- ПЕРВИЧНАЯ СТРУКТУРА БЕЛКА

Содержание
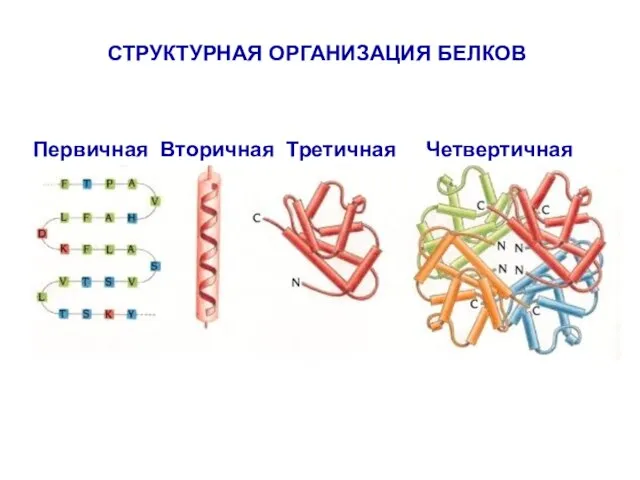
- 2. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ БЕЛКОВ Первичная Вторичная Третичная Четвертичная
- 3. Белки представляют собой высокомолекулярные биополимеры, построенные из аминокислотных остатков. Практически все белки построены из 22 аминокислот.
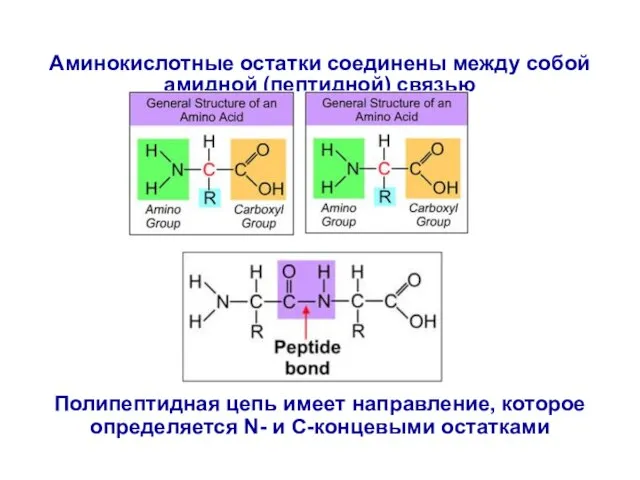
- 4. Аминокислотные остатки соединены между собой амидной (пептидной) связью Полипептидная цепь имеет направление, которое определяется N- и
- 5. ПРИМЕРЫ ПОСТТРАНСЛЯЦИОННЫХ МОДИФИКАЦИЙ БЕЛКОВ Lys: метилирование, ацетилирование Arg: метилирование, дезаминирование His: метилирование Glu: карбоксилирование Asp: образование
- 6. ОСНОВНЫЕ ЭТАПЫ ОПРЕДЕЛЕНИЯ ПЕРВИЧНОЙ СТРУКТУРЫ БЕЛКА ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ ОПРЕДЕЛЕНИЕ АМИНОКИСЛОТНОГО СОСТАВА (АМИНОКИСЛОТНЫЙ АНАЛИЗ) ОПРЕДЕЛЕНИЕ N-
- 7. ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ МАСС-СПЕКТРОМЕТРИЯ ЭЛЕКТРОФОРЕЗ 3. ГЕЛЬ-ФИЛЬТРАЦИЯ 4. УЛЬТРАЦЕНТРИФУГИРОВАНИЕ
- 8. Принцип аминокислотного анализа Гидролиз в 5,7н HCl при 1100C в вакууме в течение 24-96 часов
