- Модели молекулярной эволюции, кладистика по Хеннигу

Содержание
- 2. Модели молекулярной эволюции ДНК: 1 5 10 tagcaaaatg
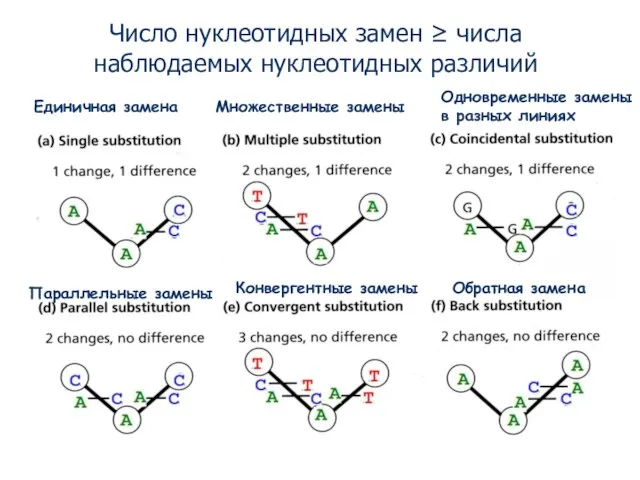
- 3. Соотношения между нуклеотидными заменами и нуклеотидными различиями Единичная замена Множественные замены Параллельные замены Конвергентные замены Обратная
- 4. Единичная замена Множественные замены Параллельные замены Конвергентные замены Обратная замена Одновременные замены в разных линиях Число
- 5. явные и скрытые генетические дистанции 1 10 20 30 Что такое генетическая дистанция? d = p,
- 6. Единичная замена Множественные замены Число нуклеотидных замен ≥ числа наблюдаемых нуклеотидных отличий Проблема дистанций состоит в
- 7. Наблюдаемые генетические дистанции как правило меньше реальных эволюционных дистанций, так как есть скрытые замены Но как
- 8. Закономерности накопления замен
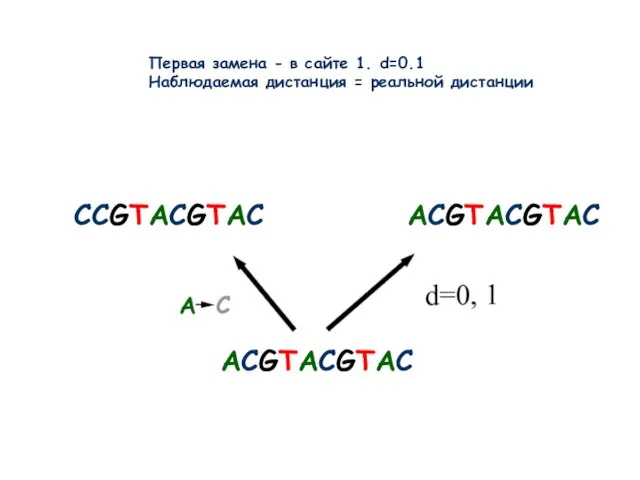
- 9. ACGTACGTAC CCGTACGTAC ACGTACGTAC Первая замена - в сайте 1. d=0.1 Наблюдаемая дистанция = реальной дистанции
- 10. CCGAACGTAC ACGTACGTAC Вторая замена – Имеется вероятность 0.1, что она будет повторной (т.е. тоже в сайте
- 11. CGTACGTACG ACGTACGTAC Третья замена имеет большую вероятность быть повторной, четвертая – еще большую, и. т.д. Т.е.
- 12. Зависимость между временем дивергенции и числом наблюдаемых нуклеотидных отличий в гене CytB у жвачных копытных животных
- 13. “Сырые” (нескорректированные) генетические дистанции легко вычислить, но они могут быть сильно занижены. Необходима коррекция Ее можно
- 14. Purines = adenin and guanine Pirimidines = cytosine and thymine
- 15. Кривые накопления повторных замен для транзиций и трансверсий Каждая точка – это сравнение, т.е. пара видов
- 16. Кривая накопления транзиций по отношению к трансверсиям
- 17. Генетический код Замена в первой позиции кодона ведет к замене аминокислоты Замена в третьей позиции кодона
- 18. Кривые накопления повторных замен для третьей и первой позиций кодона
- 19. Какие параметры можно извлечь из нуклеотидного выравнивания? 1 10 20 30
- 20. Какие параметры можно извлечь из нуклеотидного выравнивания? (1) Длина (2) Доля изменчивых сайтов (3) Доля инвариантных
- 21. Какие параметры можно извлечь из нуклеотидного выравнивания? Длина выравнивания Зависит от задач и технических возможностей 1
- 22. Какие параметры можно извлечь из нуклеотидного выравнивания? (2) Доля изменчивых сайтов (3) Доля инвариантных сайтов 1
- 23. Какие параметры можно извлечь из нуклеотидного выравнивания? (4) Соотношение нуклеотидов разных типов (A C G T)
- 24. Какие параметры можно извлечь из нуклеотидного выравнивания? (5) Доля транзиций и трансверсий 1 10 20 30
- 25. Какие параметры можно извлечь из нуклеотидного выравнивания? (6) Доля нуклеотидных замен разных типов 1 10 20
- 26. Какие параметры можно извлечь из нуклеотидного выравнивания? (7) Все это (2-6) отдельно для каждой позиции кодона
- 27. Какие параметры можно извлечь из нуклеотидного выравнивания? (8) Доля синонимичных и несинонимичных замен (9) Доля синонимичных
- 28. Предпосылки 1) нуклеотидные замены одного типа равновероятны в разных частях одного гена 2) нуклеотидные замены обратимы
- 29. Если вероятности нуклеотидных замен (p) и частоты нуклеотидов (f) константны во времени, то суммарная эволюционная дистанция
- 30. Если вероятности нуклеотидных замен (p) и частоты нуклеотидов (f) константны во времени, то суммарная эволюционная дистанция
- 31. частоты нуклеотидов и доли замен разного типа берутся непосредственно из выравнивания
- 32. JC Вероятности всех замен одинаковы, частоты нуклеотидов равны D= D = -(3/4)ln(1-4/3 где p – это
- 33. Двухпараметрическая модель Кимуры K2P Вероятности транзиций и трансверсий разные, частоты нуклеотидов равны α – транзиция β
- 34. F81 Вероятности всех замен одинаковы, но частоты нуклеотидов разные
