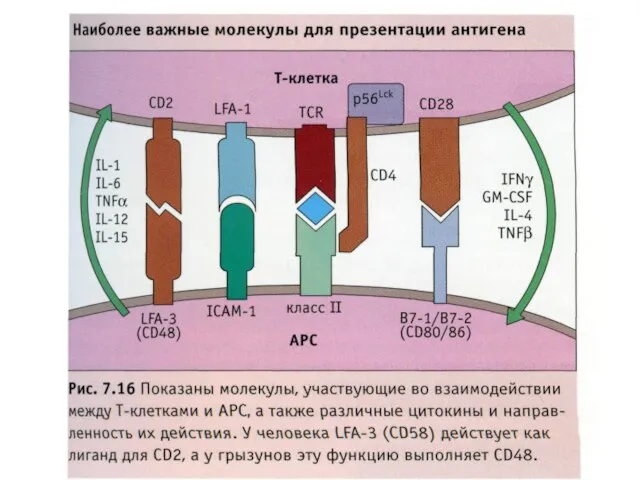
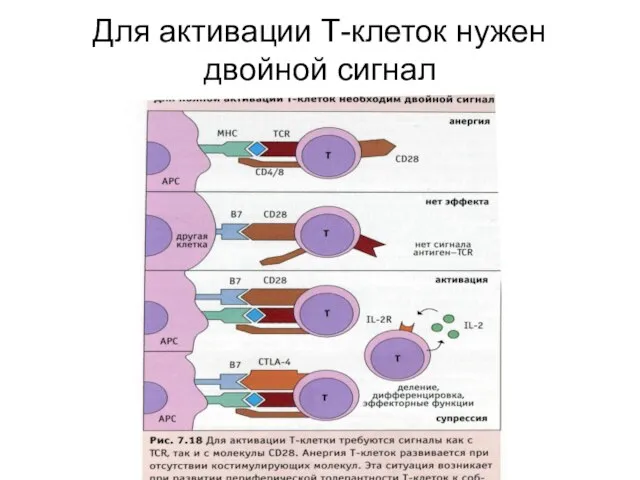
Слайд 44Для активации Т-клеток нужен двойной сигнал
Для активации Т-клеток нужен двойной сигнал
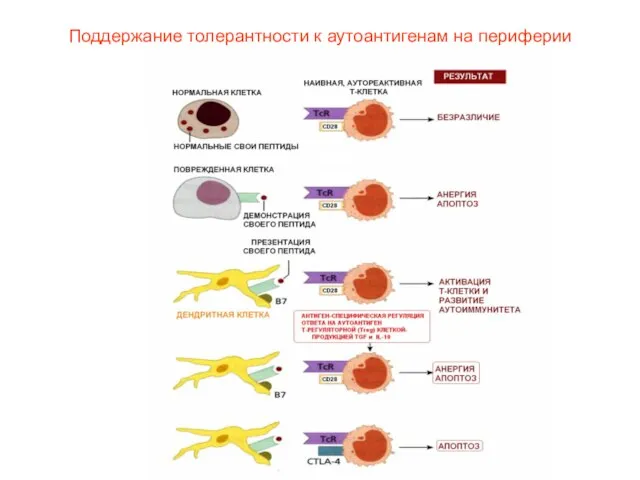
Слайд 45Поддержание толерантности к аутоантигенам на периферии
Поддержание толерантности к аутоантигенам на периферии
Слайд 46Толерантность в привилегированных зонах- яичке и глазном яблоке
Для предотвращения воспаления и последующей
Толерантность в привилегированных зонах- яичке и глазном яблоке
Для предотвращения воспаления и последующей

Слайд 47В-клеточная толерантность
В-клеточная толерантность выражена в меньшей степени, чем толерантность Т-клеток в связи
В-клеточная толерантность
В-клеточная толерантность выражена в меньшей степени, чем толерантность Т-клеток в связи

