- Общая физиология возбудимых клеток.Синапс

Содержание
- 2. Возбудимые клетки: их виды, свойства. Понятие о раздражимости, возбудимости и возбуждении, порогах раздражения, зависимости ответной реакции
- 3. Нейрон (возбудимость, проводимость) Мышечная клетка (возбудимость, проводимость, сократимость)
- 6. Раздражимость – это универсальное свойство клеток отвечать на действие раздражителя изменением процессов их жизнедеятельности. Возбудимость –
- 7. Порог силы раздражителя - это минимальная величина силы действующего раздражителя, достаточная для вызова возбуждения. Раздражители, сила
- 8. Порог времени раздражения - минимальное время, в течении которого должен действовать раздражитель пороговой силы, чтобы вызвать
- 9. Минимальный градиент (крутизна) нарастания силы раздражителя во времени. Это минимальная скорость увеличения силы раздражителя во времени,
- 10. Хронаксия - минимальное время, в течение которого должен действовать раздражитель, равный по силе двум реобазам, чтобы
- 11. Адаптация – это приспособление к изменению условий внешней или внутренней среды, направленное на поддержание гомеостаза, обеспечивающее
- 12. Лабильность - функциональная подвижность возбудимой ткани, зависящая от скорости элементарных физико-химических превращений, лежащих в основе одиночного
- 13. 2 Законы реагирования возбудимых клеток на действие раздражителей.
- 14. Характер реагирования возбудимых тканей на действие раздражителей в классической физиологии принято описывать 5 законами раздражения. 1)
- 15. Закон силы раздражения - утверждает, что при увеличении сверхпорогового раздражителя до определенного предела возрастает и величина
- 16. Закон длительности раздражения – утверждает, что чем больше продолжительность сверхпорогового раздражения, тем больше величина ответной реакции,
- 17. Закон градиента раздражения – утверждает, что чем больше градиент нарастания силы раздражителя во времени, тем больше
- 18. Закон “все или ничего” – утверждает, что при действии подпороговых раздражителей возбуждение не возникает, а при
- 19. Закон полярного действия тока (закон Пфлюгера) – при действии на возбудимые клетки постоянного электрического тока в
- 20. 3 Современные представления о природе возбуждения. Биоэлектрогенез: потенциал покоя, локальный ответ, потенциал действия. Мембранно-ионная теория их
- 21. К возбудимым клеткам относят нервные и мышечные клетки, способные генерировать ПД. Биопотенциалы (биоэлектрические потенциалы) возникают вследствие
- 22. Роль биопотенциалов возбудимых тканей Участие в механизмах восприятия воздействия различных факторов среды существования 2. Обеспечение быстрой
- 23. Потенциал покоя – это разность потенциалов между внутренней и наружной поверхностью мембраны клетки, которая находится в
- 26. Потенциал действия – это кратко-временное высокоамплитудное колебание разности потенциалов мембраны возбудимой клетки, сопровождающееся изменением знака её
- 27. Признак ПД - появление кратковременной инверсии (перемены) знака заряда на мембране. Снаружи он на короткое время
- 28. ПД подразделяется на 2 большие фазы: - Деполяризации - Реполяризации
- 29. Деполяризация – уменьшение электроотрицательности внутренней стороны мембраны, на графике вся восходящая часть, выделяют медленную деполяризацию от
- 30. Реполяризация – нисходящая часть ПД, которая отражает процесс восстановления исходной поляризации мембраны (исходного потенциала). Выделяют: А)
- 31. Гиперполяризация – увеличение электроотрицательности внутренней стороны мембраны, от Е0 в сторону увеличения заряда внутренней мембраны по
- 32. Механизм возникновения ПД: Важнейшая роль – ионы Na и K. При действии на клетку электрического тока
- 33. 4 Изменение возбудимости мембраны в различные фазы одиночного цикла возбуждения. Рефрактерность.