- 35. HKY model Вероятности транзиций и трансверсий разные, частоты нуклеотидов разные
- 36. REV Вероятности ВСЕХ ЗАМЕН разные, частоты нуклеотидов разные
- 38. Чем хороши и чем плохи сложные и простые модели?
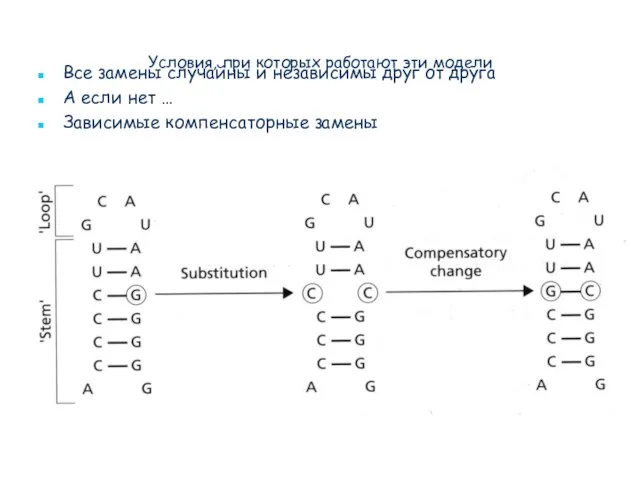
- 39. Условия, при которых работают эти модели Это стохастические модели, которые предполагают, что все замены случайны и
- 40. Структура 18S rDNA
- 41. 18S rDNA (фрагмент) 18S rDNA (фрагмент)
- 42. Условия, при которых работают эти модели Все замены случайны и независимы друг от друга А если
- 43. Общие принципы построения филогений 1) Анализ признаков, 2) выбор оптимальной модели эволюции признака, 3) выбор методов
- 44. Подходы к выявлению филогений традиционный (Геккелевский, эмпирико-интуитивный) традиционная кладистика (Hennig, 1950, 1966) фенетика метод максимальной парсимонии
- 45. Традиционный (эмпирико-интуитивный) метод выведения филогений Строго научный и, как правило, очень качественный анализ признаков сочетается с
- 46. По Геккелю филогенетика – наука о путях, закономерностях и причинах исторического развития организмов Ernst Haeckel (1834-1919)
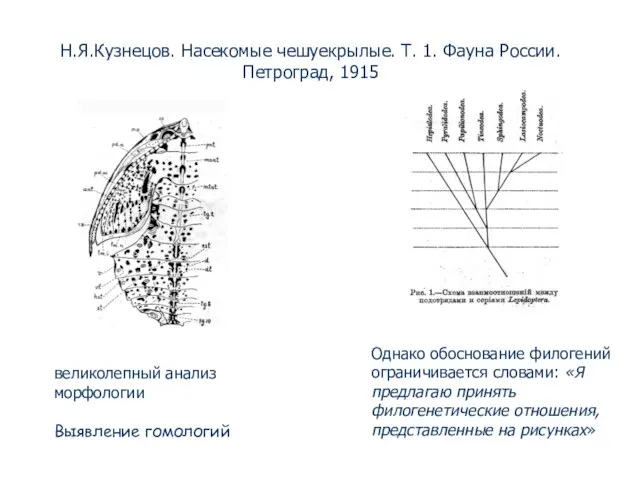
- 47. Н.Я.Кузнецов. Насекомые чешуекрылые. Т. 1. Фауна России. Петроград, 1915 Однако обоснование филогений ограничивается словами: «Я предлагаю
- 48. “Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими древесами, на которые я указал
- 49. Традиционная кладистика (Hennig, 1950, 1966) Хенниг предложил строго научные принципы перехода от анализа признаков к реконструкции
- 50. Признаки Негомологичные (гомоплазии) Гомологичные Плезиоморфии Апоморфии Синапоморфии
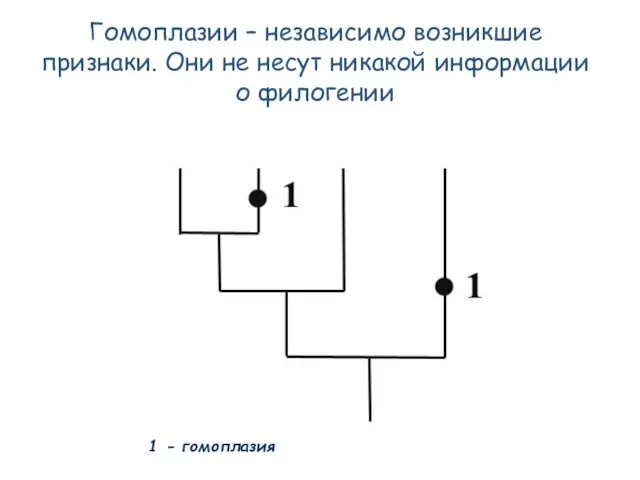
- 51. Гомоплазии – независимо возникшие признаки. Они не несут никакой информации о филогении 1 - гомоплазия
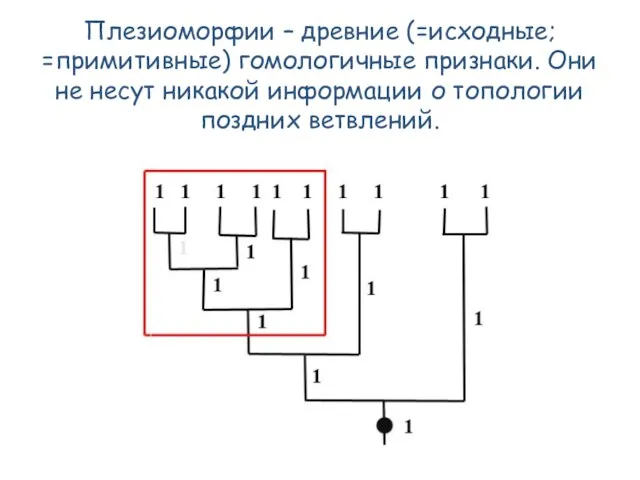
- 52. Плезиоморфии – древние (=исходные; =примитивные) гомологичные признаки. Они не несут никакой информации о топологии поздних ветвлений.
- 53. Апоморфия – новый (=продвинутый; =производный; =прогрессивынй) гомологичный признак. Единичная апоморфия, возникшая в концевой ветви, метит только
- 54. Но если апоморфия возникла до разделения ветвей и передалась в обе ветки, то наличие такой апоморфии
- 55. Для построения филогении трех таксонов (два ветвления) необходимо наличие одной синапоморфии Для построения филогении трех таксонов
- 56. В общем виде для полного разрешения филогении, включающей n ветвлений, необходимо и достаточно n-1 синапоморфий (по
- 57. Филогения строится как система соподчиненных (вложенных одна в другую) клад (монофилетических групп), каждая из которых выявляется
- 58. Модель эволюции в кладистике по Геннигу Топология - строгая дихотомия Процесс – накопление синапоморфий.
- 59. Алгоритм анализа Одна истинная синапоморфия может разрешить узел ветвления филогенетического дерева Выявление филогении – многоступенчатый процесс
- 60. Построение молекулярного дерева с использованием кладистики по Хеннигу 1 AAGT 2 AAGT 3 ACGT
- 61. Построение молекулярного дерева таксонов 1-4 с использованием кладистики по Хеннигу 1 AAGTT 2 AAGTT 3 ACGTT
- 62. Состояние ACGTA плезиоморфно 1 AAGTT 2 AAGTT 3 ACGTT 4 ACGTA 5 ACGTA 6 ACGTA 7
- 63. A во второй позиции – синапоморфия 1 + 2 1 AAGTT 2 AAGTT 3 ACGTT 4
- 64. T в пятой позиции – синапоморфия 1 + 2 +3 1 AAGTT 2 AAGTT 3 ACGTT
- 65. Проблема гомоплазий Презумпция: Синапоморфии встречаются чаще, чем гомоплазии
- 66. Конфликт между потенциальными синапоморфиями 1 AAGTT 2 AACTT 3 ACCTT 4 ACGTT
- 67. Принципы традиционной кладистики Если возникает конфликт между потенциальными синапоморфиями, то основной путь его решения – переисследование
- 68. Другие проблемы генниговской кладистики: “Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить все узлы