Слайд 48Оральная толерантность
Оральная толерантность

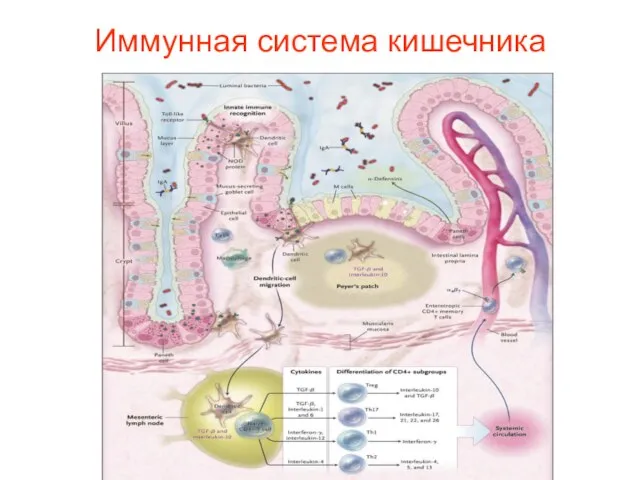
Слайд 49Иммунная система кишечника
Иммунная система кишечника
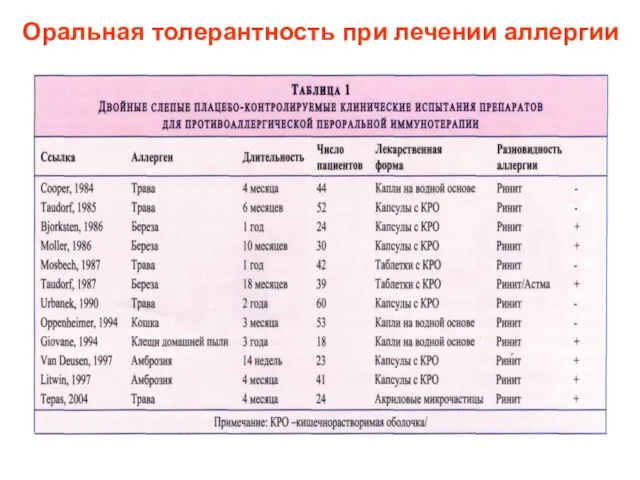
Слайд 50 Оральная толерантность при лечении аллергии
Оральная толерантность при лечении аллергии
Слайд 51
ИММУННЫЕ МЕХАНИЗМЫ
РАЗВИТИЯ ПАТОЛОГИЧЕСКИХ ПРОЦЕССОВ
Клеточный
ответ
Гуморальный
ответ
ИММУННЫЕ МЕХАНИЗМЫ
РАЗВИТИЯ ПАТОЛОГИЧЕСКИХ ПРОЦЕССОВ
Клеточный
ответ
Гуморальный
ответ