- 34. 0 – ПП 1 – деполяризация 2 – инверсия 3 – реполяризация 4 – медл реполяризация
- 35. 6 Рецепция: определение понятия. Биологическое значение. Рецептор: определение понятия, структура, виды. Морфологическая особенность клеточных и сенсорных
- 36. Рецепция (от лат. receptio — принятие) - это восприятие и преобразование (трансформация) механических, термических, электромагнитных, химических
- 37. Основные категории информационных сигналов. Химической природы: молекула вкусовых веществ; молекулы пахучих веществ; гормоны; нейромедиаторы; цитокины; факторы
- 38. Коммуникационные виды межклеточных контактов
- 39. Рецепторы Клеточные рецепторы Сенсорные рецепторы Рецептор – это генетически детерминированные макромолекулярные сенсоры (белки, глико-, липопротеины), локализованные
- 40. Предназначены: для специфического взаимодействия с сигналами химической или физической природы; для восприятия, трансформации и передачи информации,
- 41. Классификация рецепторов. Мембранные рецепторы: 7 ТМС рецепторы (7-TMSRs) 1 ТМС рецепторы (1-TMSRs) рецепторы со свойствами гуанилатциклазы
- 42. 7 ТМС рецепторы (более 250 видов). 7-TMSRs – 7 trans-membrane segment receptors или GPCRs – G-
- 43. Внеклеточная часть представлена гидрофильными петлями, с которыми взаимодействуют лиганды Трансмембранная часть представлена семью спиральными сегментами белковой
- 44. Важнейшая функция G- белка – дальнейшая передача внеклеточного сигнала на пострецепторные структуры, с помощью которых формируется
- 45. ЧСС (Ад) НА β-адр. Gs АЦ цАМФ ПКА гликогенолиз липолиз Gs - джи белок стимулирующая субъединица
- 46. 1 ТМС - (1 TMSRs – single transmembrane segment receptors) Белковая макромолекула, состоящая из трех частей:
- 47. Внеклеточный домен узнает и связывает молекулу-лиганд. Внутриклеточный домен не связан с G-белком и сам проявляет каталитическую
- 48. ЛЗИК – лиганд зависимый ионный канал – формируется белковыми макромолекулами, которые одновременно выполняют функцию ионных каналов
- 49. Внутриклеточные (цитозольные и ядерные) рецепторы. Лиганды – молекулы гидрофобной природы, легко проникают в клетку через плазматическую
- 50. Цитозольные рецепторы стероидных гормонов представлены белками цитоплазмы клетки, которые в отсутствие лиганда связаны с белками теплового
- 51. Рецепторами тиреоидных гормонов являются негистоновые белки, непосредственно в ядре клетки. Тиреоидные гормоны связываются с рецептором локализованным
- 52. 7 Понятие о сенсорных рецепторах. Общие механизмы рецепции. Кодирование информации в рецепторах. Рецепторный и генераторный потенциал.
- 53. Сенсорные рецепторы - это специализированные чувствительные клетки или нервные окончания чувствительных нейронов, которые воспринимают, преобразуют (кодируют),
- 54. Одной из общих особенностей механизмов восприятия и преобразования информационных сигналов в сенсорных рецепторах является то, что
- 55. КЛАССИФИКАЦИЯ СЕНСОРНЫХ РЕЦЕПТОРОВ (СР)
- 56. СТРУКТУРА сенсорных рецепторов. Они формируются: свободными нервными окончаниями дендритов чувствительных нейронов (болевые рецепторы, терморецепторы); 2) несвободными
- 57. ФУНКЦИИ СЕНСОРНЫХ РЕЦЕПТОРОВ (СР) Восприятие действия сигналов (раздражителей) и преобразование их энергии в электрические рецепторные потенциалы
- 58. Формирование РП. Тельца Фаттер-Пачини, капсула этих телец покрывает не миелинизированное нервное окончание, окружая его концентрическими пластинками
- 59. Механизм возникновения рецепторного потенциала (РП) При действии внешнего стимула на рецепторную клетку происходят конформационные изменения белковых
- 61. Скачать презентацию
Слайд 2Возбудимые клетки: их виды, свойства. Понятие о раздражимости, возбудимости и возбуждении, порогах
Возбудимые клетки: их виды, свойства. Понятие о раздражимости, возбудимости и возбуждении, порогах

