- Молекулярные основы наследственности

Содержание
- 2. План лекции: Нуклеиновые кислоты – классификация, строение, функции. Макромолекулярная структура ДНК РНК: виды, структура, функции 4.Центральная
- 3. Нуклеиновые кислоты (НК) Два вида НК: ДНК (хранение наследственной информации) РНК (реализация наследственной информации)
- 4. Нуклеиновые кислоты (НК) Н.К.- биологические полимеры Мономеры- нуклеотиды
- 5. Состав нуклеотида
- 7. Нуклеозиды
- 8. Нуклеотиды
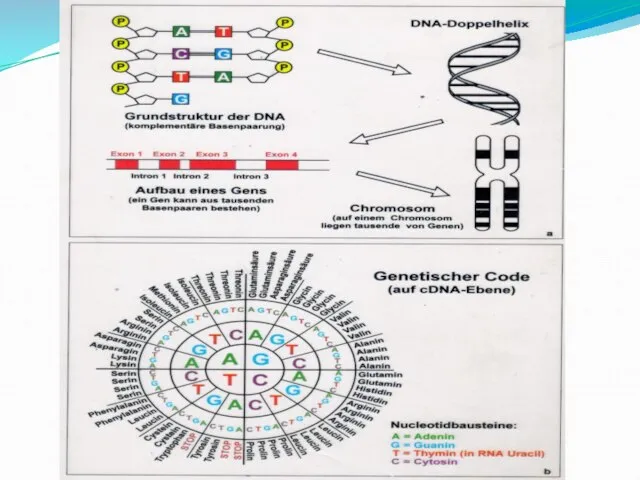
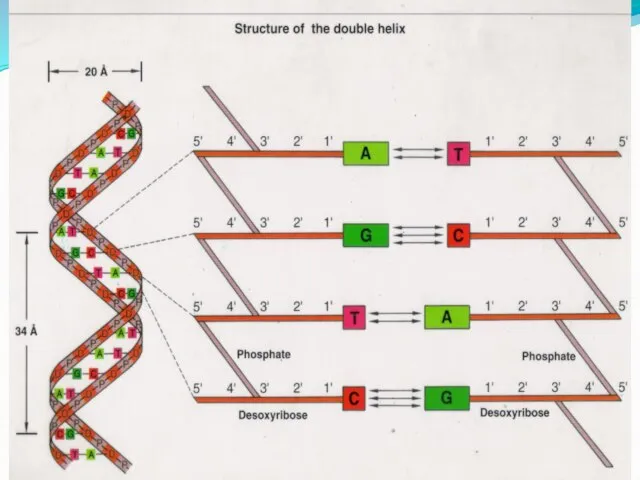
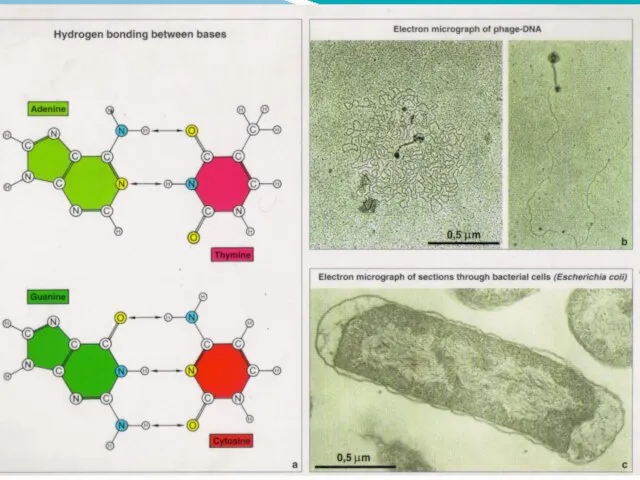
- 9. Модель Уотсона-Крика (1953) 1. ДНК-полимер из нуклеотидов, соединенных 3-5 фосфодиэфирными связями Состав нуклеотидов ДНК подчиняется правилам
- 12. Френсис Крик (1916-2004 гг) Джеймс Уотсон (р. 1928 г.) Нобелевская премия (1962 г.)
- 14. ДНК При горизонтальном изображении: 5---АТТГАЦАГГЦ---3 3---ТААЦТГТЦЦГ---5 В ядре человеческой клетки – 46 молекул ДНК Общая длина
- 15. Разнообразие форм ДНК Не все ДНК являются двуцепочечными. Геномы некоторых мелких вирусов бактерий, растений и животных
- 16. Формы ДНК В-форма (правозакрученная спираль)- основная форма существования ДНК Z –форма (левозакрученная спираль), содержит последовательности Г-Ц
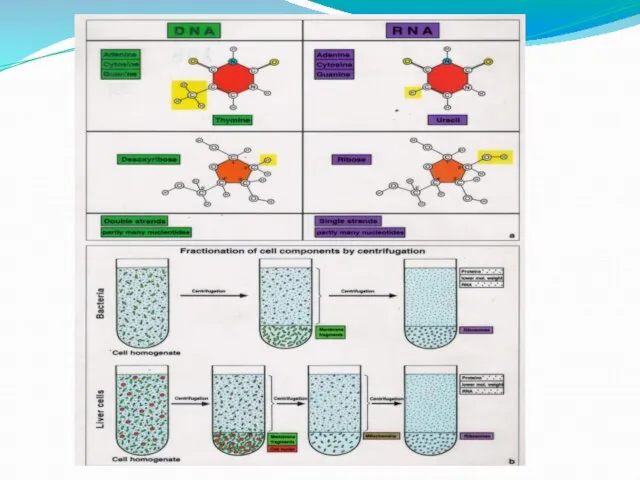
- 19. РНК: структура и функции Р-РНК(80-85%) Т-РНК (около10%) М(И)-РНК (5%) Мя-РНК (2%)
- 20. Р-РНК Р-РНК-структурная основа рибосом Р-РНК взаимодействуют с м-РНК и аминоацил-тРНК в процессе трансляции Это стабильные, нерастворимые
- 21. М-РНК М-РНК несет информацию о синтезе белка на ней самой, а также информацию о времени, количестве,
- 22. Строение М-РНК 1.Информативная зона - транслируемая область, работает как матрица в процессе биосинтеза белка 2.Неинформативная зона:
- 23. Строение М-РНК КЭП 5’-НТО AUG ТРАНСЛИРУЕМАЯ ОБЛАСТЬ СТОП 3’-НТО ПОЛИ (А) КЭП (от англ.сар- кепка,шапка )-
- 24. Т-РНК Т-РНК переносят аминокислоты в белоксинтезирующий аппарат клетки и выступают в роли затравки (праймера) в процессе
- 25. Т-РНК, особенности 1.По сравнению с р-РНК и м-РНК имеют меньшие размеры 2.Соотношение А:У и Г:Ц близко
- 26. Т-РНК Содержит в своей структуре: 1.Сайт прикрепления аминокислоты (акцепторный конец) 2.Сайт связывания с рибосомой 3.Антикодон
- 34. Постулат Крика Центральная догма молекулярной биологии: передача наследственной информации происходит только в одном направлении – от
- 35. Типы переноса наследственной информации Б) РНК---ДНК (обратная транскрипция). Происходит в клетках животных, зараженных вирусами определенного типа
- 36. Типы переноса наследственной информации 3. Запрещенный перенос (это процессы не зарегистрированные нигде и никогда). Это: Белок---ДНК,
- 37. Репликация ДНК – процесс самоудвоения, самовоспроизведения, самокопирования наследственной информации. Суть репликации ДНК - образование идентичных копий
- 38. Репликация ДНК идет на основе следующих принципов: Полуконсервативность Синтез ДНК начинается с расхождения цепей, каждая из
- 39. Консервативный способ репликации – когда после удвоения одна молекула состоит из двух старых цепей, другая –
- 40. Комплементарность Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В состав растущей цепи включается
- 42. Участок ДНК в пределах которого репликация начинается и заканчивается называется репликоном. В репликоне различают точку начала
- 43. Молекула ДНК, вступающая в репликацию: Точка, в которой происходит репликация называется репликационной вилкой (иногда наз. точкой
- 44. Репликация может идти либо в одном направлении, либо в двух направлениях. Это зависит от количества репликационных
- 45. Область, которая уже реплицирована, имеет вид «глазка» внутри нереплицировавшейся ДНК. Этот глазок выглядит одинаково при однонаправленной
- 46. При однонаправленной репликации глазок имеет фиксированную точку начала и движущуюся репликационную вилку. При двунаправленной репликации он
- 47. Этапы репликации: 1.Инициация идет с участием белков и ферментов, которые должны обеспечить: 1)Раскручивание ДНК 2)Связь инициирующих
- 48. Точки начала репликации богаты парами А-Т. Геликаза (от helix - спираль) расплетает двойную цепь родительской ДНК
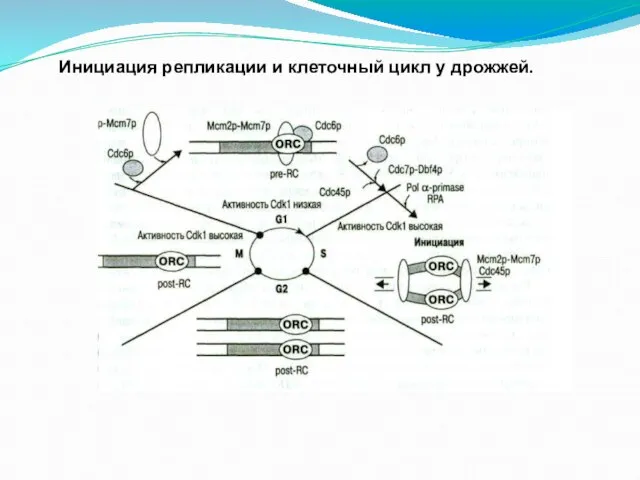
- 49. Инициация репликации и клеточный цикл у дрожжей.
- 50. Инициация репликации у эукариот. Белки инициации и происходящие процессы сходны с прокариотами. Отличия: 1.Участие дополнительного белка
- 51. 2. Элонгация. Идет при помощи ферментов ДНК-полимераз. Все полимеразы обеспечивают синтез новых цепей ДНК, новая цепь
- 52. Синтез другой цепи идет короткими фрагментами – фрагментами Оказаки. Эта цепь называется запаздывающей. Длина ФО у
- 53. ДНК-полимеразы. У прокариот известно 3 вида ДНК-полимераз: 1) ДНК-П I 2) ДНК-П II 3) ДНК-П III