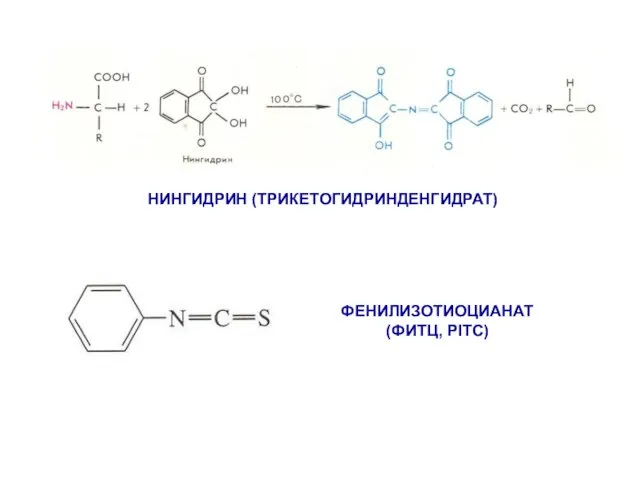
- 9. ФЕНИЛИЗОТИОЦИАНАТ (ФИТЦ, PITC) НИНГИДРИН (ТРИКЕТОГИДРИНДЕНГИДРАТ)
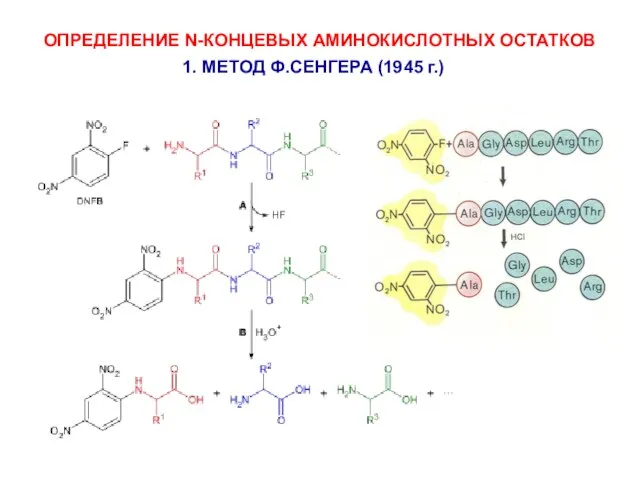
- 10. ОПРЕДЕЛЕНИЕ N-КОНЦЕВЫХ АМИНОКИСЛОТНЫХ ОСТАТКОВ 1. МЕТОД Ф.СЕНГЕРА (1945 г.)
- 11. 2. ДАНСИЛЬНЫЙ МЕТОД (В.Р.ГРЕЙ, Б.С.ХАРТЛИ, 1963 г.)
- 12. 3. МЕТОД ЭДМАНА (1956 г.) ФЕНИЛИЗОТИОЦИАНАТ (ФИТЦ, PITC, РЕАГЕНТ ЭДМАНА)
- 13. 4. ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ АМИНОПЕПТИДАЗАМИ ЛЕЙЦИНАМИНОПЕПТИДАЗА (С НАИБОЛЬШЕЙ СКОРОСТЬЮ ГИДРОЛИЗУЕТ СВЯЗИ Leu, Ile, Val) АМИНОПЕПТИДАЗА М (ГИДРОЛИЗУЕТ
- 14. ОПРЕДЕЛЕНИЕ С-КОНЦЕВЫХ АМИНОКИСЛОТНЫХ ОСТАТКОВ МЕТОД ГИДРАЗИНОЛИЗА (Сиро АКАБОРИ, 1952 г.) Безводный NH2-NH2, 1200C, 10 час
- 15. 2. ОКСАЗОЛОНОВЫЙ МЕТОД ПО МАТСУО ИЛИ МЕТОД ТРИТИЕВОЙ МЕТКИ (Hisayuki MATSUO, 1965 г.)
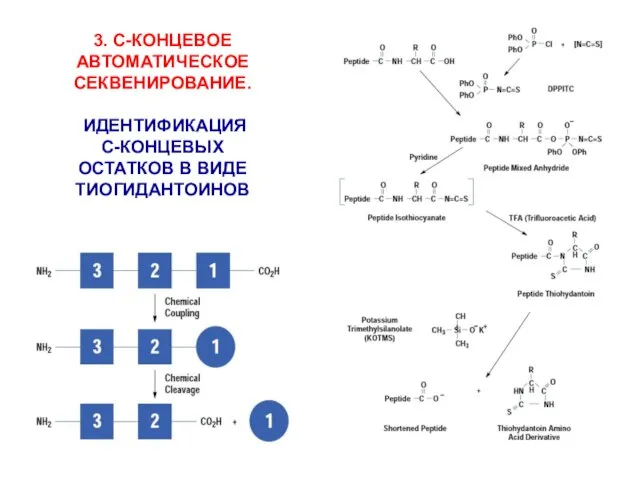
- 16. 3. С-КОНЦЕВОЕ АВТОМАТИЧЕСКОЕ СЕКВЕНИРОВАНИЕ. ИДЕНТИФИКАЦИЯ С-КОНЦЕВЫХ ОСТАТКОВ В ВИДЕ ТИОГИДАНТОИНОВ
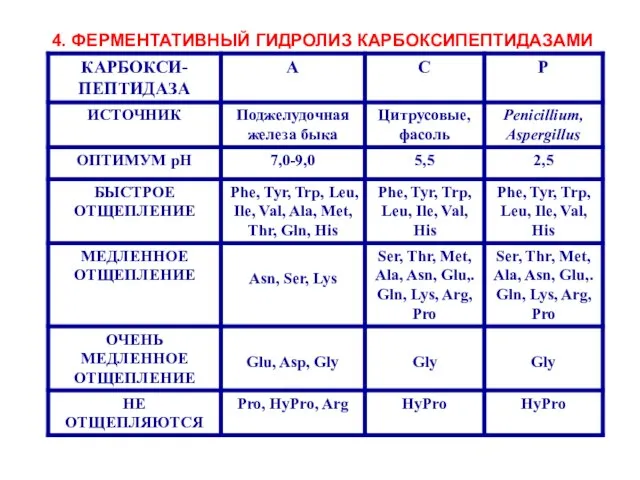
- 17. 4. ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ КАРБОКСИПЕПТИДАЗАМИ