Слайд 3Нейрон (возбудимость, проводимость)
Мышечная клетка (возбудимость, проводимость, сократимость)
Мышечная клетка (возбудимость, проводимость, сократимость)

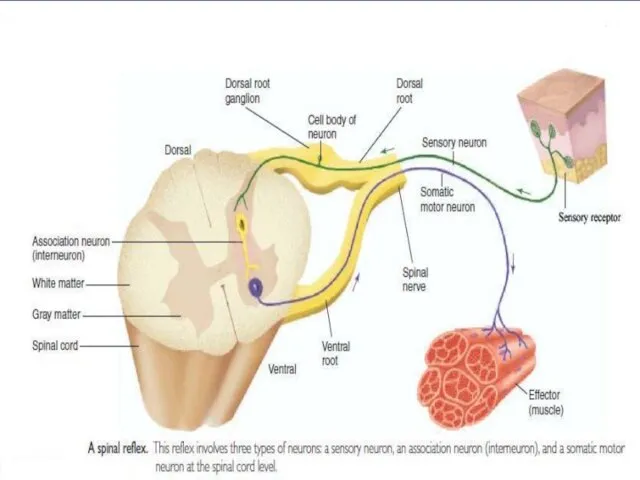
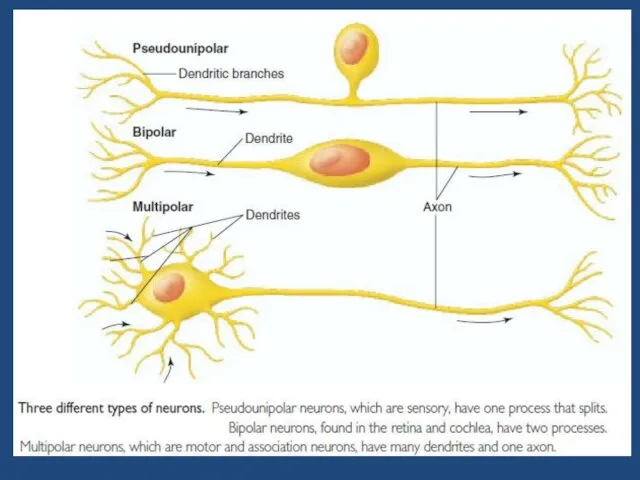
Слайд 6Раздражимость – это универсальное свойство клеток отвечать на действие раздражителя изменением процессов
Раздражимость – это универсальное свойство клеток отвечать на действие раздражителя изменением процессов

Слайд 7 Порог силы раздражителя - это минимальная величина силы действующего раздражителя, достаточная
Порог силы раздражителя - это минимальная величина силы действующего раздражителя, достаточная

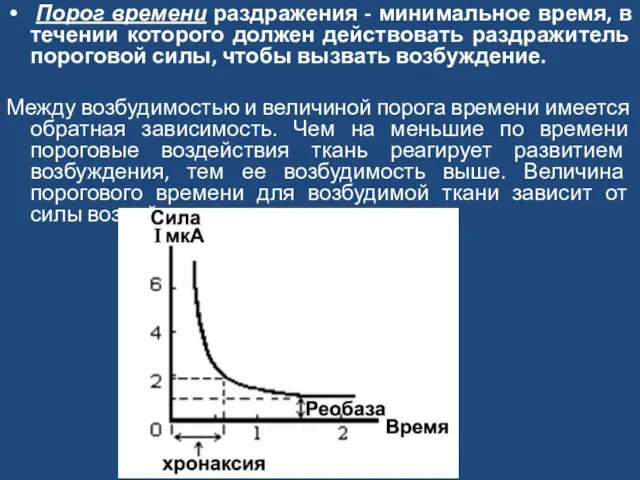
Слайд 8 Порог времени раздражения - минимальное время, в течении которого должен действовать
Порог времени раздражения - минимальное время, в течении которого должен действовать
Слайд 9Минимальный градиент (крутизна) нарастания силы раздражителя во времени. Это минимальная скорость увеличения
Минимальный градиент (крутизна) нарастания силы раздражителя во времени. Это минимальная скорость увеличения

Слайд 10 Хронаксия - минимальное время, в течение которого должен действовать раздражитель, равный
Хронаксия - минимальное время, в течение которого должен действовать раздражитель, равный

Слайд 11Адаптация – это приспособление к изменению условий внешней или внутренней среды, направленное
Адаптация – это приспособление к изменению условий внешней или внутренней среды, направленное

Слайд 12Лабильность - функциональная подвижность возбудимой ткани, зависящая от скорости элементарных физико-химических превращений,
Лабильность - функциональная подвижность возбудимой ткани, зависящая от скорости элементарных физико-химических превращений,

Слайд 132
Законы реагирования возбудимых клеток на действие раздражителей.
2
Законы реагирования возбудимых клеток на действие раздражителей.