- 54. ДНК-П I открыта в 1960 году А. Корнбергом и поэтому её называют ферментом Корнберга. ДНК-П II
- 55. 3. Терминация репликации. У кишечной палочки есть ter-сайты, где происходит терминация репликации. У эукариот терминация репликации
- 57. В 80-х годах 20 века было установлено, что на концах хромосом есть особые структуры – теломеры,
- 58. Во время деления теломеры теряют от 5 до 20 фрагментов и с каждым делением становятся короче,
- 59. Проблема концевой недорепликации. Репликация на отстающей цепи ДНК начинается с синтеза коротких РНК-праймеров или затравок, с
- 60. 8-12 нуклеотидов короче исходной. Таким образом, если в клетке нет механизмов, которые могли бы компенсировать потерю
- 61. Т.о., к началу 90-х годов ХХ века молекулярная структура теломеры была открыта, а проблема неполной репликации
- 62. Перед репликацией ДНК теломераза добавляет несколько копий теломерных повторов на 3` конец ДНК. Теломераза удлиняет не
- 63. По окончании репликации остается незаполненным только участок РНК-затравки, синтезированный на теломерной последовательности. В итоге дочерние цепи
- 64. Транскрипция Реализация генетической информации о структуре определенного белка включает два этапа: транскрипцию и трансляцию. Транскрипция -
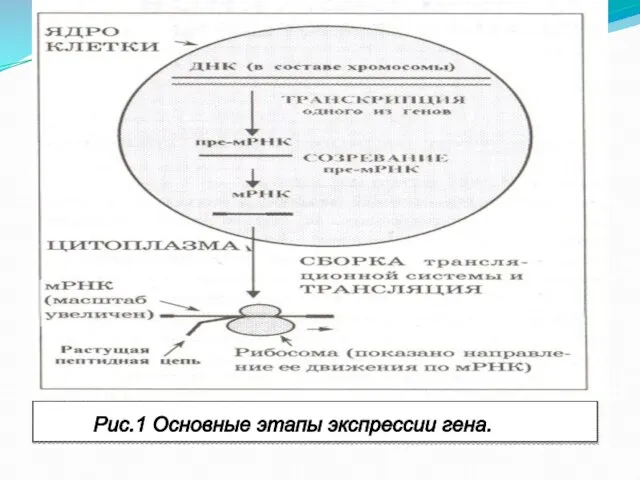
- 65. Рис.1 Основные этапы экспрессии гена.
- 66. Транскрипция ДНК происходит отдельными участками, в которые входит один или несколько генов. (см.например, оперон). Каждый ген
- 67. Транскриптон Транскрипции подвергается не вся молекула ДНК, а только ее определенный участок – транскриптон. Начало транскриптона
- 68. Транскрипция осуществляется специальным ферментом РНК–П. РНК- П узнает начало транскрибируемого участка (промотор), присоединяется к нему, расплетает
- 69. Строение и функции РНК-П РНК- П обнаружена во всех про- и эукариотических организмах. Наиболее изучен фермент
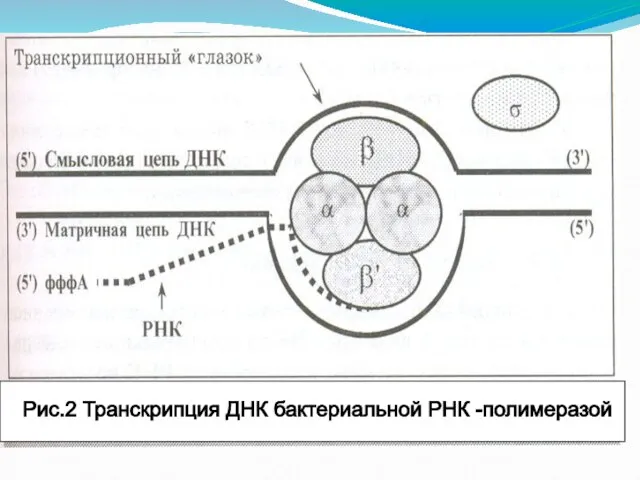
- 70. Рис.2 Транскрипция ДНК бактериальной РНК -полимеразой
- 71. Сигма – субъединица необходима для распознавания РНК-П специфического участка - промотора на ДНК, с которого начинается
- 72. Взаимодействие core-фермента с δ-субъединицей приводит к формированию холоэнзима РНК-П. Сигма –фактор после начала синтеза РНК, сразу
- 73. У прокариот функционирует одна единственная РНК-П, которая принимает участие в синтезе всех видов РНК: мРНК, тРНК
- 74. Характерные особенности фермента РНК-П: 1). способность с помощью δ-субъединицы выбирать цепь ДНК, с которой будет производиться
- 75. В клетках эукариот существуют три различные РНК-П (I,II,III), каждая из которых представлена самостоятельными полипептидами. Эукариотические РНК-П
- 76. Функции РНК-полимераз у эукариот: РНК –полимераза І сосредоточена в основном в ядрышке и транскрибирует гены предшественников
- 77. РНК –П ІІІ содержится в нуклеоплазме и транскрибирует в основном гены пре - тРНК т.е. транспортные
- 78. Этапы транскрипции Инициация-это первый этап транскрипции, где у прокариот РНК-П с участием δ-фактора узнает промотор и
- 79. Элонгация транскрипции. РНК-полимераза перемещается вдоль структурных генов оперона, соответственно перемещается и «транскрипционный глазок», синтезируется молекула РНК
- 80. Нити ДНК перед транскрипционным комплексом (ДНК – РНК полимераза- РНК ) разделяются, а позади него вновь
- 81. Терминация - завершающий этап транскрипции. Сигналом терминации служат специальные ГЦ богатые участки в конце генов. Сила
- 82. У бактерий, специальный белок RhO- фактор также обладает расплетающей активностью и облегчает расхождение цепей РНК и
- 83. Обычно на каждом транскрибируемом гене работают , двигаясь друг за другом несколько молекул РНК – полимераз
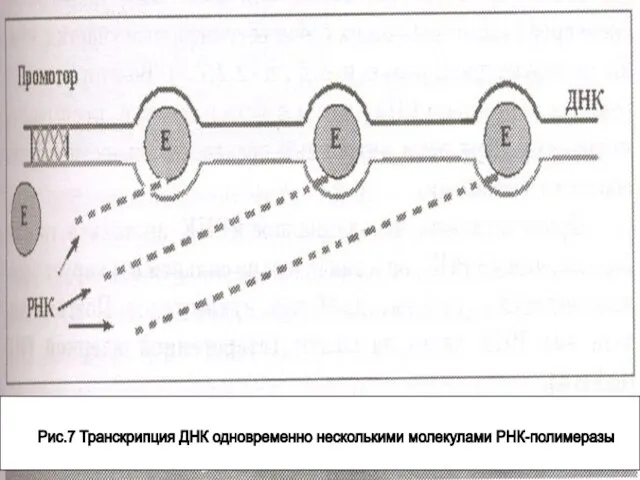
- 84. Рис.7 Транскрипция ДНК одновременно несколькими молекулами РНК-полимеразы
- 85. Особенности транскрипции у эукариот: Хромосомы эукариот имеют нуклеосомное строение, поэтому необходимы приспособления для освобождения ДНК от
- 86. . К настоящему времени выделены шесть общих транскрипционных факторов: ТFIIА, ТFIIВ, ТFIIД, ТFIIЕ, ТFIIF, ТFIIН.
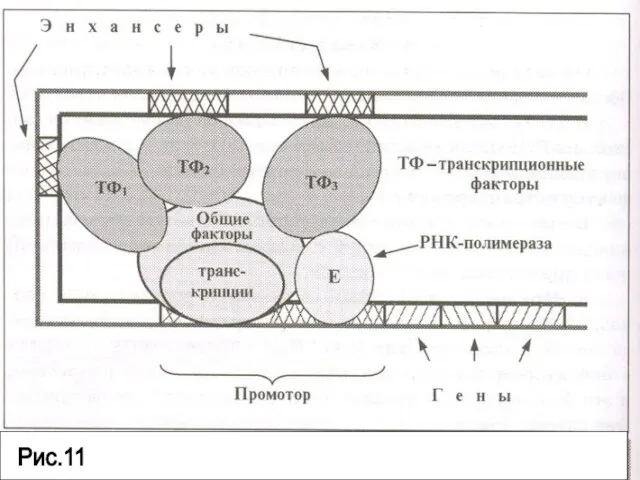
- 87. . Для активации транскрипции необходимо участие также специфических факторов транскрипции – энхансеров. Энхансеры располагаются достаточно далеко
- 88. ДНК образует петли благодаря чему энхансеры сближаются с промоторной зоной и влияют на активность транскрипционного комплекса.
- 89. Параллельно энхансерам, усиливающим транскрипцию, в геноме существуют специфические последовательности- сайленсеры. Сайленсеры также связываются с комплексом специфических
- 90. Рис.11
- 91. Продукты транскрипции. В результате транскрипции у эукариот образуются предшественники тех или иных РНК: м РНК,рРНК,и тРНК.