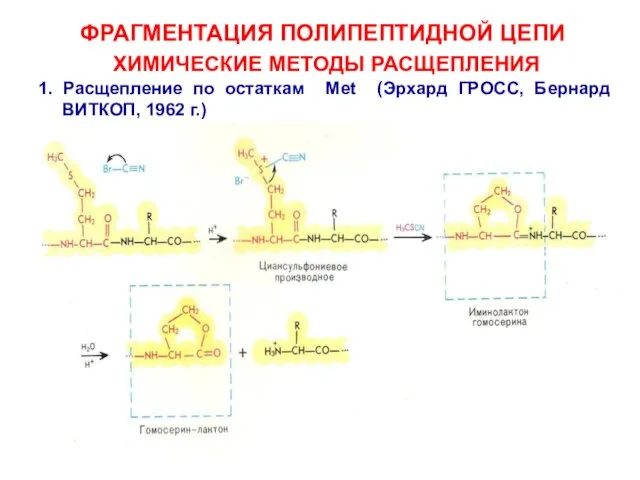
- 18. ФРАГМЕНТАЦИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ ХИМИЧЕСКИЕ МЕТОДЫ РАСЩЕПЛЕНИЯ 1. Расщепление по остаткам Met (Эрхард ГРОСС, Бернард ВИТКОП, 1962
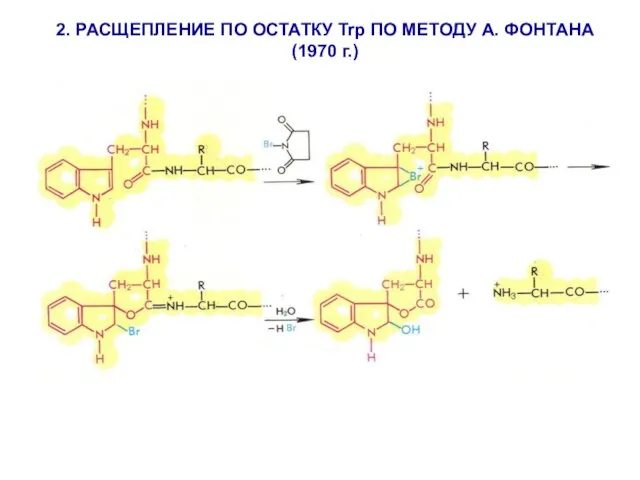
- 19. 2. РАСЩЕПЛЕНИЕ ПО ОСТАТКУ Trp ПО МЕТОДУ А. ФОНТАНА (1970 г.)
- 20. 2-(2-нитрофенилсульфенил)-3-метил-3-броминдол (скатол = 3-метилиндол)
- 21. 3. РАСЩЕПЛЕНИЕ СВЯЗИ Asn-Gly
- 22. 2-иминотиозолидинкарбоновая кислота 5. РАСЩЕПЛЕНИЕ ПО ОСТАТКАМ Cys (А. ПАТЧОРНИК, 1974 г.) 2-нитро-5-тиоцианатбензойная кислота
- 23. ФЕРМЕНТАТИВНЫЕ МЕТОДЫ ФРАГМЕНТАЦИИ ПОЛИПЕПТИДНОЙ ЦЕПИ ВЫСОКАЯ СПЕЦИФИЧНОСТЬ 1. Трипсин pH 7,0-9,0 Lys—X Arg—X Lys—Pro Arg—Pro Протеаза
- 24. ХИМОТРИПСИН ТЕРМОЛИЗИН pH 7,8 – 9,0 С МЕНЬШЕЙ СКОРОСТЬЮ pH 7,0 – 9,0 С МЕНЬШЕЙ СКОРОСТЬЮ
- 25. ЛИЗИН-СПЕЦИФИЧЕСКАЯ ПРОТЕАЗА ИЗ Armillaria mellea pH 8,0 ЭНДОПРОТЕИНАЗА Lys-C ИЗ Lysobacter ensymogenes КЛОСТРИПАИН ИЗ Clostridium histolyticum
- 27. Скачать презентацию
Слайд 3Белки представляют собой высокомолекулярные биополимеры, построенные из аминокислотных остатков.