Слайд 14Характер реагирования возбудимых тканей на действие раздражителей в классической физиологии принято описывать
Характер реагирования возбудимых тканей на действие раздражителей в классической физиологии принято описывать

Слайд 15Закон силы раздражения - утверждает, что при увеличении сверхпорогового раздражителя до определенного
Закон силы раздражения - утверждает, что при увеличении сверхпорогового раздражителя до определенного

Слайд 16Закон длительности раздражения – утверждает, что чем больше продолжительность сверхпорогового раздражения, тем
Закон длительности раздражения – утверждает, что чем больше продолжительность сверхпорогового раздражения, тем

Слайд 17Закон градиента раздражения – утверждает, что чем больше градиент нарастания силы раздражителя
Закон градиента раздражения – утверждает, что чем больше градиент нарастания силы раздражителя

Слайд 18Закон “все или ничего” – утверждает, что при действии подпороговых раздражителей возбуждение
Закон “все или ничего” – утверждает, что при действии подпороговых раздражителей возбуждение

Слайд 19Закон полярного действия тока (закон Пфлюгера) – при действии на возбудимые клетки
Закон полярного действия тока (закон Пфлюгера) – при действии на возбудимые клетки

Слайд 203
Современные представления о природе возбуждения. Биоэлектрогенез: потенциал покоя, локальный ответ, потенциал действия.
3
Современные представления о природе возбуждения. Биоэлектрогенез: потенциал покоя, локальный ответ, потенциал действия.

Слайд 21К возбудимым клеткам относят нервные и мышечные клетки, способные генерировать ПД.
Биопотенциалы (биоэлектрические
К возбудимым клеткам относят нервные и мышечные клетки, способные генерировать ПД.
Биопотенциалы (биоэлектрические

Слайд 22Роль биопотенциалов возбудимых тканей
Участие в механизмах восприятия воздействия различных факторов среды существования
2.
Роль биопотенциалов возбудимых тканей
Участие в механизмах восприятия воздействия различных факторов среды существования
2.

Слайд 23Потенциал покоя – это разность потенциалов между внутренней и наружной поверхностью мембраны
Потенциал покоя – это разность потенциалов между внутренней и наружной поверхностью мембраны



Слайд 26Потенциал действия – это кратко-временное высокоамплитудное колебание разности потенциалов мембраны возбудимой клетки,
Потенциал действия – это кратко-временное высокоамплитудное колебание разности потенциалов мембраны возбудимой клетки,

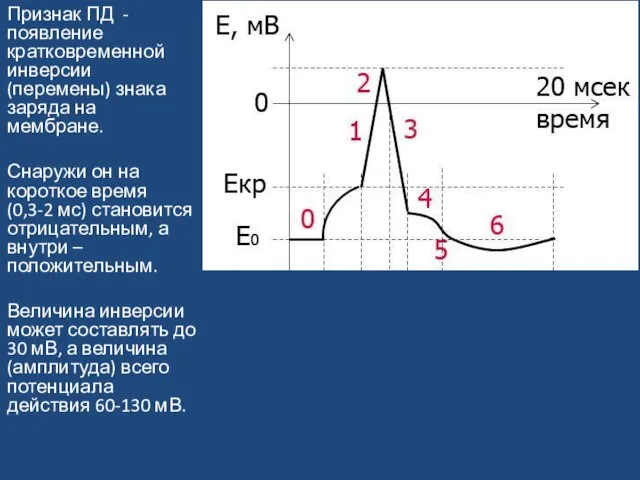
Слайд 27Признак ПД - появление кратковременной инверсии (перемены) знака заряда на мембране.
Снаружи он
Признак ПД - появление кратковременной инверсии (перемены) знака заряда на мембране.
Снаружи он
Слайд 28ПД подразделяется на 2 большие фазы:
- Деполяризации
- Реполяризации
- Деполяризации
- Реполяризации

Слайд 29Деполяризация – уменьшение электроотрицательности внутренней стороны мембраны, на графике вся восходящая часть,
Деполяризация – уменьшение электроотрицательности внутренней стороны мембраны, на графике вся восходящая часть,

Слайд 30Реполяризация – нисходящая часть ПД, которая отражает процесс восстановления исходной поляризации мембраны
Реполяризация – нисходящая часть ПД, которая отражает процесс восстановления исходной поляризации мембраны

Слайд 31Гиперполяризация – увеличение электроотрицательности внутренней стороны мембраны, от Е0 в сторону увеличения
Гиперполяризация – увеличение электроотрицательности внутренней стороны мембраны, от Е0 в сторону увеличения

Слайд 32Механизм возникновения ПД:
Важнейшая роль – ионы Na и K.
При действии на клетку
Механизм возникновения ПД:
Важнейшая роль – ионы Na и K.
При действии на клетку

Слайд 334
Изменение возбудимости мембраны в различные фазы одиночного цикла возбуждения. Рефрактерность.
4
Изменение возбудимости мембраны в различные фазы одиночного цикла возбуждения. Рефрактерность.