- 92. Длина таких пре м- РНК у разных молекул варьирует от 2 тыс. до 20 тысяч нуклеотидов.
- 93. Процессинг (созревание) мРНК Процессинг включает следующие преобразования: кэпирование полиаденилирование сплайсинг
- 94. Рис.13
- 95. Эукариотические мРНК стабильны в течение часов и суток.Это объясняется,во-первых, стабилизацией 5’- и 3’- концов, а во-вторых,
- 96. Альтернативный сплайсинг - это соединение экзонов одного гена в разных комбинациях с образованием различных зрелых мРНК.
- 97. Механизмы альтернативного сплайсинга 1.Использование разл. промоторов. При этом образуются транскрипты с разным количеством экзонов, имеющие разные
- 98. 3.Выбор различных экзонов из одинаковых пре-мРНК. При этом для формирования зрелых РНК могут использоваться различные экзоны,
- 99. Понятие о генетическом коде Систему расположения нуклеотидов в ДНК, определяющую последовательность расположения аминокислот в белке называют
- 100. Свойства генетического кода: Триплетность. Единицей информации в кодирующей цепи ДНК является триплет- последовательность из трех нуклеотидов.
- 101. 3. Однозначность. Каждый триплет кодирует только одну определенную аминокислоту. 4. Непрерывность. Код внутри гена не содержит
- 102. Коллинеарность. Последовательность триплетов в экзонах гена соответствует последовательности аминокислот в белке. Универсальность. Генетический код един для
- 103. Трансляция- это второй этап реализации генетической информации. Она заключается в синтезе полипептидов на рибосоме. В трансляции
- 104. Исходными материалом, из которого строится белок, являются аминокислоты. Однако свободные аминокислоты не используются рибосомой. Для того
- 105. Этапы трансляции 1. Инициация 2. Элонгация 3. Терминация
- 106. Инициация К малой субъединице рибосомы присоединяются т-РНК с метионином, инициирующий кодон АУГ. Антикодон т-РНК с метионином
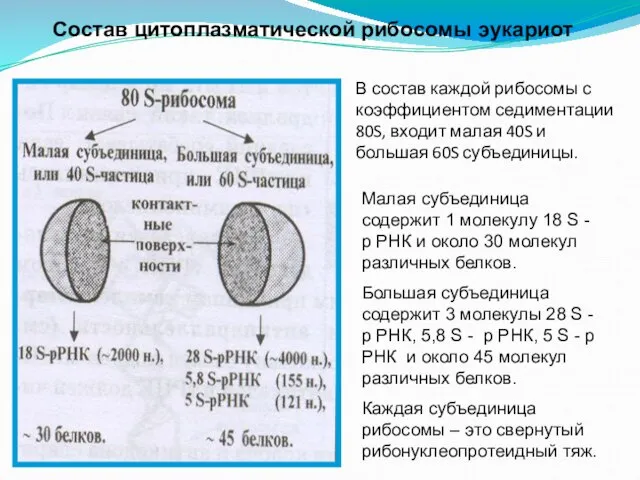
- 107. В состав каждой рибосомы с коэффициентом седиментации 80S, входит малая 40S и большая 60S субъединицы. Малая
- 108. Процесс трансляции начинается со сборки активной рибосомы – инициации трансляции. Сборка происходит строго упорядоченным образом, что
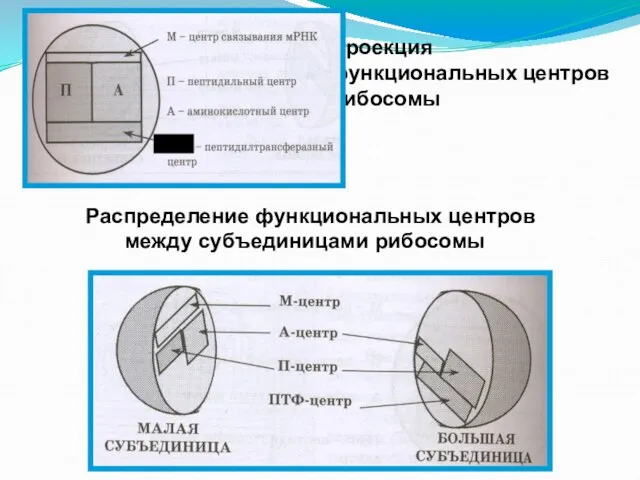
- 109. Функциональные центры рибосом Собранная рибосома напоминает форму сердца (без полостей), правые отделы образованы малой субъединицей, а
- 110. Выполнение функций рибосомы связано с наличием на ней определенных функциональных центров: а) Центр связывания м РНК
- 111. в) Аминокислотный центр (А-центр): место связывания очередной аа-т-РНК г) Пептидилтрансферазный центр (ПТФ-центр): он катализирует перенос пептидила
- 112. Проекция функциональных центров рибосомы ПТЦ Распределение функциональных центров между субъединицами рибосомы
- 113. Элонгация Включает 3 процесса: 1.В рибосому, у которой в П-центре находится Мет-тРНК, в А-центр присоединяется первая
- 114. 2.Образование пептидной связи 3.Транслокация (продвижение рибосомы). Рибосома перемещается на один кодон в направлении от 5 к
- 115. а) Связывание аа-тРНК со свободным А- участком. Путем спаривания нуклеотидов антикодона с тремя нуклеотидами мРНК в
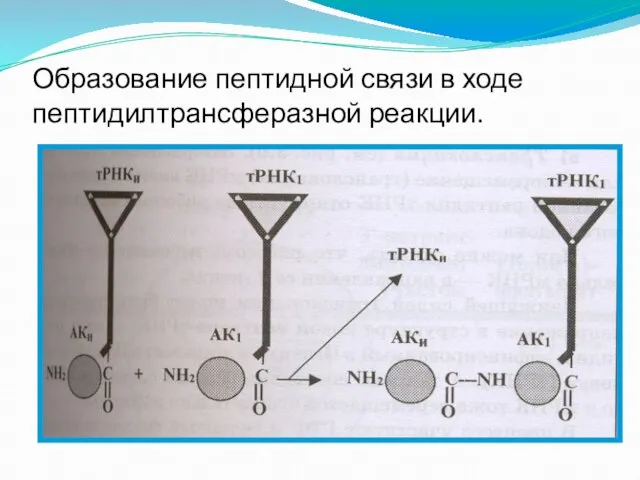
- 116. б) Карбоксильный конец полипептидной цепи отделяется в П-участке от молекулы тРНК и образует пептидную связь с
- 117. Образование пептидной связи в ходе пептидилтрансферазной реакции.
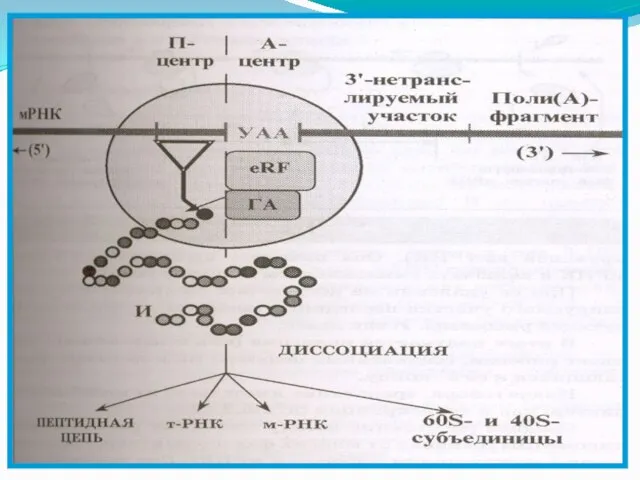
- 118. Терминация трансляции Сигналом об окончании трансляции служит появление в рибосоме одного из бессмысленных кодонов мРНК- УАА,
- 121. Особенности трансляции у прокариот: 1) Субъединицы рибосом несколько меньше по размеру чем у эукариот(30S и 50S)
- 122. 4) Сопряжение трянсляции с транскрипцией. 5) Сопряжение синтеза нескольких пептидных цепей. 6) Инициаторная аа-тРНК в виде
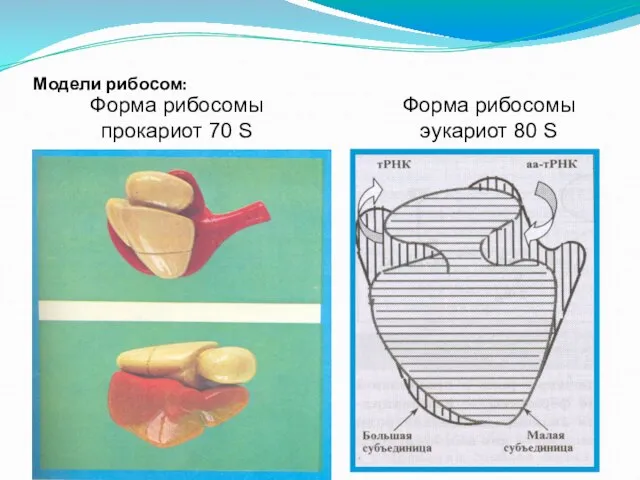
- 123. Модели рибосом: Форма рибосомы эукариот 80 S Форма рибосомы прокариот 70 S
- 124. Модели субчастицы 30S рибосомы: три различные проекции (прокариот) Субчастица 30 S содержит 1 молекулу 16 S
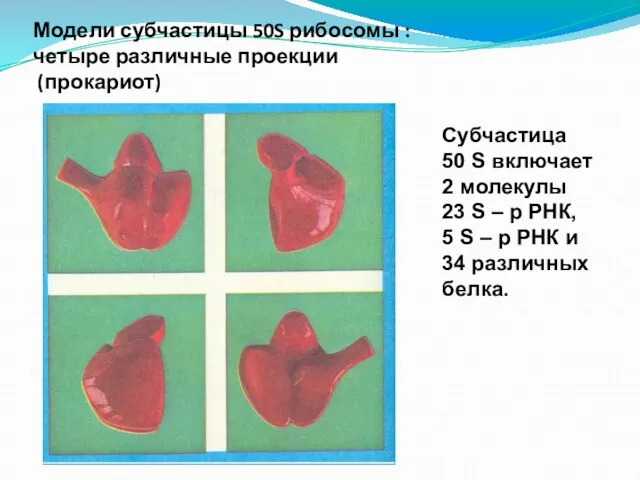
- 125. Модели субчастицы 50S рибосомы : четыре различные проекции (прокариот) Субчастица 50 S включает 2 молекулы 23
- 126. Общие представления о фолдинге белков Трансляция мРНК приводит к образованию пептидной цепи со строго определенной последовательностью
- 127. Вспомогательные факторы фолдинга: а) Фолдазы – белки с каталитической активностью: протеиндисульфидизомераза, пептидилпролилизомераза. б) Молекулярные шапероны. К
- 129. Скачать презентацию
Слайд 2
План лекции:
Нуклеиновые кислоты – классификация, строение, функции.
Макромолекулярная структура ДНК
РНК:
План лекции:
Нуклеиновые кислоты – классификация, строение, функции.
Макромолекулярная структура ДНК
РНК:

Слайд 3Нуклеиновые кислоты (НК)
Два вида НК:
ДНК (хранение наследственной информации)
РНК (реализация наследственной информации)
Нуклеиновые кислоты (НК)
Два вида НК:
ДНК (хранение наследственной информации)
РНК (реализация наследственной информации)

Слайд 4Нуклеиновые кислоты (НК)
Н.К.- биологические полимеры
Мономеры- нуклеотиды
Нуклеиновые кислоты (НК)
Н.К.- биологические полимеры
Мономеры- нуклеотиды

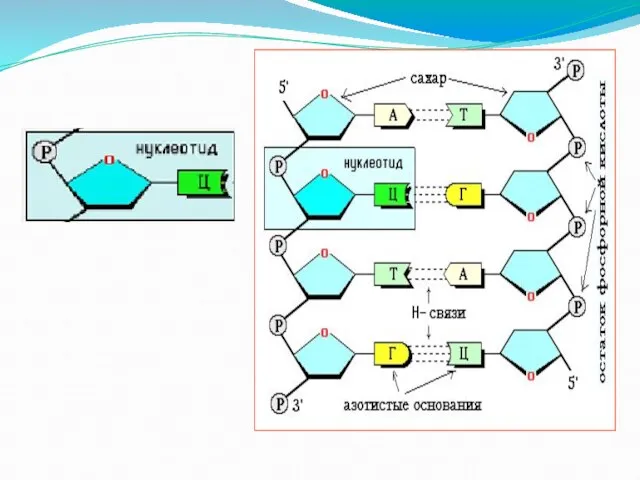
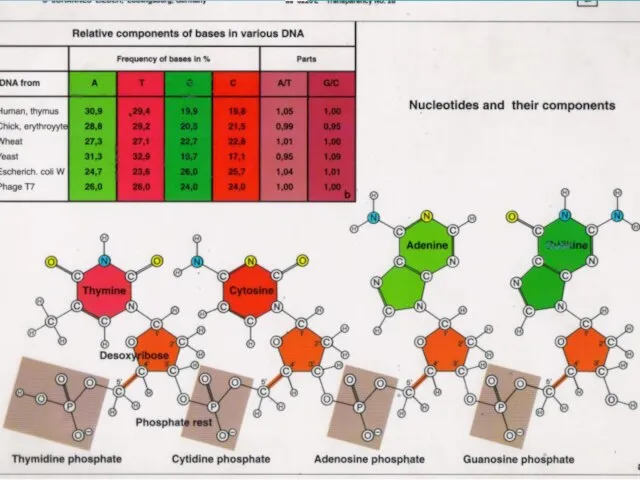
Слайд 5Состав нуклеотида
Состав нуклеотида


Слайд 7Нуклеозиды
Нуклеозиды
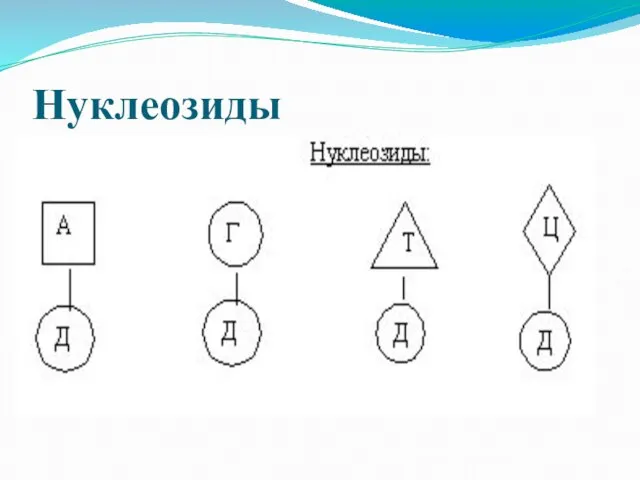
Слайд 8Нуклеотиды
Нуклеотиды

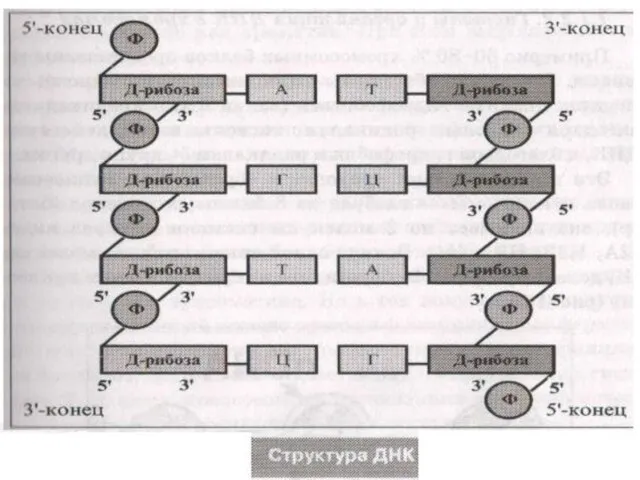
Слайд 9Модель Уотсона-Крика (1953)
1. ДНК-полимер из нуклеотидов, соединенных 3-5 фосфодиэфирными
связями
Состав нуклеотидов
Модель Уотсона-Крика (1953)
1. ДНК-полимер из нуклеотидов, соединенных 3-5 фосфодиэфирными
связями
Состав нуклеотидов