- 69. Проблемы традиционной кладистики “Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить все узлы ветвления
- 70. Проблемы традиционной кладистики “Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить все узлы ветвления
- 71. Картины филогенезов, которуе создает кладистический (по Геннигу и парсимониальный) анализ, неполны и однобоки: Анагенез не учитывается
- 72. Принцип монофилии лежит в самой основе алгоритма построения дерева в хенниговской кладистике. Сипапоморфии однозначно определяют только
- 73. Кладизм объявляет парафилетические группы вне закона просто по той причине, что он не умеет их выявлять
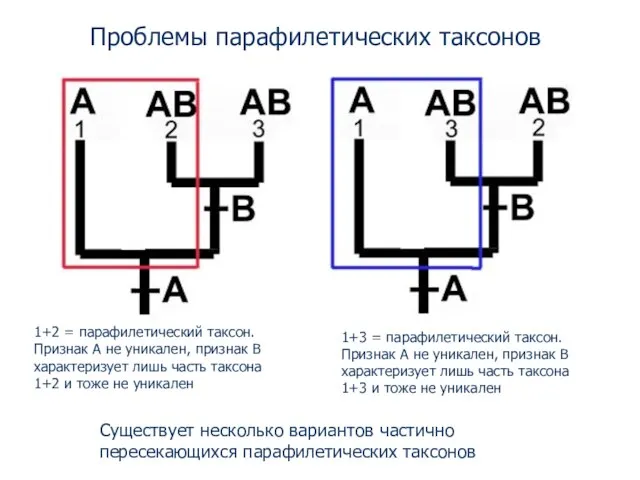
- 74. Проблемы парафилетических таксонов 1+2 = парафилетический таксон. Признак A не уникален, признак B характеризует лишь часть
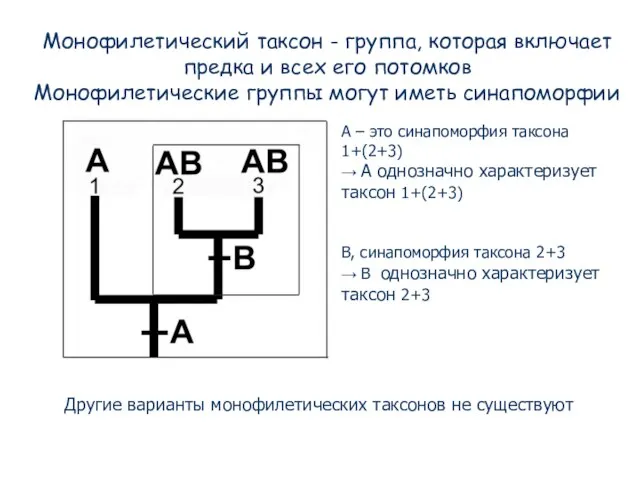
- 75. Монофилетический таксон - группа, которая включает предка и всех его потомков Монофилетические группы могут иметь синапоморфии
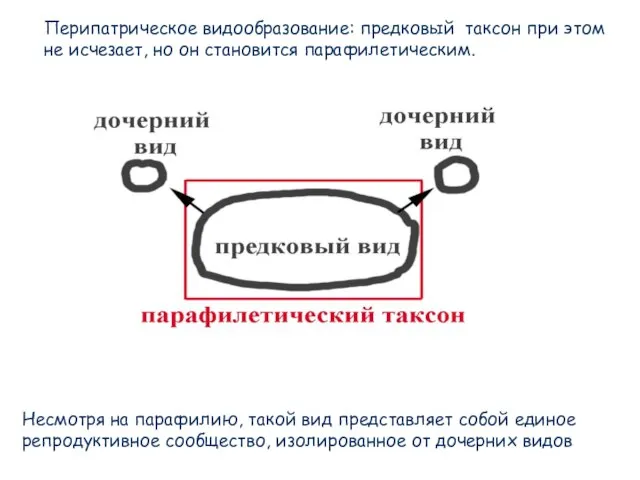
- 76. Перипатрическое видообразование: предковый таксон при этом не исчезает, но он становится парафилетическим. Несмотря на парафилию, такой
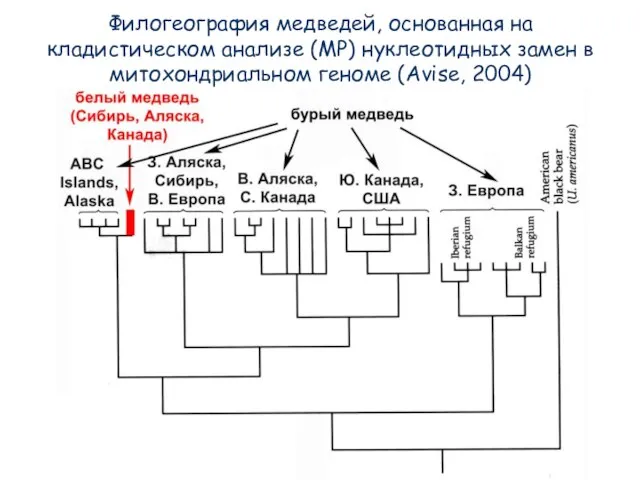
- 77. Филогеография медведей, основанная на кладистическом анализе (MP) нуклеотидных замен в митохондриальном геноме (Avise, 2004)
- 78. Кладистика по Геннигу остается рабочим инструментом филогенетики!
- 79. Фенетика В кладистике процедура выявления гомологичных признаков (дифференциация от гомоплазий) не формализована. Это может быть причиной
- 80. Фенетика Отказ от доминирования принципа гомологии (в фенетике все признаки имеют равный вес) Степень родства =
- 81. Фенетика Кластерный анализ (выявление группировок по степени их сходства). Иерархии таких группировок можно интерпретировать в качестве
- 82. Фенетика Пример научной, но неправильной (неадекватной) методологии Научность – строгое следование принципам научной логики, избегание субъективизма
- 83. Традиционная и нумерическая кладистика Увеличение числа признаков приводит к противоречиям между предполагаемыми синапоморфиями, которые свидетельствуют о
- 84. Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения: 1)переисследование материала, поиск и
- 85. Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения: 1)переисследование материала, поиск и
- 86. Нумерическая кладистика и метод максимальной парсимонии При наличии противоречий между “синапоморфиями” возможны разные варианты филогении Как
- 87. Метод максимальной парсимонии (наибольшей экономии)
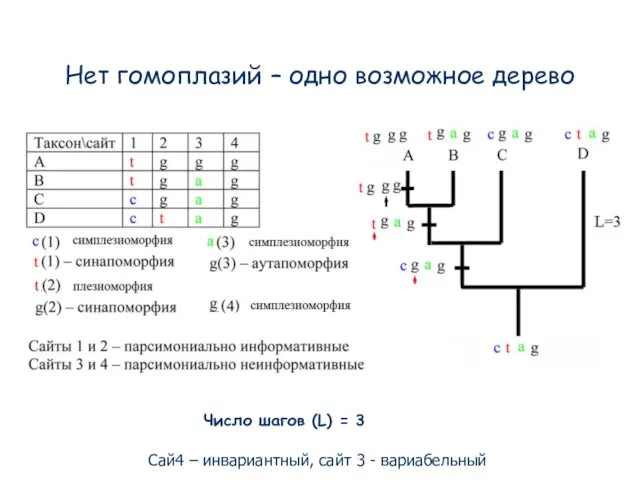
- 88. Нет гомоплазий – одно возможное дерево Число шагов (L) = 3 Сай4 – инвариантный, сайт 3
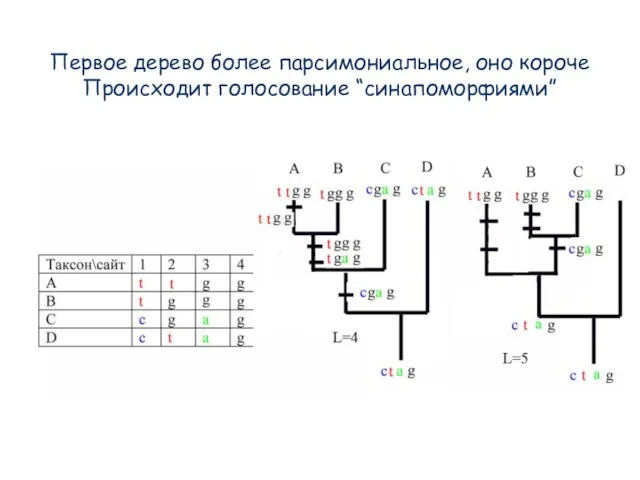
- 89. Первое дерево более парсимониальное, оно короче Происходит голосование “синапоморфиями”
- 90. в реальности у нас исходно нет ни топологии дерева, ни распределения признаков по нему, ни анцестрального
- 91. Шаг 1: выявление признаков и их состояний Признак – цвет глаз Состояния – коричневый, голубой, зеленый
- 92. Признак – цвет глаз Состояния – коричневый (0), голубой (1), зеленый (2) Признак – группа крови
- 93. Шаг 3: Составление матрицы признаков
- 94. Бинарная матрица Матрица множественных состояний
- 95. Нуклеотидное (или аминокислотное) выравнивание – это уже готовая матрица признаков 4 состояния – A C G
- 96. Шаг 4: выбор модели эволюции Модель Камина-Сокола (Camin- Sokal parsimony): анцестральное состояние известно, тогда 0 —›
- 97. Модель Долло (Dollo parsimony) (основана на принципе необратимости эволюции) - допускаются изменения признака в любую сторону,
- 98. Модель Фитча-Вагнера (Fitch-Wagner parsimony) – симметричная модель 0 дерево неукорененное!!!
- 99. Модель Фитча-Вагнера (Fitch-Wagner parsimony) для множественных состояний признака 0 0 0 1 1 2 дерево неукорененное!!!
- 101. Скачать презентацию