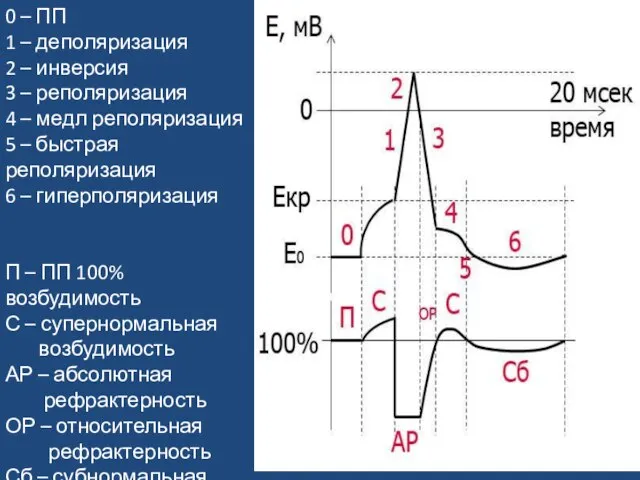
Слайд 340 – ПП
1 – деполяризация
2 – инверсия
3 – реполяризация
4 – медл
0 – ПП
1 – деполяризация
2 – инверсия
3 – реполяризация
4 – медл
Слайд 356
Рецепция: определение понятия. Биологическое значение. Рецептор: определение понятия, структура, виды. Морфологическая особенность
6
Рецепция: определение понятия. Биологическое значение. Рецептор: определение понятия, структура, виды. Морфологическая особенность

Слайд 36Рецепция (от лат. receptio — принятие) - это восприятие и преобразование (трансформация)
Рецепция (от лат. receptio — принятие) - это восприятие и преобразование (трансформация)

Слайд 37Основные категории информационных сигналов.
Химической природы:
молекула вкусовых веществ;
молекулы пахучих веществ;
гормоны;
нейромедиаторы;
цитокины;
факторы роста;
эйкозаноиды и другие
Основные категории информационных сигналов.
Химической природы:
молекула вкусовых веществ;
молекулы пахучих веществ;
гормоны;
нейромедиаторы;
цитокины;
факторы роста;
эйкозаноиды и другие

Слайд 38Коммуникационные виды межклеточных контактов
Коммуникационные виды межклеточных контактов

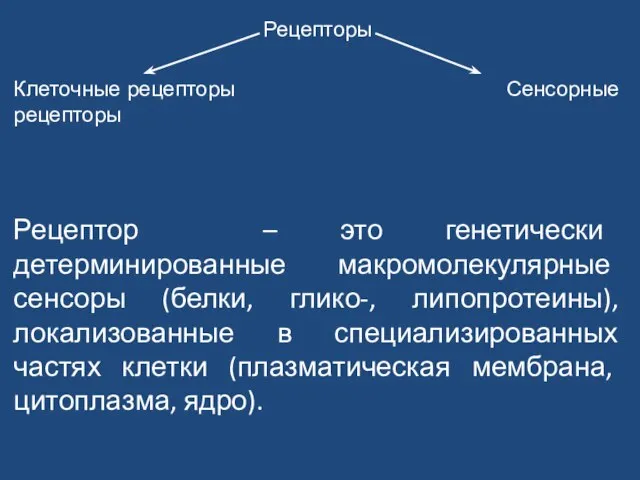
Слайд 39Рецепторы
Клеточные рецепторы Сенсорные рецепторы
Рецептор – это генетически детерминированные макромолекулярные сенсоры (белки, глико-,
Рецепторы
Клеточные рецепторы Сенсорные рецепторы
Рецептор – это генетически детерминированные макромолекулярные сенсоры (белки, глико-,
Слайд 40Предназначены:
для специфического взаимодействия с сигналами химической или физической природы;
для восприятия, трансформации и
Предназначены:
для специфического взаимодействия с сигналами химической или физической природы;
для восприятия, трансформации и

Слайд 41Классификация рецепторов.
Мембранные рецепторы:
7 ТМС рецепторы (7-TMSRs)
1 ТМС рецепторы (1-TMSRs)
рецепторы со свойствами гуанилатциклазы
рецепторы
Классификация рецепторов.
Мембранные рецепторы:
7 ТМС рецепторы (7-TMSRs)
1 ТМС рецепторы (1-TMSRs)
рецепторы со свойствами гуанилатциклазы
рецепторы