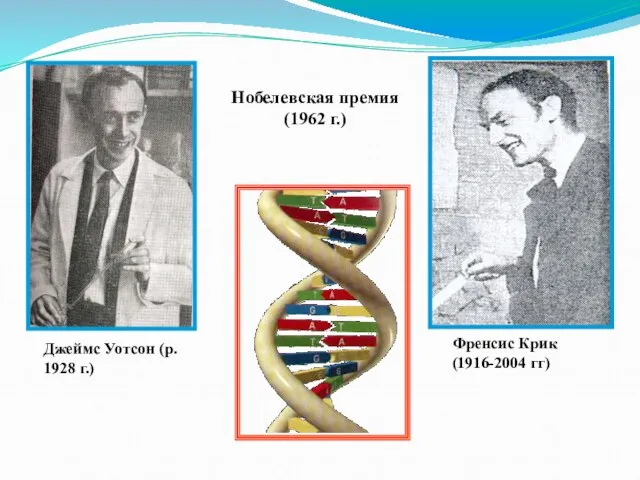
Слайд 12Френсис Крик (1916-2004 гг)
Джеймс Уотсон (р. 1928 г.)
Нобелевская премия (1962 г.)
Френсис Крик (1916-2004 гг)
Джеймс Уотсон (р. 1928 г.)
Нобелевская премия (1962 г.)
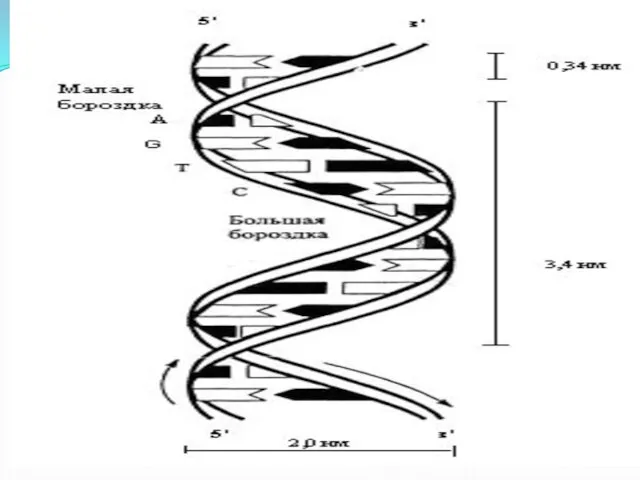
Слайд 14ДНК
При горизонтальном изображении:
5---АТТГАЦАГГЦ---3
3---ТААЦТГТЦЦГ---5
В ядре человеческой клетки – 46 молекул ДНК
Общая длина их
ДНК
При горизонтальном изображении:
5---АТТГАЦАГГЦ---3
3---ТААЦТГТЦЦГ---5
В ядре человеческой клетки – 46 молекул ДНК
Общая длина их

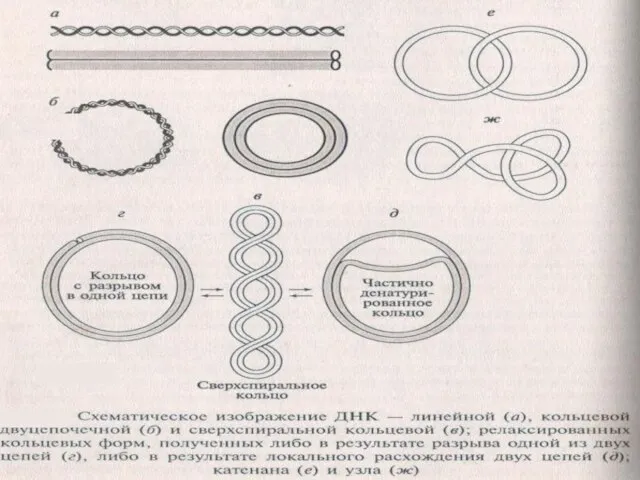
Слайд 15Разнообразие форм ДНК
Не все ДНК являются двуцепочечными. Геномы некоторых мелких вирусов бактерий,
Разнообразие форм ДНК
Не все ДНК являются двуцепочечными. Геномы некоторых мелких вирусов бактерий,

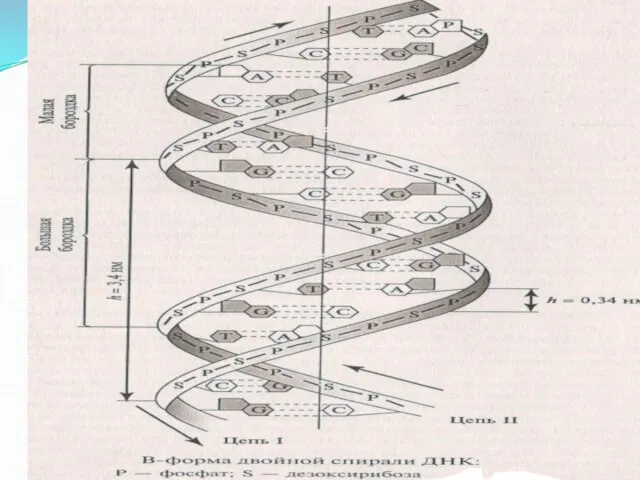
Слайд 16Формы ДНК
В-форма (правозакрученная спираль)- основная форма существования ДНК
Z –форма (левозакрученная спираль), содержит
Формы ДНК
В-форма (правозакрученная спираль)- основная форма существования ДНК
Z –форма (левозакрученная спираль), содержит

Слайд 19РНК: структура и функции
Р-РНК(80-85%)
Т-РНК (около10%)
М(И)-РНК (5%)
Мя-РНК (2%)
РНК: структура и функции
Р-РНК(80-85%)
Т-РНК (около10%)
М(И)-РНК (5%)
Мя-РНК (2%)

Слайд 20Р-РНК
Р-РНК-структурная основа рибосом
Р-РНК взаимодействуют с м-РНК и аминоацил-тРНК в процессе трансляции
Это стабильные,
Р-РНК
Р-РНК-структурная основа рибосом
Р-РНК взаимодействуют с м-РНК и аминоацил-тРНК в процессе трансляции
Это стабильные,

Слайд 21М-РНК
М-РНК несет информацию о синтезе белка на ней самой, а также информацию
М-РНК
М-РНК несет информацию о синтезе белка на ней самой, а также информацию

Слайд 22Строение М-РНК
1.Информативная зона - транслируемая область, работает как матрица в процессе биосинтеза
Строение М-РНК
1.Информативная зона - транслируемая область, работает как матрица в процессе биосинтеза

Слайд 23Строение М-РНК
КЭП 5’-НТО AUG ТРАНСЛИРУЕМАЯ ОБЛАСТЬ СТОП 3’-НТО ПОЛИ (А)
КЭП (от англ.сар-
Строение М-РНК
КЭП 5’-НТО AUG ТРАНСЛИРУЕМАЯ ОБЛАСТЬ СТОП 3’-НТО ПОЛИ (А)
КЭП (от англ.сар-

Слайд 24Т-РНК
Т-РНК переносят аминокислоты в белоксинтезирующий аппарат клетки и выступают в роли затравки
Т-РНК
Т-РНК переносят аминокислоты в белоксинтезирующий аппарат клетки и выступают в роли затравки

Слайд 25Т-РНК, особенности
1.По сравнению с р-РНК и м-РНК имеют меньшие размеры
2.Соотношение А:У и
Т-РНК, особенности
1.По сравнению с р-РНК и м-РНК имеют меньшие размеры
2.Соотношение А:У и

Слайд 26Т-РНК
Содержит в своей структуре:
1.Сайт прикрепления аминокислоты (акцепторный конец)
2.Сайт связывания с рибосомой
3.Антикодон
Т-РНК
Содержит в своей структуре:
1.Сайт прикрепления аминокислоты (акцепторный конец)
2.Сайт связывания с рибосомой
3.Антикодон

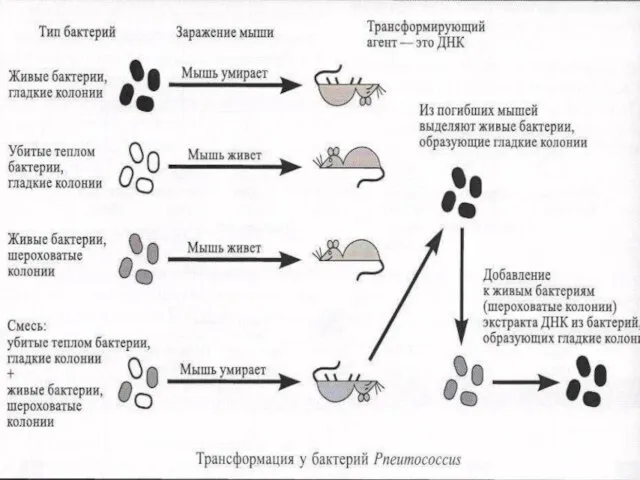
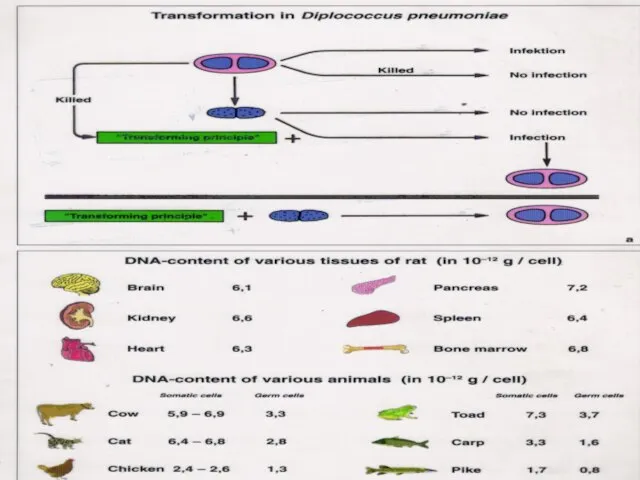
Слайд 34Постулат Крика
Центральная догма молекулярной биологии: передача наследственной информации происходит только в одном
Постулат Крика
Центральная догма молекулярной биологии: передача наследственной информации происходит только в одном

Слайд 35Типы переноса наследственной информации
Б) РНК---ДНК (обратная транскрипция). Происходит в клетках животных, зараженных
Типы переноса наследственной информации
Б) РНК---ДНК (обратная транскрипция). Происходит в клетках животных, зараженных