Слайд 3Соотношения между нуклеотидными
заменами и нуклеотидными различиями
Единичная замена
Множественные замены
Параллельные замены
Конвергентные замены
Обратная замена
Одновременные
Соотношения между нуклеотидными
заменами и нуклеотидными различиями
Единичная замена
Множественные замены
Параллельные замены
Конвергентные замены
Обратная замена
Одновременные

Слайд 4Единичная замена
Множественные замены
Параллельные замены
Конвергентные замены
Обратная замена
Одновременные замены
в разных линиях
Число нуклеотидных замен ≥
Единичная замена
Множественные замены
Параллельные замены
Конвергентные замены
Обратная замена
Одновременные замены
в разных линиях
Число нуклеотидных замен ≥
Слайд 5явные и скрытые генетические дистанции
1
10
20
30
Что такое генетическая дистанция?
d = p, где p
явные и скрытые генетические дистанции
1
10
20
30
Что такое генетическая дистанция?
d = p, где p

Слайд 6Единичная замена
Множественные замены
Число нуклеотидных замен ≥ числа наблюдаемых нуклеотидных отличий
Проблема дистанций состоит
Единичная замена
Множественные замены
Число нуклеотидных замен ≥ числа наблюдаемых нуклеотидных отличий
Проблема дистанций состоит
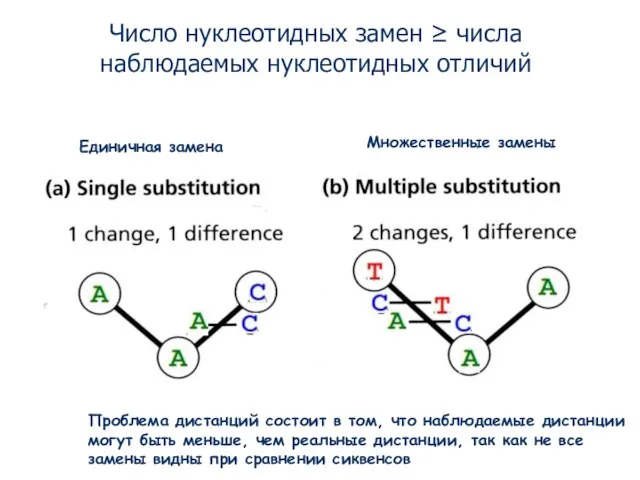
Слайд 7Наблюдаемые генетические дистанции как правило меньше реальных эволюционных дистанций, так как есть
Наблюдаемые генетические дистанции как правило меньше реальных эволюционных дистанций, так как есть

Слайд 8Закономерности накопления замен
Закономерности накопления замен
Слайд 9ACGTACGTAC
CCGTACGTAC
ACGTACGTAC
Первая замена - в сайте 1. d=0.1
Наблюдаемая дистанция = реальной дистанции
ACGTACGTAC
CCGTACGTAC
ACGTACGTAC
Первая замена - в сайте 1. d=0.1
Наблюдаемая дистанция = реальной дистанции
Слайд 10CCGAACGTAC
ACGTACGTAC
Вторая замена –
Имеется вероятность 0.1, что она будет повторной
(т.е. тоже
CCGAACGTAC
ACGTACGTAC
Вторая замена –
Имеется вероятность 0.1, что она будет повторной
(т.е. тоже

Слайд 11CGTACGTACG
ACGTACGTAC
Третья замена имеет большую вероятность быть повторной,
четвертая – еще большую, и.
CGTACGTACG
ACGTACGTAC
Третья замена имеет большую вероятность быть повторной,
четвертая – еще большую, и.

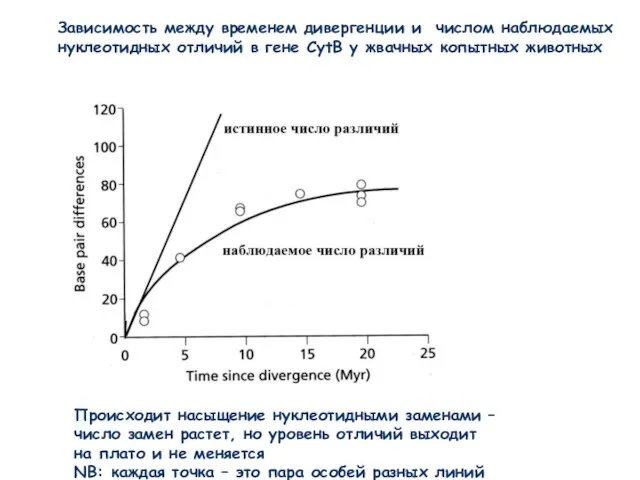
Слайд 12Зависимость между временем дивергенции и числом наблюдаемых
нуклеотидных отличий в гене CytB
Зависимость между временем дивергенции и числом наблюдаемых
нуклеотидных отличий в гене CytB
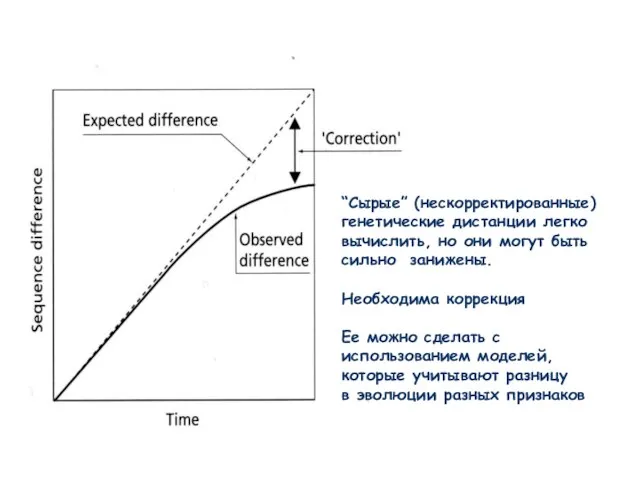
Слайд 13“Сырые” (нескорректированные)
генетические дистанции легко
вычислить, но они могут быть
сильно занижены.
Необходима коррекция
Ее
“Сырые” (нескорректированные)
генетические дистанции легко
вычислить, но они могут быть
сильно занижены.
Необходима коррекция
Ее
Слайд 14Purines = adenin and guanine
Pirimidines = cytosine and thymine
Purines = adenin and guanine
Pirimidines = cytosine and thymine
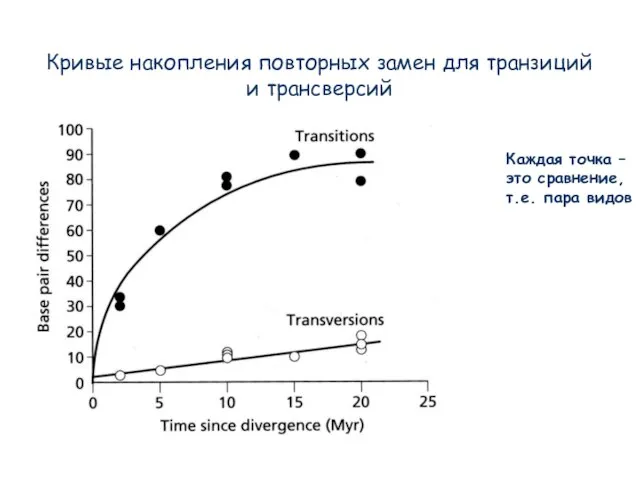
Слайд 15Кривые накопления повторных замен для транзиций и трансверсий
Каждая точка –
это сравнение,
т.е. пара
Кривые накопления повторных замен для транзиций и трансверсий
Каждая точка –
это сравнение,
т.е. пара
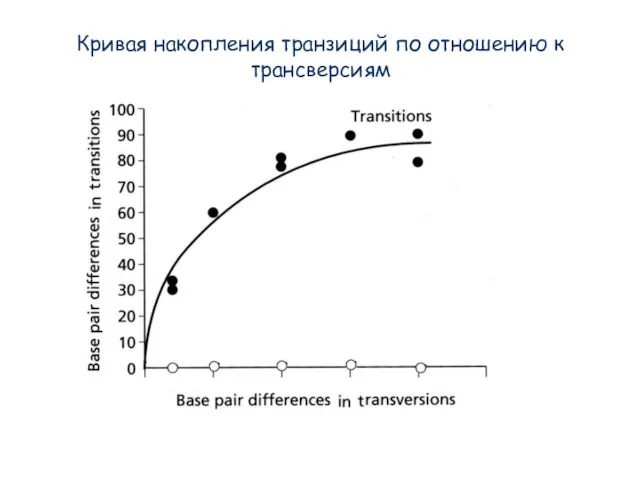
Слайд 16Кривая накопления транзиций по отношению к трансверсиям
Кривая накопления транзиций по отношению к трансверсиям
Слайд 17Генетический код
Замена в первой позиции кодона ведет к замене аминокислоты
Замена в третьей
Генетический код
Замена в первой позиции кодона ведет к замене аминокислоты
Замена в третьей

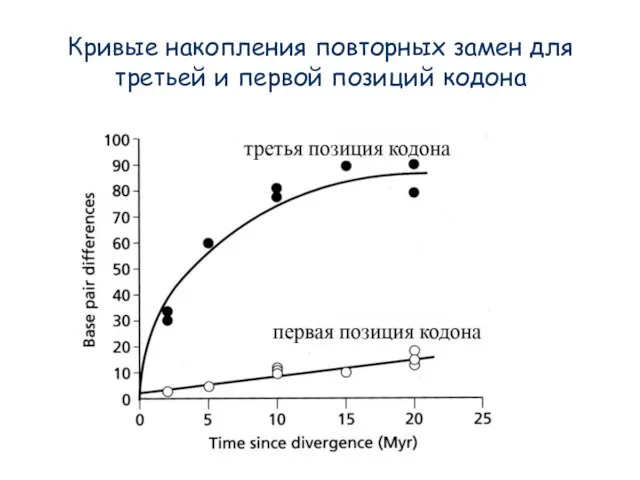
Слайд 18Кривые накопления повторных замен для третьей и первой позиций кодона
Кривые накопления повторных замен для третьей и первой позиций кодона
Слайд 19Какие параметры можно извлечь из нуклеотидного выравнивания?
1
10
20
30
Какие параметры можно извлечь из нуклеотидного выравнивания?
1
10
20
30