Слайд 52Анафилактическая
гиперчувствитель-ность немедленного типа (ГНТ)
В 1963 г. Кумбс и Джелл предложили
Анафилактическая
гиперчувствитель-ность немедленного типа (ГНТ)
В 1963 г. Кумбс и Джелл предложили
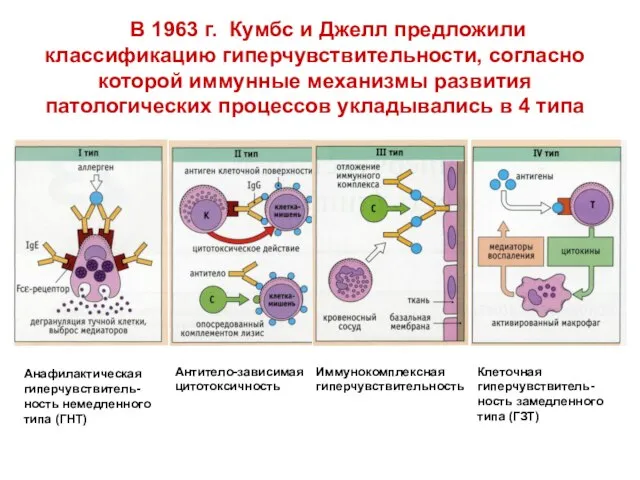
Слайд 53АБЗИМ - АНТИТЕЛО С ФЕРМЕНТАТИВНОЙ АКТИВНОСТЬЮ
АБЗИМ - АНТИТЕЛО С ФЕРМЕНТАТИВНОЙ АКТИВНОСТЬЮ
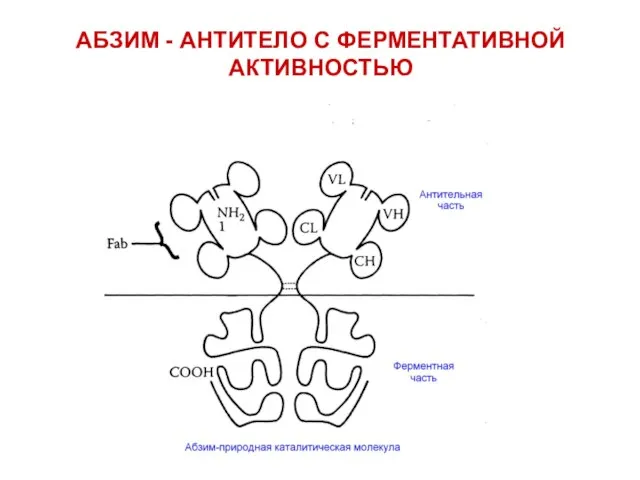
Слайд 54МЕХАНИЗМЫ РЕАЛИЗАЦИИ ЦИТОТОКСИЧЕСКОГО
ПОТЕНЦИАЛА ДНК-АБЗИМОВ
МЕХАНИЗМЫ РЕАЛИЗАЦИИ ЦИТОТОКСИЧЕСКОГО
ПОТЕНЦИАЛА ДНК-АБЗИМОВ
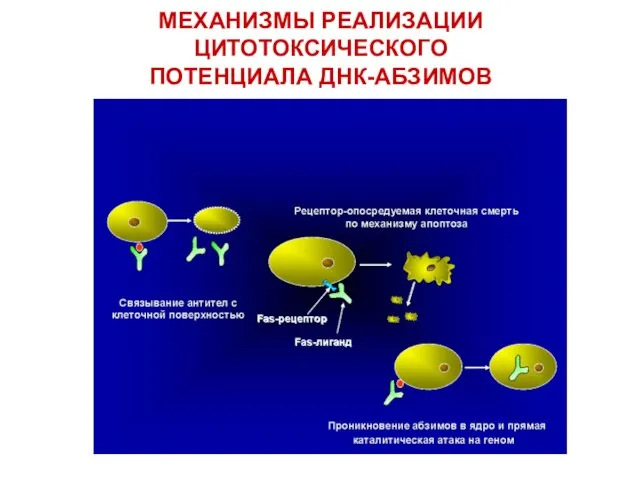
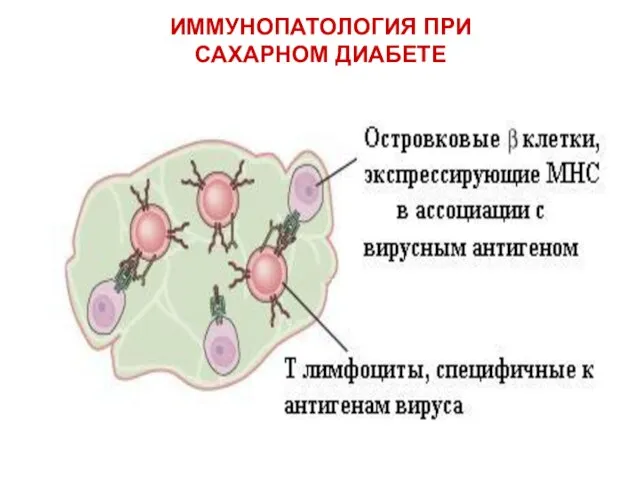
Слайд 55ИММУНОПАТОЛОГИЯ ПРИ
САХАРНОМ ДИАБЕТЕ
ИММУНОПАТОЛОГИЯ ПРИ
САХАРНОМ ДИАБЕТЕ
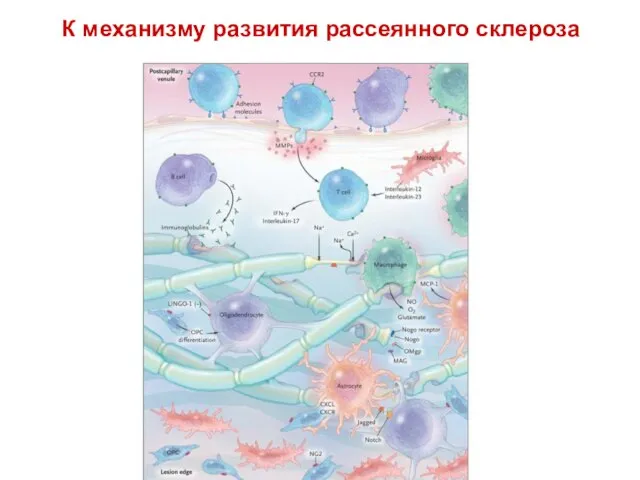
Слайд 56К механизму развития рассеянного склероза
Figure 4. The Multiple Sclerosis Plaque.
Within a
К механизму развития рассеянного склероза
Figure 4. The Multiple Sclerosis Plaque.
Within a
Слайд 57Эктопическая лимфоидная ткань
У части пациентов с аутоиммунными болезнями в мишеневом органе эктопически
Эктопическая лимфоидная ткань
У части пациентов с аутоиммунными болезнями в мишеневом органе эктопически