Практически все белки построены
Белки представляют собой высокомолекулярные биополимеры, построенные из аминокислотных остатков.
Практически все белки построены

Слайд 4Аминокислотные остатки соединены между собой амидной (пептидной) связью
Полипептидная цепь имеет направление,
Аминокислотные остатки соединены между собой амидной (пептидной) связью
Полипептидная цепь имеет направление,
Слайд 5ПРИМЕРЫ ПОСТТРАНСЛЯЦИОННЫХ МОДИФИКАЦИЙ БЕЛКОВ
Lys: метилирование, ацетилирование
Arg: метилирование, дезаминирование
His: метилирование
Glu: карбоксилирование
Asp:
ПРИМЕРЫ ПОСТТРАНСЛЯЦИОННЫХ МОДИФИКАЦИЙ БЕЛКОВ
Lys: метилирование, ацетилирование
Arg: метилирование, дезаминирование
His: метилирование
Glu: карбоксилирование
Asp:

Слайд 6ОСНОВНЫЕ ЭТАПЫ ОПРЕДЕЛЕНИЯ
ПЕРВИЧНОЙ СТРУКТУРЫ БЕЛКА
ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ
ОПРЕДЕЛЕНИЕ АМИНОКИСЛОТНОГО СОСТАВА (АМИНОКИСЛОТНЫЙ АНАЛИЗ)
ОПРЕДЕЛЕНИЕ
ОСНОВНЫЕ ЭТАПЫ ОПРЕДЕЛЕНИЯ
ПЕРВИЧНОЙ СТРУКТУРЫ БЕЛКА
ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ
ОПРЕДЕЛЕНИЕ АМИНОКИСЛОТНОГО СОСТАВА (АМИНОКИСЛОТНЫЙ АНАЛИЗ)
ОПРЕДЕЛЕНИЕ

Слайд 7ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ
МАСС-СПЕКТРОМЕТРИЯ
ЭЛЕКТРОФОРЕЗ
3. ГЕЛЬ-ФИЛЬТРАЦИЯ
4. УЛЬТРАЦЕНТРИФУГИРОВАНИЕ
ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ
МАСС-СПЕКТРОМЕТРИЯ
ЭЛЕКТРОФОРЕЗ
3. ГЕЛЬ-ФИЛЬТРАЦИЯ
4. УЛЬТРАЦЕНТРИФУГИРОВАНИЕ

Слайд 8Принцип
аминокислотного
анализа
Гидролиз в 5,7н HCl
при 1100C в вакууме
в течение 24-96
Принцип
аминокислотного
анализа
Гидролиз в 5,7н HCl
при 1100C в вакууме
в течение 24-96

Слайд 9ФЕНИЛИЗОТИОЦИАНАТ
(ФИТЦ, PITC)
НИНГИДРИН (ТРИКЕТОГИДРИНДЕНГИДРАТ)
ФЕНИЛИЗОТИОЦИАНАТ
(ФИТЦ, PITC)
НИНГИДРИН (ТРИКЕТОГИДРИНДЕНГИДРАТ)
Слайд 10ОПРЕДЕЛЕНИЕ N-КОНЦЕВЫХ АМИНОКИСЛОТНЫХ ОСТАТКОВ
1. МЕТОД Ф.СЕНГЕРА (1945 г.)
ОПРЕДЕЛЕНИЕ N-КОНЦЕВЫХ АМИНОКИСЛОТНЫХ ОСТАТКОВ
1. МЕТОД Ф.СЕНГЕРА (1945 г.)