Слайд 20Какие параметры можно извлечь из нуклеотидного выравнивания?
(1) Длина
(2) Доля изменчивых сайтов
(3)
Какие параметры можно извлечь из нуклеотидного выравнивания?
(1) Длина
(2) Доля изменчивых сайтов
(3)

Слайд 21Какие параметры можно извлечь из нуклеотидного выравнивания?
Длина выравнивания
Зависит от задач и технических
Какие параметры можно извлечь из нуклеотидного выравнивания?
Длина выравнивания
Зависит от задач и технических

Слайд 22Какие параметры можно извлечь из нуклеотидного выравнивания?
(2) Доля изменчивых сайтов
(3) Доля
Какие параметры можно извлечь из нуклеотидного выравнивания?
(2) Доля изменчивых сайтов
(3) Доля

Слайд 23Какие параметры можно извлечь из нуклеотидного выравнивания?
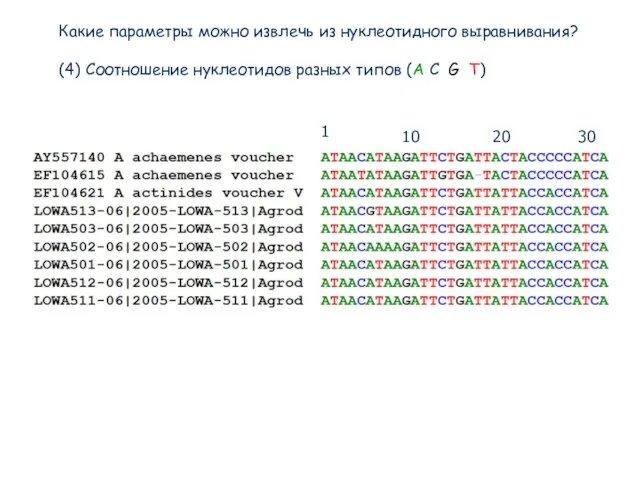
(4) Соотношение нуклеотидов разных типов (A
Какие параметры можно извлечь из нуклеотидного выравнивания?
(4) Соотношение нуклеотидов разных типов (A
Слайд 24Какие параметры можно извлечь из нуклеотидного выравнивания?
(5) Доля транзиций и трансверсий
1
10
20
30
Какие параметры можно извлечь из нуклеотидного выравнивания?
(5) Доля транзиций и трансверсий
1
10
20
30

Слайд 25Какие параметры можно извлечь из нуклеотидного выравнивания?
(6) Доля нуклеотидных замен разных типов
1
10
20
30
Какие параметры можно извлечь из нуклеотидного выравнивания?
(6) Доля нуклеотидных замен разных типов
1
10
20
30

Слайд 26Какие параметры можно извлечь из нуклеотидного выравнивания?
(7) Все это (2-6) отдельно для
Какие параметры можно извлечь из нуклеотидного выравнивания?
(7) Все это (2-6) отдельно для

Слайд 27Какие параметры можно извлечь из нуклеотидного выравнивания?
(8) Доля синонимичных и несинонимичных замен
(9)
Какие параметры можно извлечь из нуклеотидного выравнивания?
(8) Доля синонимичных и несинонимичных замен
(9)

Слайд 28Предпосылки
1) нуклеотидные замены одного типа равновероятны в разных частях одного гена
2) нуклеотидные
Предпосылки
1) нуклеотидные замены одного типа равновероятны в разных частях одного гена
2) нуклеотидные

Слайд 29Если вероятности нуклеотидных замен (p) и частоты нуклеотидов (f) константны во времени,
Если вероятности нуклеотидных замен (p) и частоты нуклеотидов (f) константны во времени,

Слайд 30Если вероятности нуклеотидных замен (p) и частоты нуклеотидов (f) константны во времени,
Если вероятности нуклеотидных замен (p) и частоты нуклеотидов (f) константны во времени,

Слайд 31 частоты нуклеотидов и доли замен разного типа берутся непосредственно из выравнивания
частоты нуклеотидов и доли замен разного типа берутся непосредственно из выравнивания

Слайд 32JC
Вероятности всех замен одинаковы, частоты нуклеотидов равны
D=
D = -(3/4)ln(1-4/3
где p – это
JC
Вероятности всех замен одинаковы, частоты нуклеотидов равны
D=
D = -(3/4)ln(1-4/3
где p – это

Слайд 33Двухпараметрическая модель Кимуры K2P
Вероятности транзиций и трансверсий разные,
частоты нуклеотидов равны
α –
Двухпараметрическая модель Кимуры K2P
Вероятности транзиций и трансверсий разные,
частоты нуклеотидов равны
α –

Слайд 34 F81
Вероятности всех замен одинаковы, но частоты нуклеотидов разные
F81
Вероятности всех замен одинаковы, но частоты нуклеотидов разные

Слайд 35
HKY model
Вероятности транзиций и трансверсий разные,
частоты нуклеотидов разные
HKY model
Вероятности транзиций и трансверсий разные,
частоты нуклеотидов разные

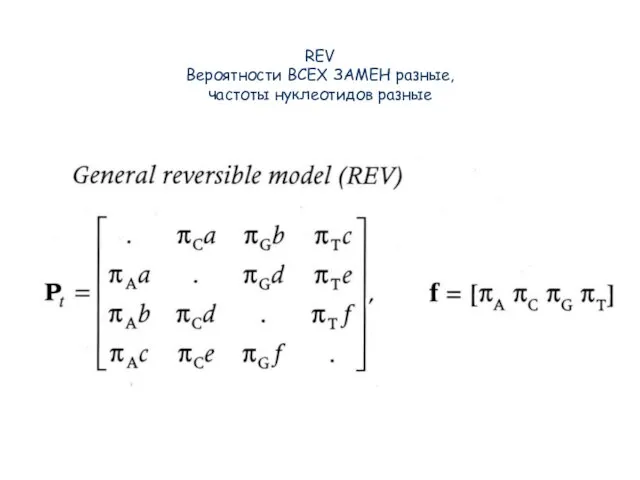
Слайд 36REV
Вероятности ВСЕХ ЗАМЕН разные,
частоты нуклеотидов разные
REV
Вероятности ВСЕХ ЗАМЕН разные,
частоты нуклеотидов разные
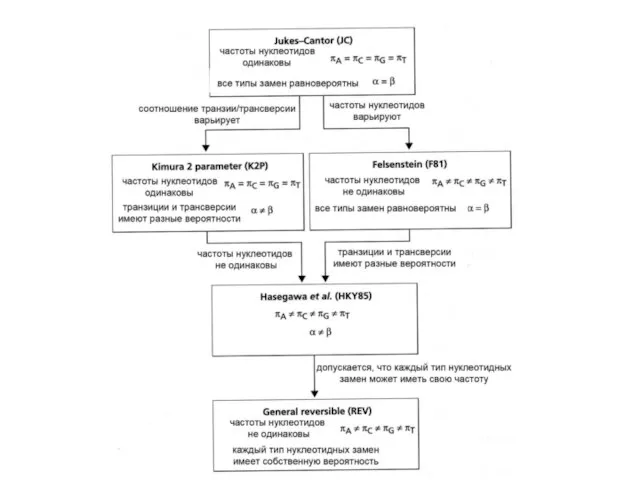
Слайд 38Чем хороши и чем плохи сложные и простые модели?
Чем хороши и чем плохи сложные и простые модели?
Слайд 39Условия, при которых работают эти модели
Это стохастические модели, которые предполагают, что все
Условия, при которых работают эти модели
Это стохастические модели, которые предполагают, что все