Слайд 36Типы переноса наследственной информации
3. Запрещенный перенос (это процессы не зарегистрированные нигде и
Типы переноса наследственной информации
3. Запрещенный перенос (это процессы не зарегистрированные нигде и

Слайд 37Репликация ДНК – процесс самоудвоения, самовоспроизведения, самокопирования наследственной информации.
Суть репликации ДНК -
Репликация ДНК – процесс самоудвоения, самовоспроизведения, самокопирования наследственной информации.
Суть репликации ДНК -

Слайд 38Репликация ДНК идет на основе следующих принципов:
Полуконсервативность
Синтез ДНК начинается с расхождения цепей,
Репликация ДНК идет на основе следующих принципов:
Полуконсервативность
Синтез ДНК начинается с расхождения цепей,

Слайд 39Консервативный способ репликации – когда после удвоения одна молекула состоит из двух
Консервативный способ репликации – когда после удвоения одна молекула состоит из двух

Слайд 40Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В состав
Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В состав

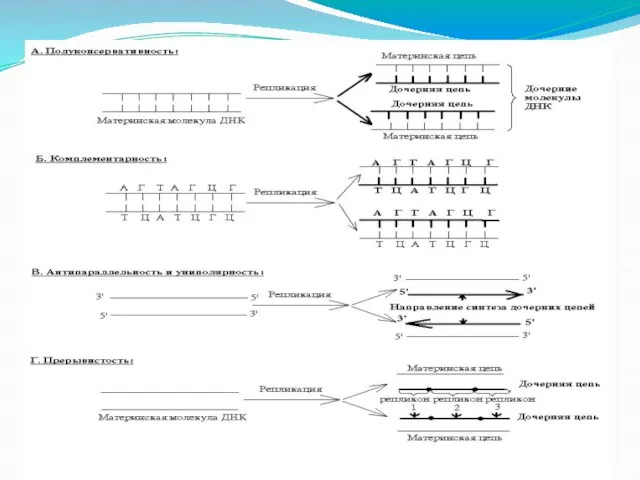
Слайд 42Участок ДНК в пределах которого репликация начинается и заканчивается называется репликоном.
В
Участок ДНК в пределах которого репликация начинается и заканчивается называется репликоном.
В

Слайд 43Молекула ДНК, вступающая в репликацию:
Точка, в которой происходит репликация называется репликационной вилкой
Молекула ДНК, вступающая в репликацию:
Точка, в которой происходит репликация называется репликационной вилкой

Слайд 44Репликация может идти либо в одном направлении, либо в двух направлениях.
Это
Репликация может идти либо в одном направлении, либо в двух направлениях.
Это

Слайд 45Область, которая уже реплицирована, имеет вид «глазка» внутри нереплицировавшейся ДНК. Этот глазок
Область, которая уже реплицирована, имеет вид «глазка» внутри нереплицировавшейся ДНК. Этот глазок
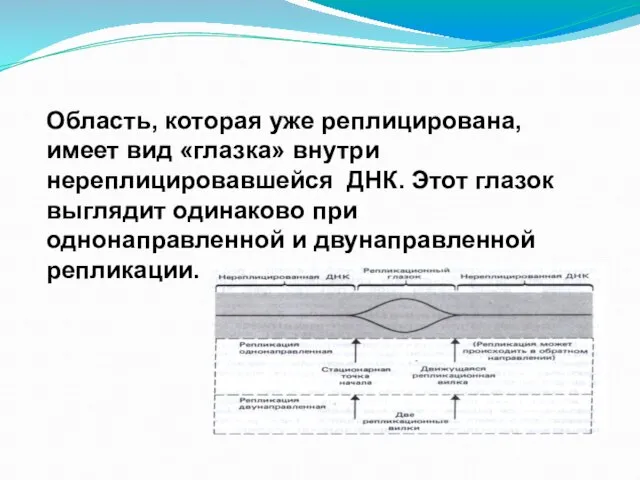
Слайд 46При однонаправленной репликации глазок имеет фиксированную точку начала и движущуюся репликационную вилку.
При однонаправленной репликации глазок имеет фиксированную точку начала и движущуюся репликационную вилку.

Слайд 47Этапы репликации:
1.Инициация идет с участием белков и ферментов, которые должны обеспечить:
1)Раскручивание ДНК
2)Связь
Этапы репликации:
1.Инициация идет с участием белков и ферментов, которые должны обеспечить:
1)Раскручивание ДНК
2)Связь

Слайд 48Точки начала репликации богаты парами А-Т.
Геликаза (от helix - спираль) расплетает
Точки начала репликации богаты парами А-Т.
Геликаза (от helix - спираль) расплетает

Слайд 49Инициация репликации и клеточный цикл у дрожжей.
Инициация репликации и клеточный цикл у дрожжей.
Слайд 50Инициация репликации у эукариот.
Белки инициации и происходящие процессы сходны с прокариотами.
Отличия:
1.Участие
Инициация репликации у эукариот.
Белки инициации и происходящие процессы сходны с прокариотами.
Отличия:
1.Участие

Слайд 512. Элонгация.
Идет при помощи ферментов ДНК-полимераз. Все полимеразы обеспечивают синтез новых цепей
2. Элонгация.
Идет при помощи ферментов ДНК-полимераз. Все полимеразы обеспечивают синтез новых цепей

Слайд 52Синтез другой цепи идет короткими фрагментами – фрагментами Оказаки. Эта цепь называется
Синтез другой цепи идет короткими фрагментами – фрагментами Оказаки. Эта цепь называется

Слайд 53ДНК-полимеразы. У прокариот известно 3 вида ДНК-полимераз:
1) ДНК-П I
2) ДНК-П II
3) ДНК-П
ДНК-полимеразы. У прокариот известно 3 вида ДНК-полимераз:
1) ДНК-П I
2) ДНК-П II
3) ДНК-П

Слайд 54ДНК-П I открыта в 1960 году А. Корнбергом и поэтому её называют
ДНК-П I открыта в 1960 году А. Корнбергом и поэтому её называют

Слайд 553. Терминация репликации.
У кишечной палочки есть ter-сайты, где происходит терминация репликации.
У эукариот
3. Терминация репликации.
У кишечной палочки есть ter-сайты, где происходит терминация репликации.
У эукариот


Слайд 57В 80-х годах 20 века было установлено, что на концах хромосом есть
В 80-х годах 20 века было установлено, что на концах хромосом есть

Слайд 58Во время деления теломеры теряют от 5 до 20 фрагментов и с
Во время деления теломеры теряют от 5 до 20 фрагментов и с

Слайд 59Проблема концевой недорепликации.
Репликация на отстающей цепи ДНК начинается с синтеза коротких
Проблема концевой недорепликации.
Репликация на отстающей цепи ДНК начинается с синтеза коротких

Слайд 608-12 нуклеотидов короче исходной. Таким образом, если в клетке нет механизмов, которые
8-12 нуклеотидов короче исходной. Таким образом, если в клетке нет механизмов, которые

Слайд 61Т.о., к началу 90-х годов ХХ века молекулярная структура теломеры была открыта,
Т.о., к началу 90-х годов ХХ века молекулярная структура теломеры была открыта,

Слайд 62Перед репликацией ДНК теломераза добавляет несколько копий теломерных повторов на 3` конец
Перед репликацией ДНК теломераза добавляет несколько копий теломерных повторов на 3` конец

Слайд 63По окончании репликации остается незаполненным только участок РНК-затравки, синтезированный на теломерной последовательности.
По окончании репликации остается незаполненным только участок РНК-затравки, синтезированный на теломерной последовательности.

Слайд 64Транскрипция
Реализация генетической информации о структуре определенного белка включает два этапа: транскрипцию и
Транскрипция
Реализация генетической информации о структуре определенного белка включает два этапа: транскрипцию и

Слайд 65Рис.1 Основные этапы экспрессии гена.
Рис.1 Основные этапы экспрессии гена.
Слайд 66Транскрипция ДНК происходит отдельными участками, в которые входит один или несколько генов.
Транскрипция ДНК происходит отдельными участками, в которые входит один или несколько генов.