Слайд 58Инфекция и аутоиммунитет
Аутоиммунные болезни связаны с вирусными инфекциями, которые часто предшествуют возникновению
Инфекция и аутоиммунитет
Аутоиммунные болезни связаны с вирусными инфекциями, которые часто предшествуют возникновению

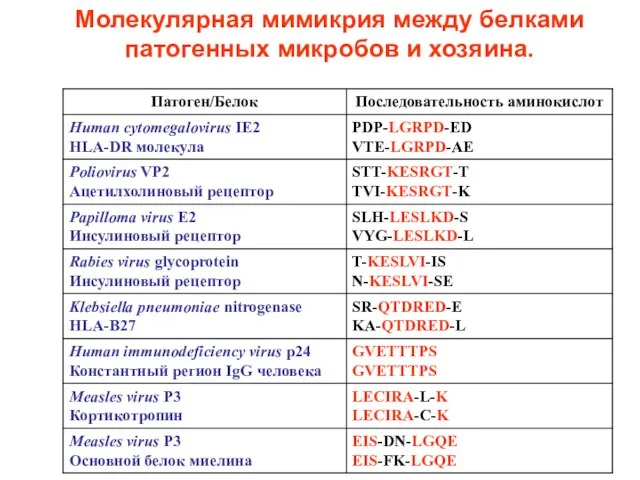
Слайд 59Молекулярная мимикрия между белками
патогенных микробов и хозяина.
Молекулярная мимикрия между белками
патогенных микробов и хозяина.
Слайд 60К механизмам развития иммунопатологических процессов (1/3)
К настоящему времени открыты новые механизмы
индукции
К механизмам развития иммунопатологических процессов (1/3)
К настоящему времени открыты новые механизмы
индукции

Слайд 61
К механизмам развития иммунопатологических процессов (2/3)
11. Антигенная мимикрия между молекулами патогенов и
К механизмам развития иммунопатологических процессов (2/3)
11. Антигенная мимикрия между молекулами патогенов и

Слайд 62
К механизмам развития иммунопатологических процессов (3/3)
21. Лимфопролиферативные заболевания
22. Злокачественные новообразования
23. Антиммунные воздействия
К механизмам развития иммунопатологических процессов (3/3)
21. Лимфопролиферативные заболевания
22. Злокачественные новообразования
23. Антиммунные воздействия