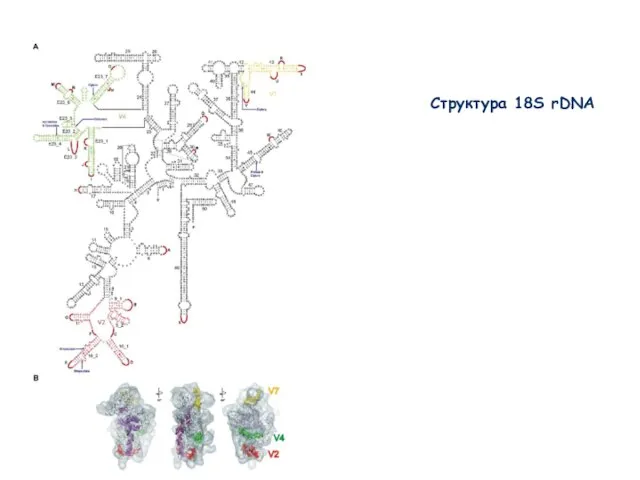
Слайд 40Структура 18S rDNA
Структура 18S rDNA
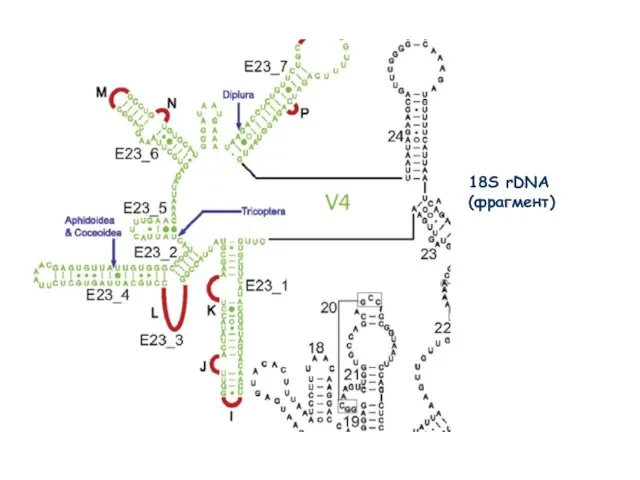
Слайд 4118S rDNA
(фрагмент)
18S rDNA
(фрагмент)
18S rDNA
(фрагмент)
18S rDNA
(фрагмент)
Слайд 42Условия, при которых работают эти модели
Все замены случайны и независимы друг от
Условия, при которых работают эти модели
Все замены случайны и независимы друг от
Слайд 43
Общие принципы построения филогений
1) Анализ признаков,
2) выбор оптимальной модели
Общие принципы построения филогений
1) Анализ признаков,
2) выбор оптимальной модели

Слайд 44Подходы к выявлению филогений
традиционный (Геккелевский, эмпирико-интуитивный)
традиционная кладистика (Hennig, 1950, 1966)
фенетика
метод
Подходы к выявлению филогений
традиционный (Геккелевский, эмпирико-интуитивный)
традиционная кладистика (Hennig, 1950, 1966)
фенетика
метод

Слайд 45Традиционный (эмпирико-интуитивный) метод выведения филогений
Строго научный и, как правило, очень качественный анализ
Традиционный (эмпирико-интуитивный) метод выведения филогений
Строго научный и, как правило, очень качественный анализ

Слайд 46По Геккелю филогенетика – наука о путях,
закономерностях и причинах исторического
развития
По Геккелю филогенетика – наука о путях,
закономерностях и причинах исторического
развития

Слайд 47Н.Я.Кузнецов. Насекомые чешуекрылые. Т. 1. Фауна России. Петроград, 1915
Однако обоснование филогений ограничивается
Н.Я.Кузнецов. Насекомые чешуекрылые. Т. 1. Фауна России. Петроград, 1915
Однако обоснование филогений ограничивается
Слайд 48 “Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими древесами,
“Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими древесами,
![“Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими древесами,](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/367055/slide-47.jpg)
Слайд 49
Традиционная кладистика (Hennig, 1950, 1966)
Хенниг предложил строго научные принципы перехода от анализа
Традиционная кладистика (Hennig, 1950, 1966) Хенниг предложил строго научные принципы перехода от анализа

Слайд 50Признаки
Негомологичные (гомоплазии)
Гомологичные
Плезиоморфии
Апоморфии
Синапоморфии
Признаки
Негомологичные (гомоплазии)
Гомологичные
Плезиоморфии
Апоморфии
Синапоморфии

Слайд 51Гомоплазии – независимо возникшие признаки. Они не несут никакой информации о филогении
1
Гомоплазии – независимо возникшие признаки. Они не несут никакой информации о филогении
1
Слайд 52Плезиоморфии – древние (=исходные; =примитивные) гомологичные признаки. Они не несут никакой информации
Плезиоморфии – древние (=исходные; =примитивные) гомологичные признаки. Они не несут никакой информации
Слайд 53Апоморфия – новый
(=продвинутый; =производный; =прогрессивынй)
гомологичный признак.
Единичная апоморфия, возникшая в
Апоморфия – новый
(=продвинутый; =производный; =прогрессивынй)
гомологичный признак.
Единичная апоморфия, возникшая в
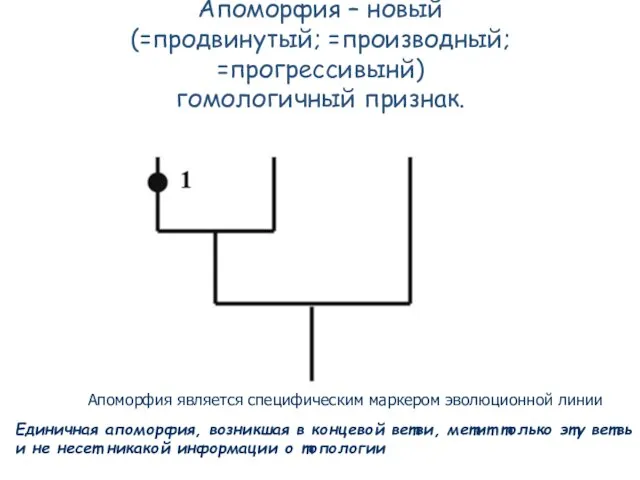
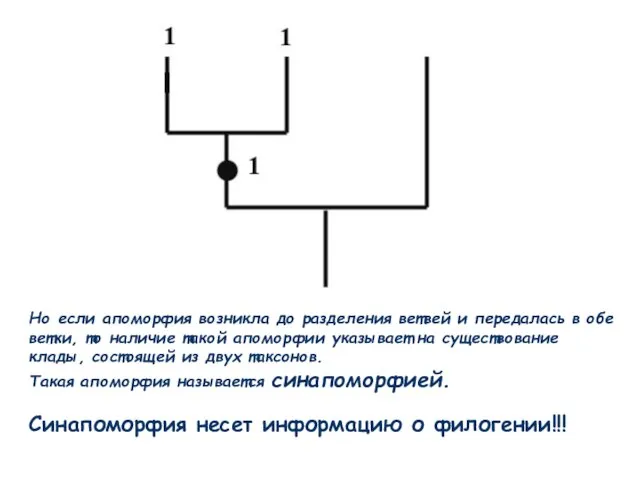
Слайд 54Но если апоморфия возникла до разделения ветвей и передалась в обе ветки,
Но если апоморфия возникла до разделения ветвей и передалась в обе ветки,
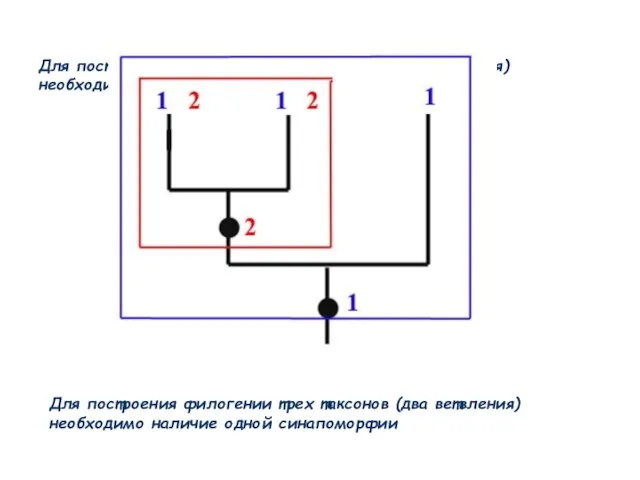
Слайд 55Для построения филогении трех таксонов (два ветвления) необходимо наличие одной синапоморфии
Для построения
Для построения филогении трех таксонов (два ветвления) необходимо наличие одной синапоморфии
Для построения
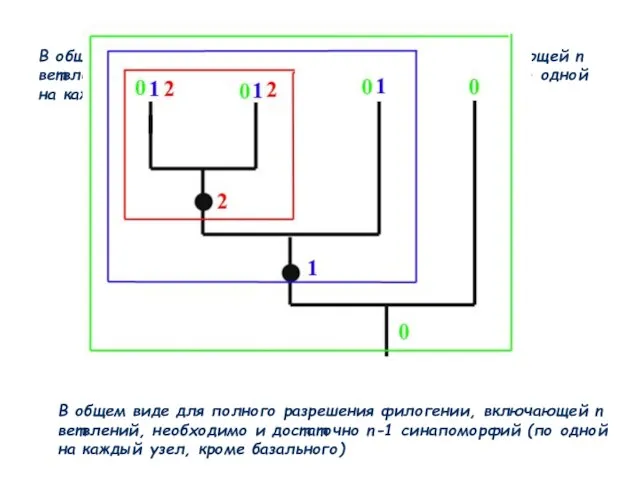
Слайд 56В общем виде для полного разрешения филогении, включающей n ветвлений, необходимо и
В общем виде для полного разрешения филогении, включающей n ветвлений, необходимо и
Слайд 57Филогения строится как система соподчиненных (вложенных одна в другую) клад (монофилетических групп),
Филогения строится как система соподчиненных (вложенных одна в другую) клад (монофилетических групп),