Слайд 67Транскриптон
Транскрипции подвергается не вся молекула ДНК, а только ее определенный участок –
Транскриптон
Транскрипции подвергается не вся молекула ДНК, а только ее определенный участок –

Слайд 68Транскрипция осуществляется специальным ферментом РНК–П.
РНК- П узнает начало транскрибируемого участка (промотор),
Транскрипция осуществляется специальным ферментом РНК–П.
РНК- П узнает начало транскрибируемого участка (промотор),

Слайд 69Строение и функции РНК-П
РНК- П обнаружена во всех про- и
Строение и функции РНК-П
РНК- П обнаружена во всех про- и

Слайд 70Рис.2 Транскрипция ДНК бактериальной РНК -полимеразой
Рис.2 Транскрипция ДНК бактериальной РНК -полимеразой
Слайд 71Сигма – субъединица необходима для распознавания РНК-П специфического участка - промотора на
Сигма – субъединица необходима для распознавания РНК-П специфического участка - промотора на

Слайд 72Взаимодействие core-фермента с δ-субъединицей приводит к формированию холоэнзима РНК-П.
Сигма –фактор после
Взаимодействие core-фермента с δ-субъединицей приводит к формированию холоэнзима РНК-П.
Сигма –фактор после

Слайд 73У прокариот функционирует одна единственная РНК-П, которая принимает участие в синтезе всех
У прокариот функционирует одна единственная РНК-П, которая принимает участие в синтезе всех

Слайд 74Характерные особенности фермента РНК-П:
1). способность с помощью δ-субъединицы выбирать цепь ДНК,
Характерные особенности фермента РНК-П:
1). способность с помощью δ-субъединицы выбирать цепь ДНК,

Слайд 75В клетках эукариот существуют три различные РНК-П (I,II,III), каждая из которых представлена
В клетках эукариот существуют три различные РНК-П (I,II,III), каждая из которых представлена

Слайд 76Функции РНК-полимераз у эукариот:
РНК –полимераза І сосредоточена в основном в ядрышке
Функции РНК-полимераз у эукариот:
РНК –полимераза І сосредоточена в основном в ядрышке

Слайд 77РНК –П ІІІ содержится в нуклеоплазме и транскрибирует в основном гены пре
РНК –П ІІІ содержится в нуклеоплазме и транскрибирует в основном гены пре

Слайд 78Этапы транскрипции
Инициация-это первый этап транскрипции, где у прокариот РНК-П с участием δ-фактора
Этапы транскрипции
Инициация-это первый этап транскрипции, где у прокариот РНК-П с участием δ-фактора

Слайд 79 Элонгация транскрипции.
РНК-полимераза перемещается вдоль структурных генов оперона, соответственно перемещается
Элонгация транскрипции.
РНК-полимераза перемещается вдоль структурных генов оперона, соответственно перемещается

Слайд 80Нити ДНК перед транскрипционным комплексом (ДНК – РНК полимераза- РНК ) разделяются,
Нити ДНК перед транскрипционным комплексом (ДНК – РНК полимераза- РНК ) разделяются,

Слайд 81Терминация - завершающий этап транскрипции. Сигналом терминации служат специальные ГЦ богатые участки
Терминация - завершающий этап транскрипции. Сигналом терминации служат специальные ГЦ богатые участки

Слайд 82У бактерий, специальный белок RhO- фактор также обладает расплетающей активностью и облегчает
У бактерий, специальный белок RhO- фактор также обладает расплетающей активностью и облегчает

Слайд 83Обычно на каждом транскрибируемом гене работают , двигаясь друг за другом несколько
Обычно на каждом транскрибируемом гене работают , двигаясь друг за другом несколько

Слайд 84Рис.7 Транскрипция ДНК одновременно несколькими молекулами РНК-полимеразы
Рис.7 Транскрипция ДНК одновременно несколькими молекулами РНК-полимеразы
Слайд 85Особенности транскрипции у эукариот:
Хромосомы эукариот имеют нуклеосомное строение, поэтому необходимы приспособления для
Особенности транскрипции у эукариот:
Хромосомы эукариот имеют нуклеосомное строение, поэтому необходимы приспособления для

Слайд 86.
К настоящему времени выделены шесть общих транскрипционных факторов: ТFIIА, ТFIIВ, ТFIIД,
.
К настоящему времени выделены шесть общих транскрипционных факторов: ТFIIА, ТFIIВ, ТFIIД,

Слайд 87.
Для активации транскрипции необходимо участие также специфических факторов транскрипции – энхансеров.
Энхансеры
.
Для активации транскрипции необходимо участие также специфических факторов транскрипции – энхансеров.
Энхансеры

Слайд 88ДНК образует петли благодаря чему энхансеры сближаются с промоторной зоной и влияют
ДНК образует петли благодаря чему энхансеры сближаются с промоторной зоной и влияют

Слайд 89Параллельно энхансерам, усиливающим транскрипцию, в геноме существуют специфические последовательности- сайленсеры.
Сайленсеры также
Параллельно энхансерам, усиливающим транскрипцию, в геноме существуют специфические последовательности- сайленсеры.
Сайленсеры также

Слайд 90Рис.11
Рис.11
Слайд 91Продукты транскрипции.
В результате транскрипции у эукариот образуются предшественники тех или иных РНК:
Продукты транскрипции.
В результате транскрипции у эукариот образуются предшественники тех или иных РНК:

Слайд 92Длина таких пре м- РНК у разных молекул варьирует от 2 тыс.
Длина таких пре м- РНК у разных молекул варьирует от 2 тыс.

Слайд 93
Процессинг (созревание) мРНК
Процессинг включает следующие преобразования:
кэпирование
полиаденилирование
сплайсинг
Процессинг (созревание) мРНК
Процессинг включает следующие преобразования:
кэпирование
полиаденилирование
сплайсинг

Слайд 94Рис.13
Рис.13
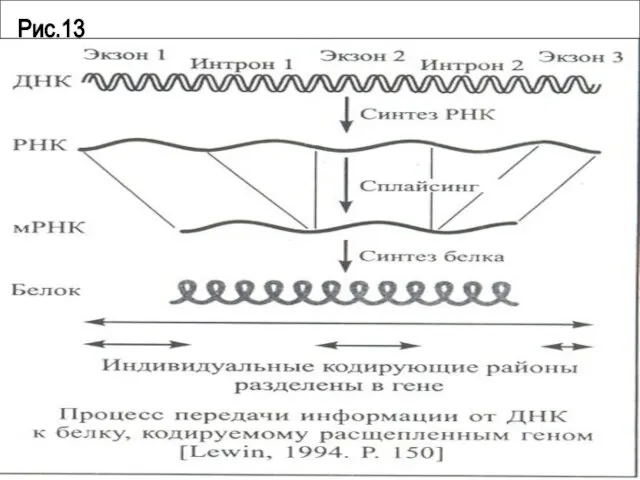
Слайд 95Эукариотические мРНК стабильны в течение часов и суток.Это объясняется,во-первых, стабилизацией 5’- и
Эукариотические мРНК стабильны в течение часов и суток.Это объясняется,во-первых, стабилизацией 5’- и

Слайд 96Альтернативный сплайсинг -
это соединение экзонов одного гена в разных комбинациях с
Альтернативный сплайсинг -
это соединение экзонов одного гена в разных комбинациях с

Слайд 97Механизмы альтернативного сплайсинга
1.Использование разл. промоторов.
При этом образуются транскрипты с разным количеством
Механизмы альтернативного сплайсинга
1.Использование разл. промоторов.
При этом образуются транскрипты с разным количеством

Слайд 983.Выбор различных экзонов из одинаковых пре-мРНК. При этом для формирования зрелых РНК
3.Выбор различных экзонов из одинаковых пре-мРНК. При этом для формирования зрелых РНК

Слайд 99Понятие о генетическом коде
Систему расположения нуклеотидов в ДНК, определяющую последовательность расположения аминокислот
Понятие о генетическом коде
Систему расположения нуклеотидов в ДНК, определяющую последовательность расположения аминокислот

Слайд 100Свойства генетического кода:
Триплетность. Единицей информации в кодирующей цепи ДНК является триплет- последовательность
Триплетность. Единицей информации в кодирующей цепи ДНК является триплет- последовательность

Слайд 1013. Однозначность. Каждый триплет кодирует только одну определенную аминокислоту.
4. Непрерывность. Код внутри
3. Однозначность. Каждый триплет кодирует только одну определенную аминокислоту.
4. Непрерывность. Код внутри

Слайд 102Коллинеарность. Последовательность триплетов в экзонах гена соответствует последовательности аминокислот в белке.
Универсальность. Генетический
Коллинеарность. Последовательность триплетов в экзонах гена соответствует последовательности аминокислот в белке.
Универсальность. Генетический

Слайд 103 Трансляция- это второй этап реализации генетической информации. Она заключается в синтезе
Трансляция- это второй этап реализации генетической информации. Она заключается в синтезе

Слайд 104Исходными материалом, из которого строится белок, являются аминокислоты. Однако свободные аминокислоты не
Исходными материалом, из которого строится белок, являются аминокислоты. Однако свободные аминокислоты не

Слайд 105Этапы трансляции
1. Инициация
2. Элонгация
3. Терминация
Этапы трансляции
1. Инициация
2. Элонгация
3. Терминация

Слайд 106Инициация
К малой субъединице рибосомы присоединяются т-РНК с метионином, инициирующий кодон АУГ. Антикодон
Инициация
К малой субъединице рибосомы присоединяются т-РНК с метионином, инициирующий кодон АУГ. Антикодон

Слайд 107В состав каждой рибосомы с коэффициентом седиментации 80S, входит малая 40S и
В состав каждой рибосомы с коэффициентом седиментации 80S, входит малая 40S и
Слайд 108Процесс трансляции начинается со сборки активной рибосомы – инициации трансляции.
Сборка происходит
Процесс трансляции начинается со сборки активной рибосомы – инициации трансляции. Сборка происходит

Слайд 109Функциональные центры рибосом
Собранная рибосома напоминает форму сердца (без полостей), правые отделы образованы
Функциональные центры рибосом Собранная рибосома напоминает форму сердца (без полостей), правые отделы образованы

Слайд 110Выполнение функций рибосомы связано
с наличием на ней определенных функциональных центров:
а) Центр
Выполнение функций рибосомы связано с наличием на ней определенных функциональных центров: а) Центр