Слайд 427 ТМС рецепторы (более 250 видов).
7-TMSRs – 7 trans-membrane segment receptors или
7 ТМС рецепторы (более 250 видов).
7-TMSRs – 7 trans-membrane segment receptors или

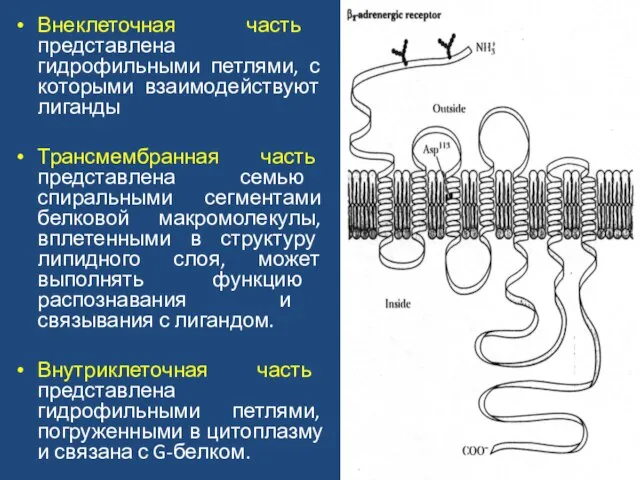
Слайд 43Внеклеточная часть представлена гидрофильными петлями, с которыми взаимодействуют лиганды
Трансмембранная часть представлена семью
Внеклеточная часть представлена гидрофильными петлями, с которыми взаимодействуют лиганды
Трансмембранная часть представлена семью
Слайд 44Важнейшая функция G- белка – дальнейшая передача внеклеточного сигнала на пострецепторные структуры,
Важнейшая функция G- белка – дальнейшая передача внеклеточного сигнала на пострецепторные структуры,

Слайд 45 ЧСС
(Ад) НА β-адр. Gs АЦ цАМФ ПКА гликогенолиз
липолиз
Gs - джи
ЧСС
(Ад) НА β-адр. Gs АЦ цАМФ ПКА гликогенолиз
липолиз
Gs - джи

Слайд 461 ТМС - (1 TMSRs – single transmembrane segment receptors)
Белковая макромолекула,
1 ТМС - (1 TMSRs – single transmembrane segment receptors)
Белковая макромолекула,

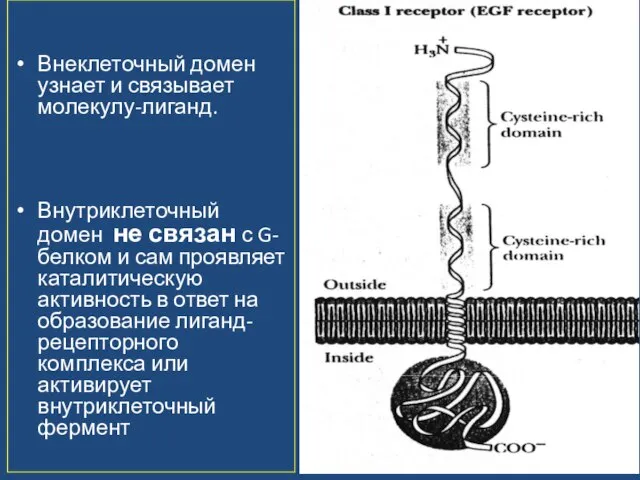
Слайд 47Внеклеточный домен узнает и связывает молекулу-лиганд.
Внутриклеточный домен не связан с G-белком и
Внутриклеточный домен не связан с G-белком и
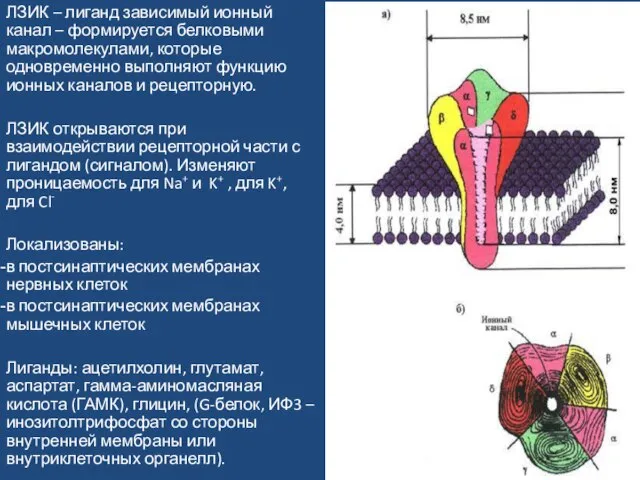
Слайд 48ЛЗИК – лиганд зависимый ионный канал – формируется белковыми макромолекулами, которые одновременно
ЛЗИК – лиганд зависимый ионный канал – формируется белковыми макромолекулами, которые одновременно
Слайд 49Внутриклеточные (цитозольные и ядерные) рецепторы.
Лиганды – молекулы гидрофобной природы, легко проникают в
Внутриклеточные (цитозольные и ядерные) рецепторы.
Лиганды – молекулы гидрофобной природы, легко проникают в