Слайд 58Модель эволюции в кладистике по Геннигу
Топология - строгая дихотомия
Процесс – накопление синапоморфий.
Модель эволюции в кладистике по Геннигу
Топология - строгая дихотомия
Процесс – накопление синапоморфий.

Слайд 59Алгоритм анализа
Одна истинная синапоморфия может разрешить узел ветвления филогенетического дерева
Выявление филогении –
Алгоритм анализа
Одна истинная синапоморфия может разрешить узел ветвления филогенетического дерева
Выявление филогении –

Слайд 60Построение молекулярного дерева с использованием кладистики по Хеннигу
1 AAGT
2 AAGT
3 ACGT
Построение молекулярного дерева с использованием кладистики по Хеннигу
1 AAGT
2 AAGT
3 ACGT

Слайд 61Построение молекулярного дерева таксонов 1-4 с использованием кладистики по Хеннигу
1 AAGTT
2 AAGTT
3
Построение молекулярного дерева таксонов 1-4 с использованием кладистики по Хеннигу
1 AAGTT
2 AAGTT
3

Слайд 62Состояние ACGTA плезиоморфно
1 AAGTT
2 AAGTT
3 ACGTT
4 ACGTA
5 ACGTA
6 ACGTA
7 ACGTA
Состояние ACGTA плезиоморфно
1 AAGTT
2 AAGTT
3 ACGTT
4 ACGTA
5 ACGTA
6 ACGTA
7 ACGTA

Слайд 63A во второй позиции – синапоморфия 1 + 2
1 AAGTT
2 AAGTT
3 ACGTT
4
A во второй позиции – синапоморфия 1 + 2
1 AAGTT
2 AAGTT
3 ACGTT
4

Слайд 64T в пятой позиции –
синапоморфия 1 + 2 +3
1 AAGTT
2 AAGTT
3
T в пятой позиции –
синапоморфия 1 + 2 +3
1 AAGTT
2 AAGTT
3

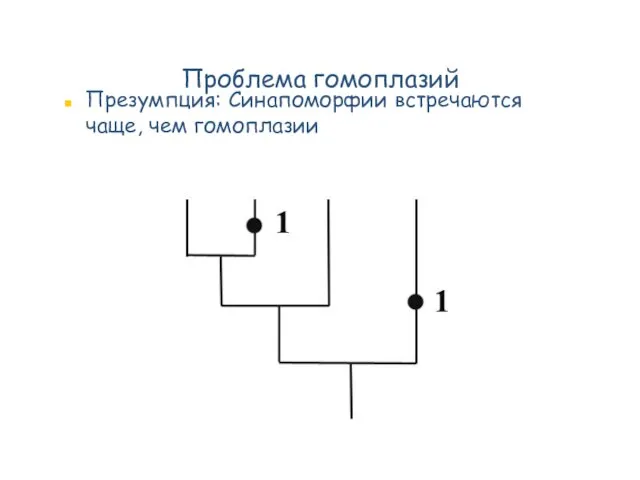
Слайд 65Проблема гомоплазий
Презумпция: Синапоморфии встречаются чаще, чем гомоплазии
Проблема гомоплазий
Презумпция: Синапоморфии встречаются чаще, чем гомоплазии
Слайд 66Конфликт между потенциальными синапоморфиями
1 AAGTT
2 AACTT
3 ACCTT
4 ACGTT
Конфликт между потенциальными синапоморфиями
1 AAGTT
2 AACTT
3 ACCTT
4 ACGTT

Слайд 67Принципы традиционной кладистики
Если возникает конфликт между потенциальными синапоморфиями, то основной путь его
Принципы традиционной кладистики
Если возникает конфликт между потенциальными синапоморфиями, то основной путь его

Слайд 68Другие проблемы генниговской кладистики:
“Надежных” синапоморфий может быть мало, недостаточно для того, что
Другие проблемы генниговской кладистики:
“Надежных” синапоморфий может быть мало, недостаточно для того, что

Слайд 69Проблемы традиционной кладистики
“Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить
Проблемы традиционной кладистики
“Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить

Слайд 70Проблемы традиционной кладистики
“Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить
Проблемы традиционной кладистики
“Надежных” синапоморфий может быть мало, недостаточно для того, что разрешить

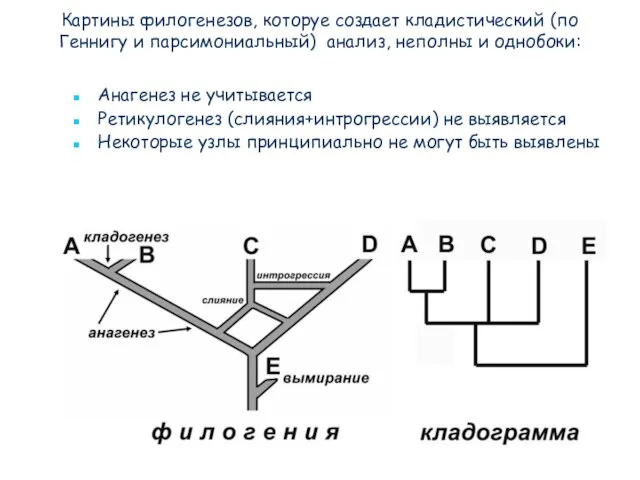
Слайд 71Картины филогенезов, которуе создает кладистический (по Геннигу и парсимониальный) анализ, неполны и
Картины филогенезов, которуе создает кладистический (по Геннигу и парсимониальный) анализ, неполны и
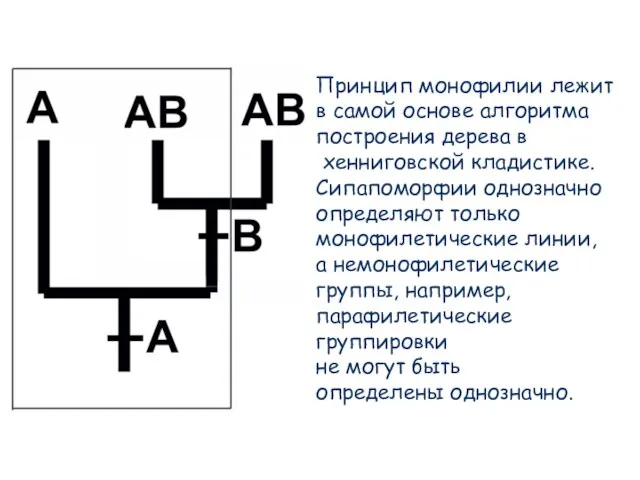
Слайд 72Принцип монофилии лежит
в самой основе алгоритма
построения дерева в
хенниговской кладистике.
Принцип монофилии лежит
в самой основе алгоритма
построения дерева в
хенниговской кладистике.
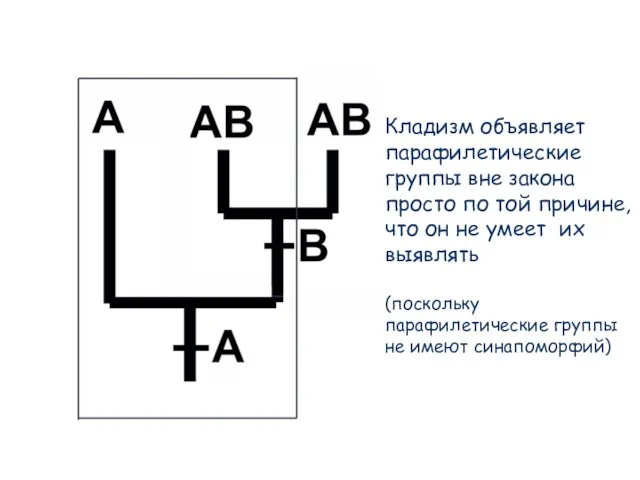
Слайд 73Кладизм объявляет парафилетические группы вне закона просто по той причине, что он
Кладизм объявляет парафилетические группы вне закона просто по той причине, что он
Слайд 74Проблемы парафилетических таксонов
1+2 = парафилетический таксон. Признак A не уникален, признак B
Проблемы парафилетических таксонов
1+2 = парафилетический таксон. Признак A не уникален, признак B
Слайд 75Монофилетический таксон - группа, которая включает предка и всех его потомков
Монофилетические
Монофилетический таксон - группа, которая включает предка и всех его потомков Монофилетические
Слайд 76Перипатрическое видообразование: предковый таксон при этом не исчезает, но он становится парафилетическим.
Перипатрическое видообразование: предковый таксон при этом не исчезает, но он становится парафилетическим.
Слайд 77Филогеография медведей, основанная на кладистическом анализе (MP) нуклеотидных замен в митохондриальном геноме
Филогеография медведей, основанная на кладистическом анализе (MP) нуклеотидных замен в митохондриальном геноме
Слайд 78Кладистика по Геннигу остается рабочим инструментом филогенетики!
Кладистика по Геннигу остается рабочим инструментом филогенетики!