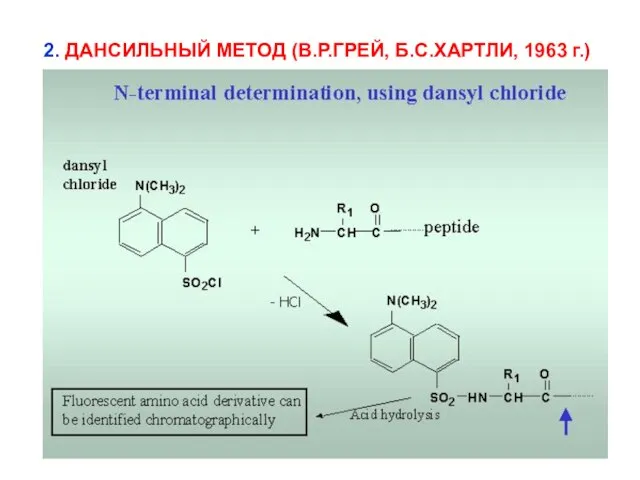
Слайд 112. ДАНСИЛЬНЫЙ МЕТОД (В.Р.ГРЕЙ, Б.С.ХАРТЛИ, 1963 г.)
2. ДАНСИЛЬНЫЙ МЕТОД (В.Р.ГРЕЙ, Б.С.ХАРТЛИ, 1963 г.)
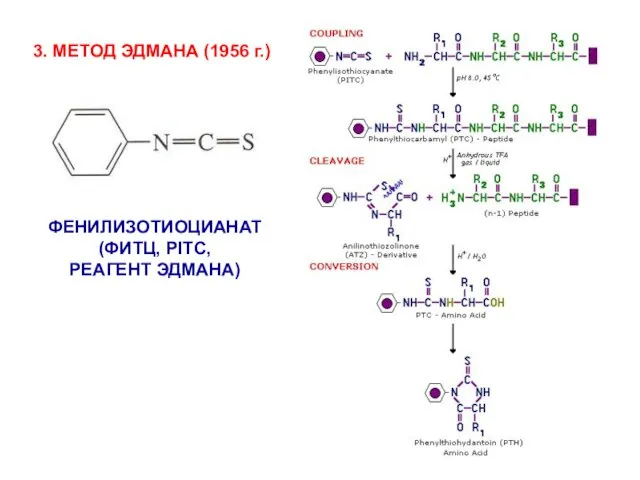
Слайд 123. МЕТОД ЭДМАНА (1956 г.)
ФЕНИЛИЗОТИОЦИАНАТ
(ФИТЦ, PITC,
РЕАГЕНТ ЭДМАНА)
3. МЕТОД ЭДМАНА (1956 г.)
ФЕНИЛИЗОТИОЦИАНАТ
(ФИТЦ, PITC,
РЕАГЕНТ ЭДМАНА)
Слайд 134. ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ АМИНОПЕПТИДАЗАМИ
ЛЕЙЦИНАМИНОПЕПТИДАЗА (С НАИБОЛЬШЕЙ СКОРОСТЬЮ ГИДРОЛИЗУЕТ СВЯЗИ Leu, Ile, Val)
АМИНОПЕПТИДАЗА
4. ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ АМИНОПЕПТИДАЗАМИ
ЛЕЙЦИНАМИНОПЕПТИДАЗА (С НАИБОЛЬШЕЙ СКОРОСТЬЮ ГИДРОЛИЗУЕТ СВЯЗИ Leu, Ile, Val)
АМИНОПЕПТИДАЗА

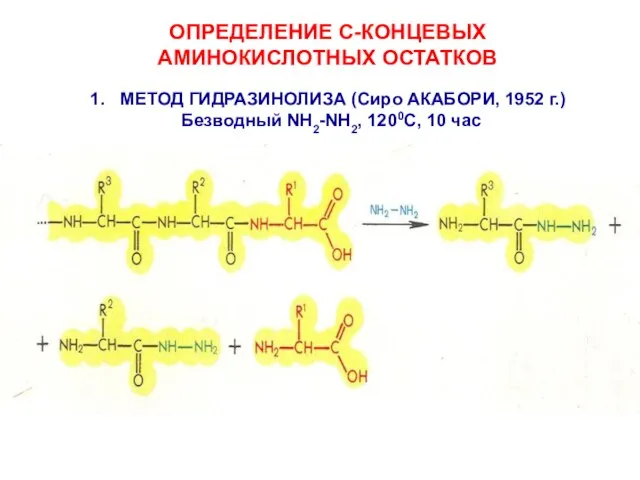
Слайд 14ОПРЕДЕЛЕНИЕ С-КОНЦЕВЫХ
АМИНОКИСЛОТНЫХ ОСТАТКОВ
МЕТОД ГИДРАЗИНОЛИЗА (Сиро АКАБОРИ, 1952 г.)