Слайд 50Цитозольные рецепторы стероидных гормонов представлены белками цитоплазмы клетки, которые в отсутствие лиганда
Цитозольные рецепторы стероидных гормонов представлены белками цитоплазмы клетки, которые в отсутствие лиганда

Слайд 51Рецепторами тиреоидных гормонов являются негистоновые белки, непосредственно в ядре клетки. Тиреоидные гормоны
Рецепторами тиреоидных гормонов являются негистоновые белки, непосредственно в ядре клетки. Тиреоидные гормоны

Слайд 527
Понятие о сенсорных рецепторах. Общие механизмы рецепции. Кодирование информации в рецепторах. Рецепторный
7
Понятие о сенсорных рецепторах. Общие механизмы рецепции. Кодирование информации в рецепторах. Рецепторный

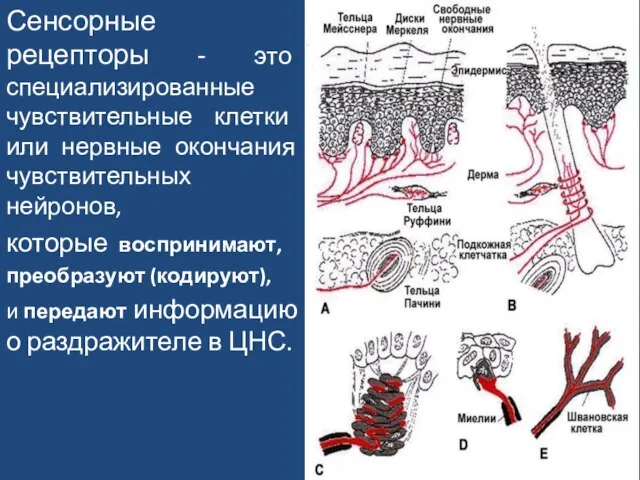
Слайд 53Сенсорные рецепторы - это специализированные чувствительные клетки или нервные окончания чувствительных нейронов,
Сенсорные рецепторы - это специализированные чувствительные клетки или нервные окончания чувствительных нейронов,
Слайд 54Одной из общих особенностей механизмов восприятия и преобразования информационных сигналов в сенсорных
Одной из общих особенностей механизмов восприятия и преобразования информационных сигналов в сенсорных

Слайд 55КЛАССИФИКАЦИЯ СЕНСОРНЫХ РЕЦЕПТОРОВ (СР)
КЛАССИФИКАЦИЯ СЕНСОРНЫХ РЕЦЕПТОРОВ (СР)

Слайд 56СТРУКТУРА сенсорных рецепторов.
Они формируются:
свободными нервными окончаниями дендритов чувствительных нейронов (болевые рецепторы,
СТРУКТУРА сенсорных рецепторов.
Они формируются:
свободными нервными окончаниями дендритов чувствительных нейронов (болевые рецепторы,

Слайд 57ФУНКЦИИ СЕНСОРНЫХ РЕЦЕПТОРОВ (СР)
Восприятие действия сигналов (раздражителей) и преобразование их энергии в
ФУНКЦИИ СЕНСОРНЫХ РЕЦЕПТОРОВ (СР)
Восприятие действия сигналов (раздражителей) и преобразование их энергии в