Слайд 79Фенетика
В кладистике процедура выявления гомологичных признаков (дифференциация от гомоплазий) не формализована. Это
Фенетика
В кладистике процедура выявления гомологичных признаков (дифференциация от гомоплазий) не формализована. Это

Слайд 80Фенетика
Отказ от доминирования принципа гомологии (в фенетике все признаки имеют равный вес)
Степень
Фенетика
Отказ от доминирования принципа гомологии (в фенетике все признаки имеют равный вес)
Степень

Слайд 81Фенетика
Кластерный анализ (выявление группировок по степени их сходства).
Иерархии таких группировок можно
Фенетика
Кластерный анализ (выявление группировок по степени их сходства).
Иерархии таких группировок можно

Слайд 82Фенетика
Пример научной, но неправильной (неадекватной) методологии
Научность – строгое следование принципам научной
Фенетика
Пример научной, но неправильной (неадекватной) методологии
Научность – строгое следование принципам научной

Слайд 83Традиционная и нумерическая кладистика
Увеличение числа признаков приводит к противоречиям между предполагаемыми синапоморфиями,
Традиционная и нумерическая кладистика
Увеличение числа признаков приводит к противоречиям между предполагаемыми синапоморфиями,

Слайд 84Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения:
Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения:

Слайд 85Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения:
Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения:

Слайд 86Нумерическая кладистика и метод максимальной парсимонии
При наличии противоречий между “синапоморфиями” возможны разные
Нумерическая кладистика и метод максимальной парсимонии
При наличии противоречий между “синапоморфиями” возможны разные

Слайд 87
Метод максимальной парсимонии
(наибольшей экономии)
Метод максимальной парсимонии
(наибольшей экономии)

Слайд 88Нет гомоплазий – одно возможное дерево
Число шагов (L) = 3
Сай4 – инвариантный,
Нет гомоплазий – одно возможное дерево
Число шагов (L) = 3
Сай4 – инвариантный,
Слайд 89Первое дерево более парсимониальное, оно короче
Происходит голосование “синапоморфиями”
Первое дерево более парсимониальное, оно короче
Происходит голосование “синапоморфиями”
Слайд 90 в реальности у нас исходно нет ни топологии дерева, ни распределения
в реальности у нас исходно нет ни топологии дерева, ни распределения

Слайд 91Шаг 1: выявление признаков и их состояний
Признак – цвет глаз
Состояния – коричневый,
Шаг 1: выявление признаков и их состояний
Признак – цвет глаз
Состояния – коричневый,

Слайд 92Признак – цвет глаз
Состояния – коричневый (0), голубой (1), зеленый (2)
Признак –
Признак – цвет глаз
Состояния – коричневый (0), голубой (1), зеленый (2)
Признак –

Слайд 93Шаг 3: Составление матрицы признаков
Шаг 3: Составление матрицы признаков

Слайд 94Бинарная матрица
Матрица множественных состояний
Бинарная матрица
Матрица множественных состояний

Слайд 95 Нуклеотидное (или аминокислотное) выравнивание – это уже готовая матрица признаков
4 состояния
Нуклеотидное (или аминокислотное) выравнивание – это уже готовая матрица признаков
4 состояния

Слайд 96Шаг 4: выбор модели эволюции
Модель Камина-Сокола (Camin- Sokal parsimony): анцестральное состояние известно,
Шаг 4: выбор модели эволюции
Модель Камина-Сокола (Camin- Sokal parsimony): анцестральное состояние известно,

Слайд 97Модель Долло (Dollo parsimony) (основана на принципе необратимости эволюции) - допускаются изменения
Модель Долло (Dollo parsimony) (основана на принципе необратимости эволюции) - допускаются изменения

Слайд 98 Модель Фитча-Вагнера (Fitch-Wagner parsimony) – симметричная модель
0 <—› 1
дерево неукорененное!!!
Модель Фитча-Вагнера (Fitch-Wagner parsimony) – симметричная модель
0 <—› 1
дерево неукорененное!!!

Слайд 99 Модель Фитча-Вагнера (Fitch-Wagner parsimony) для множественных состояний признака
0 <—› 1
0
Модель Фитча-Вагнера (Fitch-Wagner parsimony) для множественных состояний признака
0 <—› 1
0

 Презентация1 (1)
Презентация1 (1) Школа эффективного бюджета студенческого самоуправления
Школа эффективного бюджета студенческого самоуправления Земля-шарообразное тело
Земля-шарообразное тело Проект создания «Музея истории информатики и вычислительной техники»
Проект создания «Музея истории информатики и вычислительной техники» Ребусы
Ребусы Artisman. Техническое обслуживание
Artisman. Техническое обслуживание Организационно-технологическое сопровождение ГИА – 9 в новой форме в 2012 г.
Организационно-технологическое сопровождение ГИА – 9 в новой форме в 2012 г. МЕЖДУНАРОДНАЯ ВЫСТАВКА ПО АСТРОНОМИИ «ВСЕЛЕННАЯ В ЦЕНТРЕ ГОРОДА» Как все начиналось…
МЕЖДУНАРОДНАЯ ВЫСТАВКА ПО АСТРОНОМИИ «ВСЕЛЕННАЯ В ЦЕНТРЕ ГОРОДА» Как все начиналось… Портрет слова
Портрет слова USB зажигалки
USB зажигалки Коса - девичья краса
Коса - девичья краса ЛОГИКА И КОМПЬЮТЕР.
ЛОГИКА И КОМПЬЮТЕР. Профессиональная деятельность учителя ИКН, марийского языка и литературы МОУ «Кукмаринская средняя общеобразовательная школа»
Профессиональная деятельность учителя ИКН, марийского языка и литературы МОУ «Кукмаринская средняя общеобразовательная школа» Агентство экономической диверсификации западных провинций Канады Western Economic Diversification Canada Автор доклада: Даг Мейли, Заместитель ми
Агентство экономической диверсификации западных провинций Канады Western Economic Diversification Canada Автор доклада: Даг Мейли, Заместитель ми Анализ TMS - решений и трейсинга для FTL
Анализ TMS - решений и трейсинга для FTL Салфетка. Вышивка
Салфетка. Вышивка День российской науки
День российской науки Информационное обслуживание пассажиров при перевозках на автомобильном транспорте
Информационное обслуживание пассажиров при перевозках на автомобильном транспорте Познание окружающего мира через экспериментирование
Познание окружающего мира через экспериментирование Основные шаги в русском танце
Основные шаги в русском танце Тематический натюрморт
Тематический натюрморт Динамика развития бизнеса финансовых услуг.
Динамика развития бизнеса финансовых услуг. Радио Принцип работы На передающей стороне
Радио Принцип работы На передающей стороне  Итоговая аттестация выпускников 9-х классов
Итоговая аттестация выпускников 9-х классов Явление закрытого жилого пространства в Санкт-Петербурге
Явление закрытого жилого пространства в Санкт-Петербурге Машины для послеуборочной обработки зерна
Машины для послеуборочной обработки зерна Использование средствMS Officeдля создания электронного учебника
Использование средствMS Officeдля создания электронного учебника Гражданское общество - понятие, функции
Гражданское общество - понятие, функции