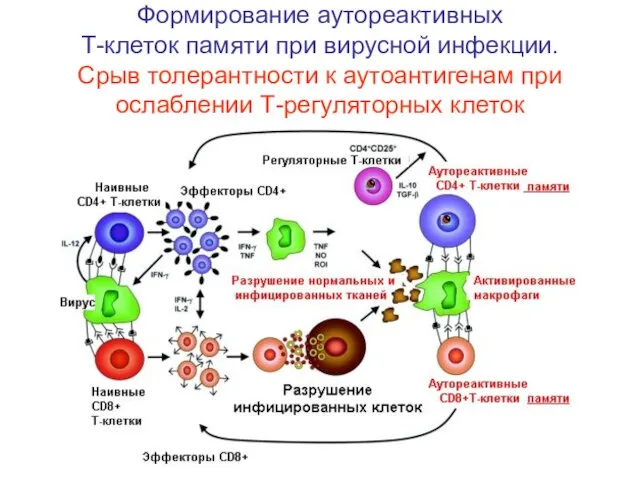
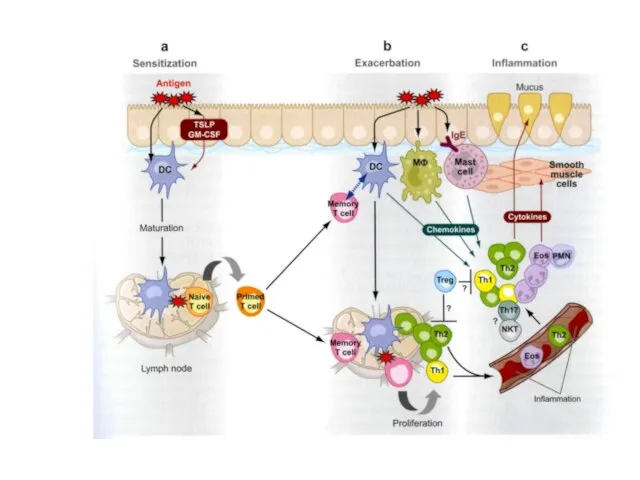
Слайд 63Формирование аутореактивных
Т-клеток памяти при вирусной инфекции.
Срыв толерантности к аутоантигенам при
Формирование аутореактивных Т-клеток памяти при вирусной инфекции. Срыв толерантности к аутоантигенам при
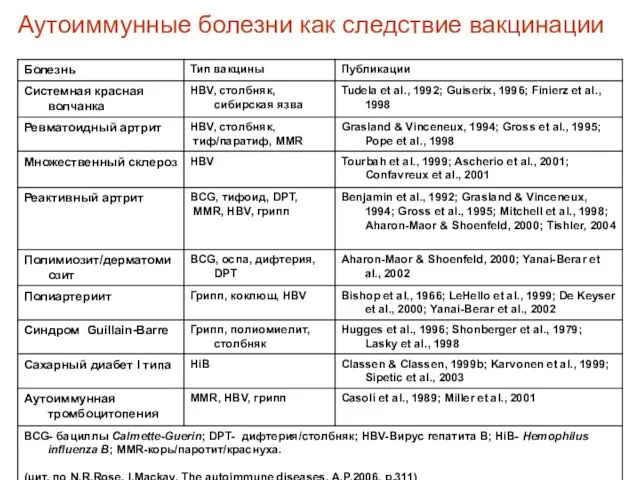
Слайд 64Аутоиммунные болезни как следствие вакцинации
Аутоиммунные болезни как следствие вакцинации
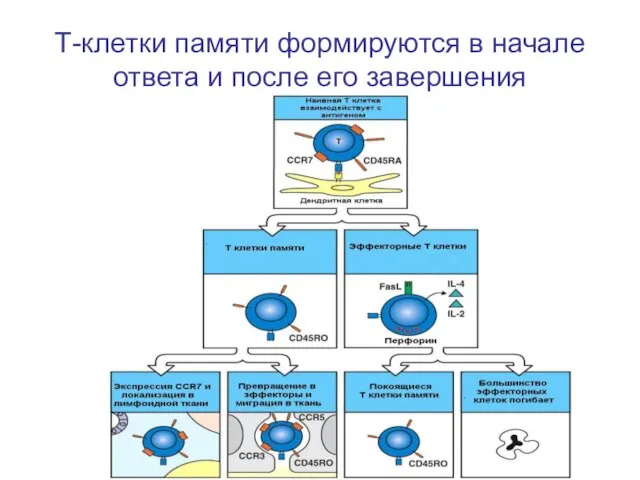