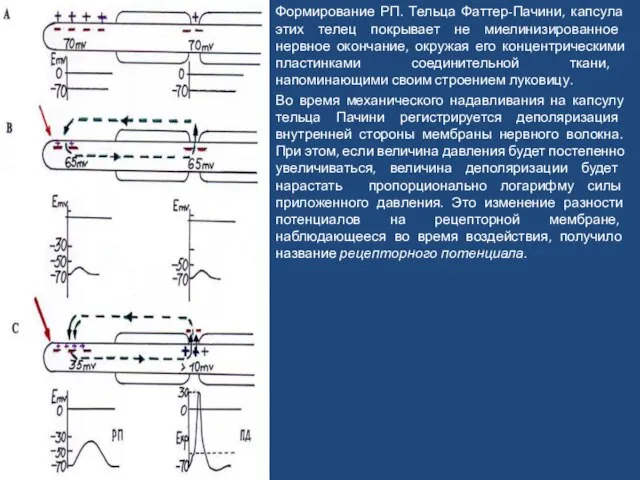
Слайд 58Формирование РП. Тельца Фаттер-Пачини, капсула этих телец покрывает не миелинизированное нервное окончание,
Формирование РП. Тельца Фаттер-Пачини, капсула этих телец покрывает не миелинизированное нервное окончание,
Слайд 59Механизм возникновения рецепторного потенциала (РП)
При действии внешнего стимула на рецепторную клетку происходят
Механизм возникновения рецепторного потенциала (РП)
При действии внешнего стимула на рецепторную клетку происходят

 Ecological problems in our country
Ecological problems in our country Фондовая биржа ММВБ: реалии и перспективы
Фондовая биржа ММВБ: реалии и перспективы Театрализованная игра как средство развития интонационной выразительности речи детей старшего дошкольного возраста
Театрализованная игра как средство развития интонационной выразительности речи детей старшего дошкольного возраста Семинар Международные стажировки и добровольчество
Семинар Международные стажировки и добровольчество Svenska helger
Svenska helger Влияние температуры на живые организмы
Влияние температуры на живые организмы Часть АА1 – А5
Часть АА1 – А5 Эффективное управление
Эффективное управление Ласло Мохой-Надь
Ласло Мохой-Надь Regiony turystyczne
Regiony turystyczne Чудеса изонити
Чудеса изонити 30 червня 2011 року м. Київ, УНЦПД РЕЗУЛЬТАТИ МОНІТОРИНГУ ІНФОРМАЦІЙНОГО НАПОВНЕННЯ ОФІЦІЙНИХ ВЕБ-СТОРІНОК МІСЦЕВИХ ДЕРЖАВНИХ АДМІНІ
30 червня 2011 року м. Київ, УНЦПД РЕЗУЛЬТАТИ МОНІТОРИНГУ ІНФОРМАЦІЙНОГО НАПОВНЕННЯ ОФІЦІЙНИХ ВЕБ-СТОРІНОК МІСЦЕВИХ ДЕРЖАВНИХ АДМІНІ Тряпичная или керамическая кукла Художкин
Тряпичная или керамическая кукла Художкин Презентация на тему виды пейзажа
Презентация на тему виды пейзажа  ИКТ в образовательном пространстве МОУ «СОШ №3» г. Заозерного
ИКТ в образовательном пространстве МОУ «СОШ №3» г. Заозерного Значение бактерий в природе и жизни человека - презентация по Биологии
Значение бактерий в природе и жизни человека - презентация по Биологии Выпускная квалификационная работа социальная адаптация детей-сирот в условиях колледжа
Выпускная квалификационная работа социальная адаптация детей-сирот в условиях колледжа Шурави – Герои Афганской войны 25.12.1979-15.02.1989гг
Шурави – Герои Афганской войны 25.12.1979-15.02.1989гг А.Н.Островский "Бедность не порок"
А.Н.Островский "Бедность не порок" Классификация учебных мотивов по М.В. Матюхиной. Классификация учебных мотивов по: познавательные и социальные мотивы
Классификация учебных мотивов по М.В. Матюхиной. Классификация учебных мотивов по: познавательные и социальные мотивы Древние мыслители о мире и человеке
Древние мыслители о мире и человеке Проектная работа Тема: «Я памятник себе воздвиг нерукотворный…» (сопоставительный анализ стихов-завещаний Горация, М.В.Ломоносов
Проектная работа Тема: «Я памятник себе воздвиг нерукотворный…» (сопоставительный анализ стихов-завещаний Горация, М.В.Ломоносов Социальное взаимодействие в строительстве
Социальное взаимодействие в строительстве Серебряная защита вашего здоровья
Серебряная защита вашего здоровья Презентация на тему Биография Платонова
Презентация на тему Биография Платонова Заочное путешествие по святым местам Белгородской области
Заочное путешествие по святым местам Белгородской области Российские и международные гранты для молодых ученых:
Российские и международные гранты для молодых ученых: Презентация на тему Современные мастера, прославившие Россию в XVIII-XXI веках
Презентация на тему Современные мастера, прославившие Россию в XVIII-XXI веках