Безводный NH2-NH2, 1200C, 10
ОПРЕДЕЛЕНИЕ С-КОНЦЕВЫХ
АМИНОКИСЛОТНЫХ ОСТАТКОВ
МЕТОД ГИДРАЗИНОЛИЗА (Сиро АКАБОРИ, 1952 г.)
Безводный NH2-NH2, 1200C, 10
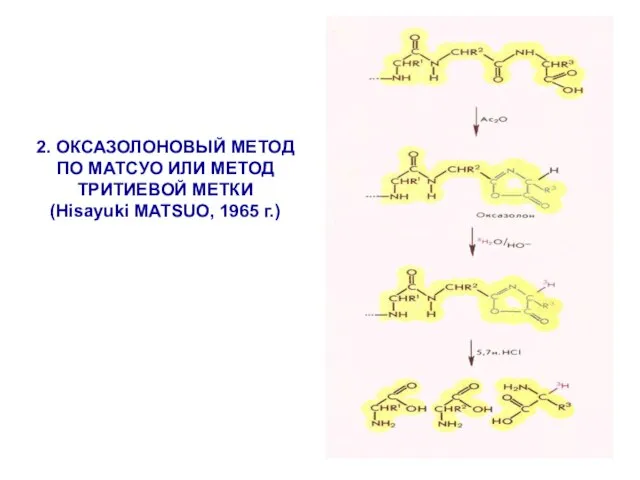
Слайд 152. ОКСАЗОЛОНОВЫЙ МЕТОД
ПО МАТСУО ИЛИ МЕТОД ТРИТИЕВОЙ МЕТКИ
(Hisayuki MATSUO, 1965
2. ОКСАЗОЛОНОВЫЙ МЕТОД
ПО МАТСУО ИЛИ МЕТОД ТРИТИЕВОЙ МЕТКИ
(Hisayuki MATSUO, 1965
Слайд 163. С-КОНЦЕВОЕ
АВТОМАТИЧЕСКОЕ
СЕКВЕНИРОВАНИЕ.
ИДЕНТИФИКАЦИЯ
С-КОНЦЕВЫХ ОСТАТКОВ В ВИДЕ ТИОГИДАНТОИНОВ
3. С-КОНЦЕВОЕ
АВТОМАТИЧЕСКОЕ
СЕКВЕНИРОВАНИЕ.
ИДЕНТИФИКАЦИЯ
С-КОНЦЕВЫХ ОСТАТКОВ В ВИДЕ ТИОГИДАНТОИНОВ
Слайд 174. ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ КАРБОКСИПЕПТИДАЗАМИ
4. ФЕРМЕНТАТИВНЫЙ ГИДРОЛИЗ КАРБОКСИПЕПТИДАЗАМИ
Слайд 18ФРАГМЕНТАЦИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ
ХИМИЧЕСКИЕ МЕТОДЫ РАСЩЕПЛЕНИЯ
1. Расщепление по остаткам Met (Эрхард ГРОСС, Бернард
ФРАГМЕНТАЦИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ
ХИМИЧЕСКИЕ МЕТОДЫ РАСЩЕПЛЕНИЯ
1. Расщепление по остаткам Met (Эрхард ГРОСС, Бернард
Слайд 192. РАСЩЕПЛЕНИЕ ПО ОСТАТКУ Trp ПО МЕТОДУ А. ФОНТАНА
(1970 г.)
2. РАСЩЕПЛЕНИЕ ПО ОСТАТКУ Trp ПО МЕТОДУ А. ФОНТАНА
(1970 г.)