Слайд 65Т-клетки памяти формируются в начале ответа и после его завершения
Т-клетки памяти формируются в начале ответа и после его завершения
Слайд 67Миграция Т-клеток памяти:
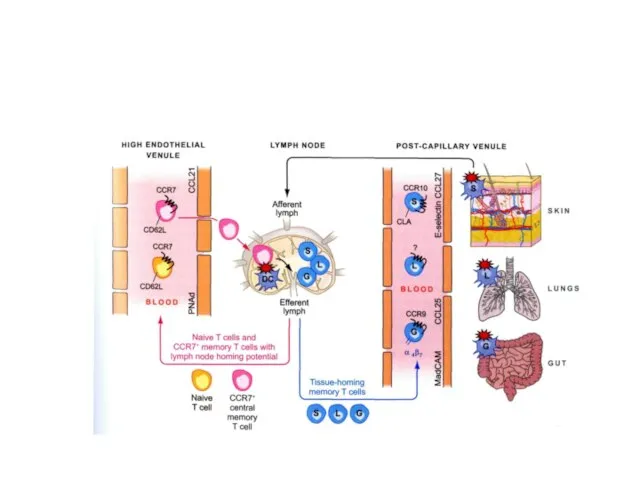
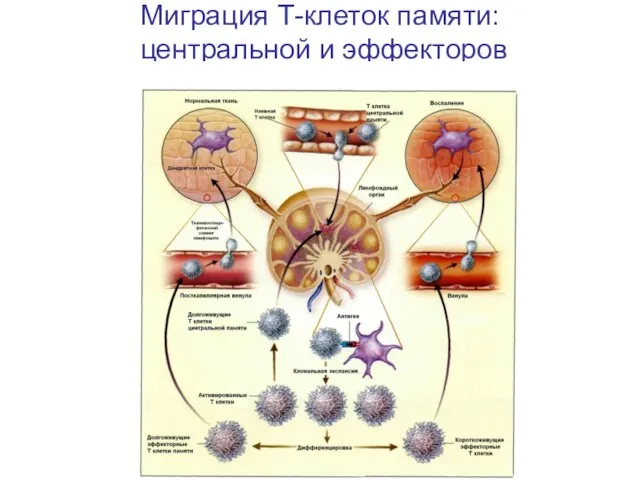
центральной и эффекторов
Миграция Т-клеток памяти:
центральной и эффекторов
Слайд 69Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней
Несколько работ свидетельствует
Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней
Несколько работ свидетельствует