Слайд 111
в) Аминокислотный центр (А-центр):
место связывания очередной аа-т-РНК
г) Пептидилтрансферазный центр (ПТФ-центр):
он катализирует
в) Аминокислотный центр (А-центр): место связывания очередной аа-т-РНК г) Пептидилтрансферазный центр (ПТФ-центр): он катализирует

Слайд 112Проекция функциональных центров рибосомы
ПТЦ
Распределение функциональных центров между субъединицами рибосомы
Проекция функциональных центров рибосомы
ПТЦ
Распределение функциональных центров между субъединицами рибосомы
Слайд 113Элонгация
Включает 3 процесса:
1.В рибосому, у которой в П-центре находится Мет-тРНК, в А-центр
Элонгация
Включает 3 процесса:
1.В рибосому, у которой в П-центре находится Мет-тРНК, в А-центр

Слайд 1142.Образование пептидной связи
3.Транслокация (продвижение рибосомы). Рибосома перемещается на один кодон в направлении
2.Образование пептидной связи
3.Транслокация (продвижение рибосомы). Рибосома перемещается на один кодон в направлении

Слайд 115а) Связывание аа-тРНК со свободным А- участком. Путем спаривания нуклеотидов антикодона с
а) Связывание аа-тРНК со свободным А- участком. Путем спаривания нуклеотидов антикодона с
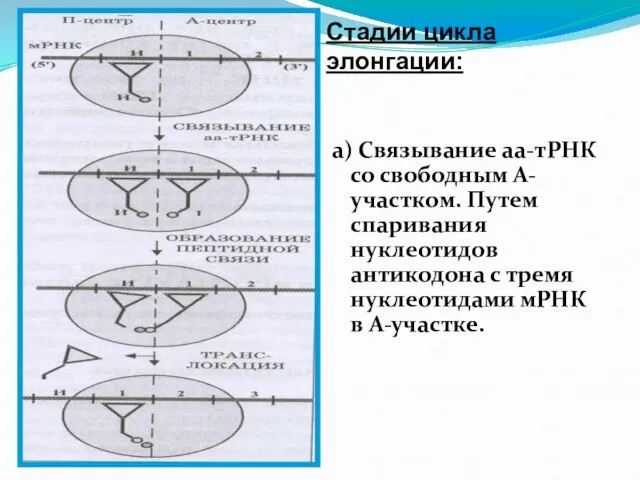
Слайд 116б) Карбоксильный конец полипептидной цепи отделяется в П-участке от молекулы тРНК и
б) Карбоксильный конец полипептидной цепи отделяется в П-участке от молекулы тРНК и

Слайд 117Образование пептидной связи в ходе пептидилтрансферазной реакции.
Образование пептидной связи в ходе пептидилтрансферазной реакции.
Слайд 118Терминация трансляции
Сигналом об окончании трансляции служит появление в рибосоме одного из бессмысленных
Терминация трансляции
Сигналом об окончании трансляции служит появление в рибосоме одного из бессмысленных


Слайд 121Особенности трансляции у прокариот:
1) Субъединицы рибосом несколько меньше по размеру чем у
Особенности трансляции у прокариот:
1) Субъединицы рибосом несколько меньше по размеру чем у

Слайд 1224) Сопряжение трянсляции с транскрипцией.
5) Сопряжение синтеза нескольких пептидных цепей.
6) Инициаторная аа-тРНК
4) Сопряжение трянсляции с транскрипцией.
5) Сопряжение синтеза нескольких пептидных цепей.
6) Инициаторная аа-тРНК

Слайд 123Модели рибосом:
Форма рибосомы эукариот 80 S
Форма рибосомы прокариот 70 S
Модели рибосом:
Форма рибосомы эукариот 80 S
Форма рибосомы прокариот 70 S
Слайд 124Модели субчастицы 30S рибосомы:
три различные проекции
(прокариот)
Субчастица 30 S содержит 1 молекулу
Модели субчастицы 30S рибосомы:
три различные проекции
(прокариот)
Субчастица 30 S содержит 1 молекулу

Слайд 125Модели субчастицы 50S рибосомы :
четыре различные проекции
(прокариот)
Субчастица 50 S включает 2
Модели субчастицы 50S рибосомы :
четыре различные проекции
(прокариот)
Субчастица 50 S включает 2
Слайд 126Общие представления о фолдинге белков
Трансляция мРНК приводит к образованию пептидной цепи со
Общие представления о фолдинге белков
Трансляция мРНК приводит к образованию пептидной цепи со

Слайд 127Вспомогательные факторы фолдинга:
а) Фолдазы – белки с каталитической активностью: протеиндисульфидизомераза,
пептидилпролилизомераза.
б) Молекулярные
Вспомогательные факторы фолдинга:
а) Фолдазы – белки с каталитической активностью: протеиндисульфидизомераза,
пептидилпролилизомераза.
б) Молекулярные

 Модульная структурасистемы ITAS
Модульная структурасистемы ITAS Инвестор, новый уровень
Инвестор, новый уровень Разработка и изготовление ансамбля коллекции женской одежды Butterfly
Разработка и изготовление ансамбля коллекции женской одежды Butterfly Илья Муромец и Соловей-разбойник
Илья Муромец и Соловей-разбойник Времена года. Лабораторная работа №5
Времена года. Лабораторная работа №5 «СОРОК МГНОВЕНИЙ НАЕДИНЕ С КОРНЕМ СТЕПЕНИ n»
«СОРОК МГНОВЕНИЙ НАЕДИНЕ С КОРНЕМ СТЕПЕНИ n» «Регион 74 в составе Российской Федерации».
«Регион 74 в составе Российской Федерации». Правовая информация для несовершеннолетних и их родителей
Правовая информация для несовершеннолетних и их родителей Дальневосточная пожарно-спасательная академия
Дальневосточная пожарно-спасательная академия Воспитательный потенциал современного образования:вызов родительской общественности
Воспитательный потенциал современного образования:вызов родительской общественности СТРЕССОВЫЕ СИТУАЦИИ НА РАБОЧЕМ МЕСТЕ
СТРЕССОВЫЕ СИТУАЦИИ НА РАБОЧЕМ МЕСТЕ Винсент Ван Гог. Подсолнухи
Винсент Ван Гог. Подсолнухи Аккумуляторная мотопила stihl gta 26
Аккумуляторная мотопила stihl gta 26 Китай
Китай Об использовании новых организационных форм медицинского обеспечения и оценки результатов новой системы оплаты труда
Об использовании новых организационных форм медицинского обеспечения и оценки результатов новой системы оплаты труда Татарское декоративно-прикладное искусство
Татарское декоративно-прикладное искусство Буквы Ч,ч, обозначающие звук [ч’]
Буквы Ч,ч, обозначающие звук [ч’] Разрезы в аксонометрических проекциях
Разрезы в аксонометрических проекциях Чему учил китайский мудрец Конфуций
Чему учил китайский мудрец Конфуций Порядок обращения за страховой пенсией по случаю потери кормильца
Порядок обращения за страховой пенсией по случаю потери кормильца Презентация на тему Шолохов «Донские рассказы»
Презентация на тему Шолохов «Донские рассказы» 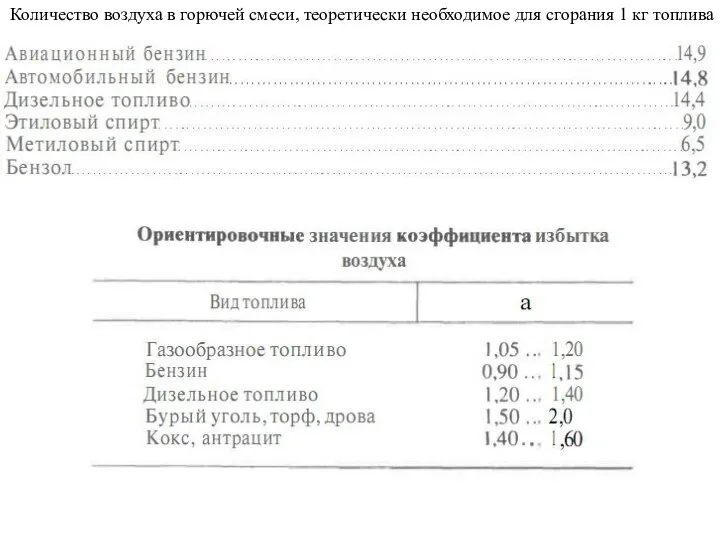 Сгорание топлива. Октановое число
Сгорание топлива. Октановое число Налоговый потенциал
Налоговый потенциал Влияние плавания на здоровье человека
Влияние плавания на здоровье человека Топочные устройства ПК
Топочные устройства ПК Иллюстрационный материал к реферату на тему: Эмпирическая школа управления
Иллюстрационный материал к реферату на тему: Эмпирическая школа управления Can you swim?
Can you swim? МОДЕЛИ УПРАВЛЕНИЯ ЗАПАСАМИ МОДЕЛИ УПРАВЛЕНИЯ ЗАПАСАМИ
МОДЕЛИ УПРАВЛЕНИЯ ЗАПАСАМИ МОДЕЛИ УПРАВЛЕНИЯ ЗАПАСАМИ