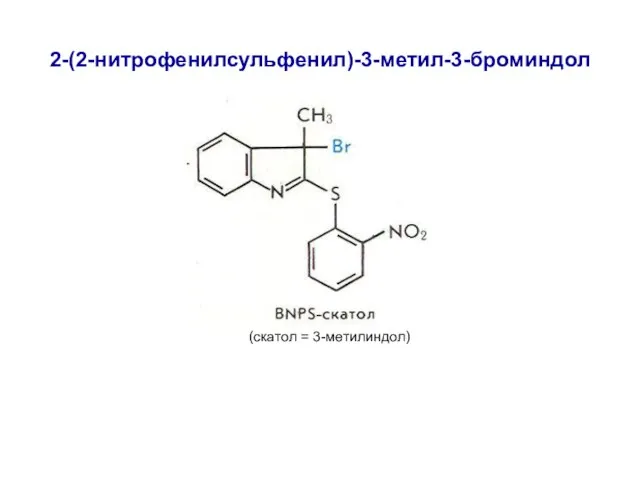
Слайд 202-(2-нитрофенилсульфенил)-3-метил-3-броминдол
(скатол = 3-метилиндол)
2-(2-нитрофенилсульфенил)-3-метил-3-броминдол
(скатол = 3-метилиндол)
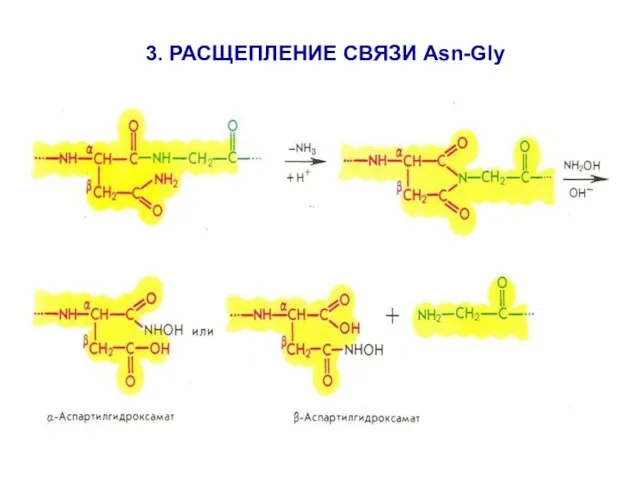
Слайд 213. РАСЩЕПЛЕНИЕ СВЯЗИ Asn-Gly
3. РАСЩЕПЛЕНИЕ СВЯЗИ Asn-Gly
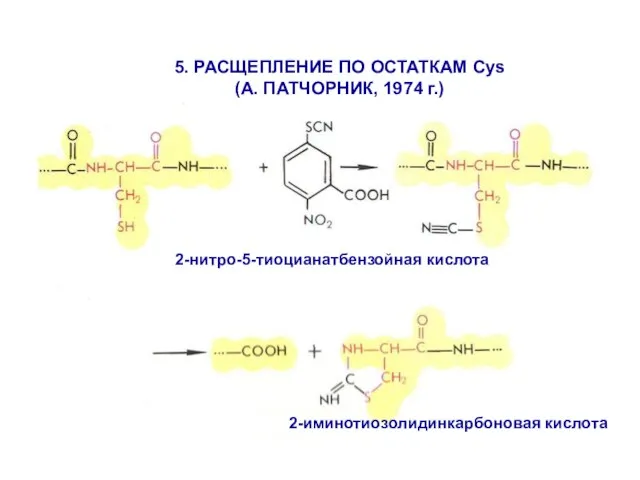
Слайд 222-иминотиозолидинкарбоновая кислота
5. РАСЩЕПЛЕНИЕ ПО ОСТАТКАМ Cys
(А. ПАТЧОРНИК, 1974 г.)
2-нитро-5-тиоцианатбензойная кислота
2-иминотиозолидинкарбоновая кислота
5. РАСЩЕПЛЕНИЕ ПО ОСТАТКАМ Cys
(А. ПАТЧОРНИК, 1974 г.)