Слайд 70Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней (продолжение)
Изолированные T-клетки памяти
Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней (продолжение)
Изолированные T-клетки памяти

Слайд 71Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней (продолжение)
Т- клетки памяти
Роль Т-клеток памяти в прогрессии или рецидиве аутоиммунных болезней (продолжение)
Т- клетки памяти

Слайд 72Имеются экспериментальные доказательстива, поддерживающие гипотезу о чрезпеченочной циркуляции кишечных лимфоцитов памяти, возникших
Имеются экспериментальные доказательстива, поддерживающие гипотезу о чрезпеченочной циркуляции кишечных лимфоцитов памяти, возникших

Слайд 73Фиброз вокруг желчного протока при склерозирующем холангите
Фиброз вокруг желчного протока при склерозирующем холангите

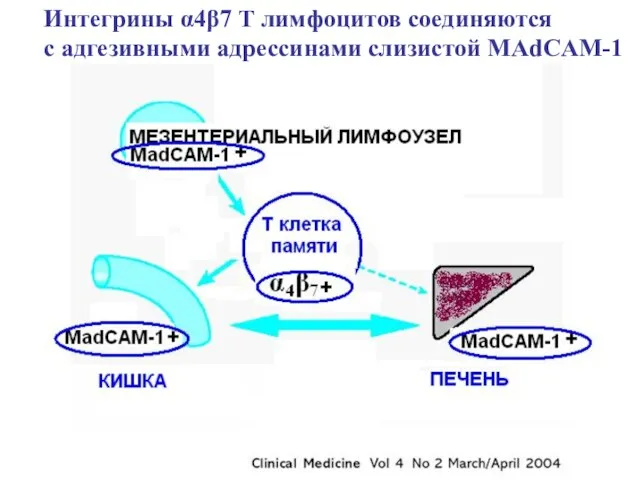
Слайд 74Интегрины α4β7 Т лимфоцитов соединяются
с адгезивными адрессинами слизистой MAdCAM-1
Интегрины α4β7 Т лимфоцитов соединяются
с адгезивными адрессинами слизистой MAdCAM-1
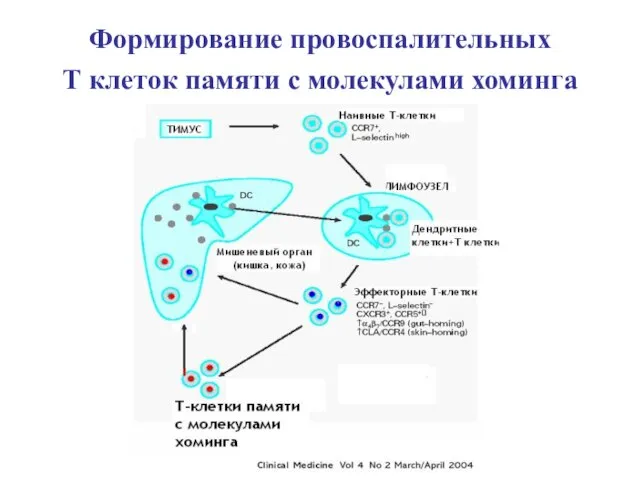
Слайд 75Формирование провоспалительных
Т клеток памяти с молекулами хоминга
Формирование провоспалительных
Т клеток памяти с молекулами хоминга
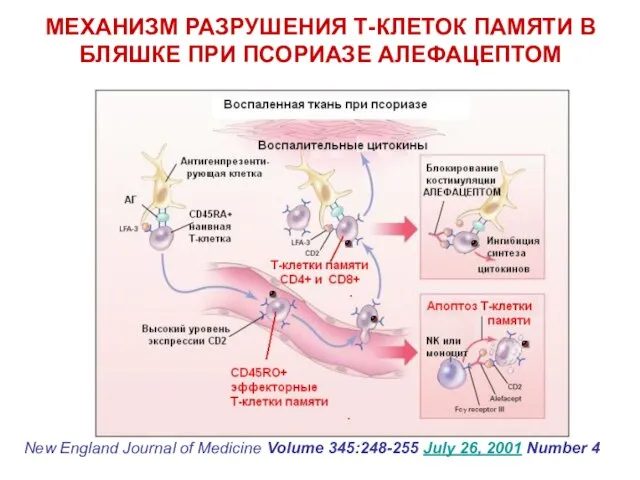
Слайд 76МЕХАНИЗМ РАЗРУШЕНИЯ Т-КЛЕТОК ПАМЯТИ В БЛЯШКЕ ПРИ ПСОРИАЗЕ АЛЕФАЦЕПТОМ
New England Journal of
МЕХАНИЗМ РАЗРУШЕНИЯ Т-КЛЕТОК ПАМЯТИ В БЛЯШКЕ ПРИ ПСОРИАЗЕ АЛЕФАЦЕПТОМ
New England Journal of
Слайд 77Увидел – расскажи другу!
Увидел – расскажи другу!

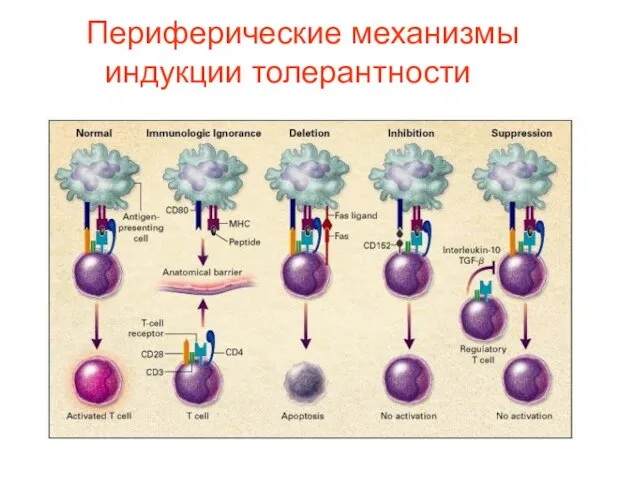
Слайд 78Peripheral Mechanisms of the Induction of Tolerance.
T cells that are physically
Peripheral Mechanisms of the Induction of Tolerance. T cells that are physically