2-нитро-5-тиоцианатбензойная кислота
Слайд 23ФЕРМЕНТАТИВНЫЕ МЕТОДЫ ФРАГМЕНТАЦИИ ПОЛИПЕПТИДНОЙ ЦЕПИ
ВЫСОКАЯ СПЕЦИФИЧНОСТЬ
1. Трипсин pH 7,0-9,0
Lys—X Arg—X
ФЕРМЕНТАТИВНЫЕ МЕТОДЫ ФРАГМЕНТАЦИИ ПОЛИПЕПТИДНОЙ ЦЕПИ
ВЫСОКАЯ СПЕЦИФИЧНОСТЬ
1. Трипсин pH 7,0-9,0
Lys—X Arg—X

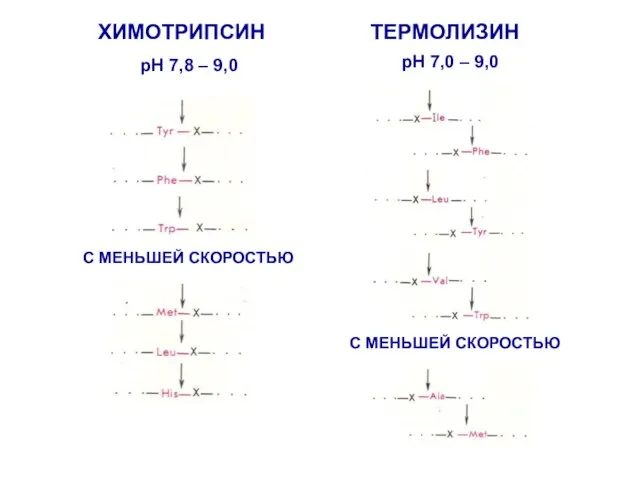
Слайд 24ХИМОТРИПСИН
ТЕРМОЛИЗИН
pH 7,8 – 9,0
С МЕНЬШЕЙ СКОРОСТЬЮ
pH 7,0 – 9,0
С МЕНЬШЕЙ СКОРОСТЬЮ
ХИМОТРИПСИН
ТЕРМОЛИЗИН
pH 7,8 – 9,0
С МЕНЬШЕЙ СКОРОСТЬЮ
pH 7,0 – 9,0
С МЕНЬШЕЙ СКОРОСТЬЮ
Слайд 25ЛИЗИН-СПЕЦИФИЧЕСКАЯ
ПРОТЕАЗА ИЗ Armillaria mellea
pH 8,0
ЭНДОПРОТЕИНАЗА Lys-C ИЗ
Lysobacter ensymogenes
КЛОСТРИПАИН
ИЗ Clostridium histolyticum
pH 7,7
pH
ЛИЗИН-СПЕЦИФИЧЕСКАЯ
ПРОТЕАЗА ИЗ Armillaria mellea
pH 8,0
ЭНДОПРОТЕИНАЗА Lys-C ИЗ
Lysobacter ensymogenes
КЛОСТРИПАИН
ИЗ Clostridium histolyticum
pH 7,7
pH

 Формирование команды. Роли в команде
Формирование команды. Роли в команде Испания - Мадрид
Испания - Мадрид Графики вокруг нас
Графики вокруг нас Языковая программа: Мальта, 2018
Языковая программа: Мальта, 2018 Вечно холодные, вчено свободные
Вечно холодные, вчено свободные Любава. Новинки: ассортимент печенья, ассортимент готовых завтраков
Любава. Новинки: ассортимент печенья, ассортимент готовых завтраков Аттестация на должность инженера I категории. Волгин Евгений Алексеевич
Аттестация на должность инженера I категории. Волгин Евгений Алексеевич МОУ «Основная общеобразовательная школа с.Раевка Ивантеевского района Саратовской области» «Использование методов исследовани
МОУ «Основная общеобразовательная школа с.Раевка Ивантеевского района Саратовской области» «Использование методов исследовани Презентация без названия
Презентация без названия Ораторы. Голос
Ораторы. Голос РЕШЕНИЕ ЗАДАЧ
РЕШЕНИЕ ЗАДАЧ Карл Линней
Карл Линней Презентация на тему Знакомство с буквой С,с. Письмо буквы С,с
Презентация на тему Знакомство с буквой С,с. Письмо буквы С,с Решение задач с помощью квадратных и рациональных уравнений
Решение задач с помощью квадратных и рациональных уравнений ОПЫТ ПРОЕКТИРОВАНИЯ ЭФФЕКТИВНЫХ КОММУНАЛЬНЫХ СИСТЕМ. ТЕПЛО.СВЕТ.
ОПЫТ ПРОЕКТИРОВАНИЯ ЭФФЕКТИВНЫХ КОММУНАЛЬНЫХ СИСТЕМ. ТЕПЛО.СВЕТ. "Гений русской науки"
"Гений русской науки" Психология групп С.А. Володина. Малые группы
Психология групп С.А. Володина. Малые группы ВЕЛИКИЙ И МОГУЧИЙ
ВЕЛИКИЙ И МОГУЧИЙ Агентский договор
Агентский договор Александр Азимов
Александр Азимов  Внутриорганизационные конфликты
Внутриорганизационные конфликты Простые и составные числительные
Простые и составные числительные Эпоха Великих географических открытий
Эпоха Великих географических открытий Презентация на тему Виды личного страхования и их характеристика
Презентация на тему Виды личного страхования и их характеристика  Конкурс Выбери мою школу
Конкурс Выбери мою школу ПРЕЗЕНТАЦИЯ
ПРЕЗЕНТАЦИЯ Общественное наблюдение за ходом голосования и подсчетом голосов избирателей
Общественное наблюдение за ходом голосования и подсчетом голосов избирателей Отделение испытаний 04
Отделение испытаний 04