Слайд 81Медицинский Уотергейт-1974
Доктор Вильям Саммерлин был руководителем клиники кожных болезней в Стэнфордском университете
Медицинский Уотергейт-1974
Доктор Вильям Саммерлин был руководителем клиники кожных болезней в Стэнфордском университете

 Виконання зварного з'єднання
Виконання зварного з'єднання Леди и джентльмены
Леди и джентльмены Сочинение-рассуждение на лингвистическую тему 3 часть ОГЭ (15.1)
Сочинение-рассуждение на лингвистическую тему 3 часть ОГЭ (15.1) Презентация на тему Упражнения для уроков по дифференциации парных согласных З-С
Презентация на тему Упражнения для уроков по дифференциации парных согласных З-С Презентация на тему Презентация История религий
Презентация на тему Презентация История религий  Классификация полимеров 11 класс Соловова Е.А., учитель химии высшей квалификационной категории, МОУ СОШ №15 г.Балашов, 2010
Классификация полимеров 11 класс Соловова Е.А., учитель химии высшей квалификационной категории, МОУ СОШ №15 г.Балашов, 2010  Мое книжное лето
Мое книжное лето Презентация на тему Нервная система и её роль в организме (3 класс)
Презентация на тему Нервная система и её роль в организме (3 класс) Работа с электронными таблицами и ведение электронного журнала успеваемости
Работа с электронными таблицами и ведение электронного журнала успеваемости Анализ работы кафедры общественных и гуманитарных наукза 2010 – 2011 учебный год
Анализ работы кафедры общественных и гуманитарных наукза 2010 – 2011 учебный год Урок - путешествие. В гостях у частей речи
Урок - путешествие. В гостях у частей речи ОРУ. Начало. Инструментарий. Организации. Руководства и управления
ОРУ. Начало. Инструментарий. Организации. Руководства и управления Движение по окружности
Движение по окружности Аппаратно-программный комплекс для контроля частотных характеристик обмоток силовых трансформаторов ГОУ
Аппаратно-программный комплекс для контроля частотных характеристик обмоток силовых трансформаторов ГОУ  Метод проектов
Метод проектов Opposites
Opposites Коммерческие юридические лица
Коммерческие юридические лица Мой город - моя Родина - презентация для начальной школы_
Мой город - моя Родина - презентация для начальной школы_ Урок чтения в 1 классе
Урок чтения в 1 классе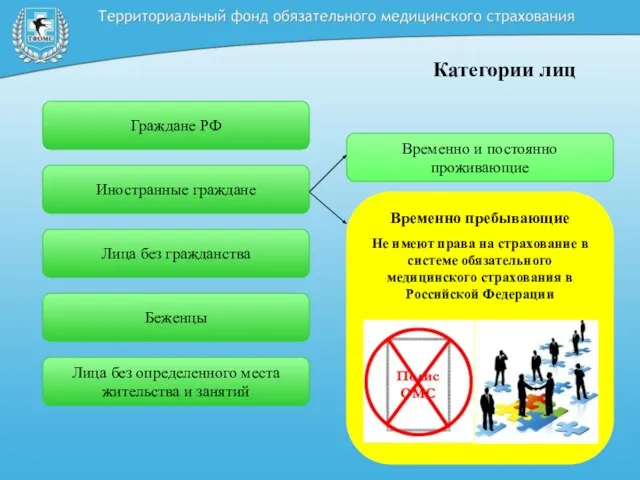 Категории лиц
Категории лиц Процесс локализации из России на Запад
Процесс локализации из России на Запад Культура поведения и общения
Культура поведения и общения Apģērbs. Apģērba materiāli
Apģērbs. Apģērba materiāli Модели экономического развития стран Азии
Модели экономического развития стран Азии Экспериментальные исследования характеристик электродугового плазмотрона коаксиального типа
Экспериментальные исследования характеристик электродугового плазмотрона коаксиального типа Фестиваль подводных профессий Погружение в Арктику
Фестиваль подводных профессий Погружение в Арктику Презентация на тему Общая физиология центральной нервной системы
Презентация на тему Общая физиология центральной нервной системы  3 день озон - статусы заказа (1)
3 день озон - статусы заказа (1)