- Питание бактерий

Содержание
- 2. Под питанием понимают процессы поступления и выведения питательных веществ в клетку и из клетки. Питание в
- 3. Для бактерий характерно многообразие источников получения питательных веществ. В зависимости от источника получения углерода бактерии делят
- 4. Процессы питания должны обеспечивать энергетические потребности бактериальной клетки. По источникам энергии микроорганизмы делят на: 1) фототрофы
- 5. По степени гетеротрофности микроорганизмы делятся на: 1) сапрофиты – питаются мертвым органическим материалом; 2) паразиты –
- 6. Факторами роста бактерий являются витамины, аминокислоты, пуриновые и пиримидиновые основания, присутствие которых ускоряет рост. Среди бактерий
- 7. Микроорганизмы ассимилируют питательные вещества в виде небольших молекул, поэтому белки, полисахариды и другие биополимеры могут служить
- 8. Транспорт веществ
- 9. Для того, чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки, они должны проникнуть в клетку
- 10. Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.
- 11. При пассивном переносе вещество проникает в клетку только по градиенту концентрации. Затрат энергии при этом не
- 12. Простая диффузия — неспецифическое проникновение веществ в клетку, при этом решающее значение имеет величина молекул и
- 13. Облегченная диффузия протекает с участием белка-переносчика. Скорость этого способа переноса зависит от концентрации вещества в наружном
- 14. При активном переносе вещество проникает в клетку против градиента концентрации при помощи белка-переносчика — пермеазы. При
- 15. При одном типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) «накачиваются» в клетку и создают концентрацию,
- 16. Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система, составной частью которой является белок-переносчик,
- 17. Поступив в клетку, органический источник углерода и энергии вступает в цепь биохимических реакций, в результате которых
- 18. Конструктивный метаболизм
- 19. Основными видами пластического обмена являются: 1) белковый; 2) углеводный; 3) липидный; 4) нуклеиновый.
- 20. Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают белки под действием протеаз с
- 21. Биосинтез аминокислот. Распад белков в аэробных условиях называется тлением, в анаэробных – гниением. В результате распада
- 22. Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные продукты фруктозодифосфатного (ФДФ) и
- 23. Биосинтез нуклеотидов. Пуриновые и пиримидиновые нуклеотиды — это те строительные блоки, из которых синтезируются нуклеиновые кислоты.
- 24. Биосинтез жиров. Жиры или липиды являются важными компонентами ЦПМ и клеточной стенки грамотрицательных бактерий, а также
- 25. Биосинтез углеводов. Углеводы представлены в бактериальной клетке в виде моно-, ди- и полисахаридов, а также комплексных
- 27. Скачать презентацию

Слайд 3Для бактерий характерно многообразие источников получения питательных веществ.
В зависимости от источника получения
Для бактерий характерно многообразие источников получения питательных веществ.
В зависимости от источника получения

Слайд 4Процессы питания должны обеспечивать энергетические потребности бактериальной клетки. По источникам энергии микроорганизмы
Процессы питания должны обеспечивать энергетические потребности бактериальной клетки. По источникам энергии микроорганизмы

Слайд 5По степени гетеротрофности микроорганизмы делятся на:
1) сапрофиты – питаются мертвым органическим
По степени гетеротрофности микроорганизмы делятся на:
1) сапрофиты – питаются мертвым органическим

Слайд 6Факторами роста бактерий являются витамины, аминокислоты, пуриновые и пиримидиновые основания, присутствие которых
Факторами роста бактерий являются витамины, аминокислоты, пуриновые и пиримидиновые основания, присутствие которых

Слайд 7Микроорганизмы ассимилируют питательные вещества в виде небольших молекул, поэтому белки, полисахариды и
Микроорганизмы ассимилируют питательные вещества в виде небольших молекул, поэтому белки, полисахариды и

Слайд 8Транспорт веществ
Транспорт веществ

Слайд 9Для того, чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки, они
Для того, чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки, они

Слайд 10Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.
Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.

Слайд 11При пассивном переносе вещество проникает в клетку только по градиенту концентрации. Затрат
При пассивном переносе вещество проникает в клетку только по градиенту концентрации. Затрат

Слайд 12Простая диффузия — неспецифическое проникновение веществ в клетку, при этом решающее значение
Простая диффузия — неспецифическое проникновение веществ в клетку, при этом решающее значение

Слайд 13Облегченная диффузия протекает с участием белка-переносчика. Скорость этого способа переноса зависит от
Облегченная диффузия протекает с участием белка-переносчика. Скорость этого способа переноса зависит от

Слайд 14При активном переносе вещество проникает в клетку против градиента концентрации при помощи
При активном переносе вещество проникает в клетку против градиента концентрации при помощи

Слайд 15При одном типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) «накачиваются» в
При одном типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) «накачиваются» в

Слайд 16Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система, составной
Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система, составной

Слайд 17Поступив в клетку, органический источник углерода и энергии вступает в цепь биохимических
Поступив в клетку, органический источник углерода и энергии вступает в цепь биохимических

Слайд 18Конструктивный метаболизм
Конструктивный метаболизм

Слайд 19Основными видами пластического обмена являются:
1) белковый;
2) углеводный;
3) липидный;
4)
Основными видами пластического обмена являются:
1) белковый;
2) углеводный;
3) липидный;
4)

Слайд 20Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают белки
Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают белки

Слайд 21Биосинтез аминокислот.
Распад белков в аэробных условиях называется тлением, в анаэробных –
Биосинтез аминокислот.
Распад белков в аэробных условиях называется тлением, в анаэробных –

Слайд 22Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные
Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные

Слайд 23Биосинтез нуклеотидов.
Пуриновые и пиримидиновые нуклеотиды — это те строительные блоки, из
Биосинтез нуклеотидов.
Пуриновые и пиримидиновые нуклеотиды — это те строительные блоки, из

Слайд 24Биосинтез жиров.
Жиры или липиды являются важными компонентами ЦПМ и клеточной стенки
Биосинтез жиров.
Жиры или липиды являются важными компонентами ЦПМ и клеточной стенки

Слайд 25Биосинтез углеводов.
Углеводы представлены в бактериальной клетке в виде моно-, ди- и
Биосинтез углеводов.
Углеводы представлены в бактериальной клетке в виде моно-, ди- и

 Поликультурное образование. Формирование идентичности обучающихся средствами курса ОРКСЭ
Поликультурное образование. Формирование идентичности обучающихся средствами курса ОРКСЭ Стратегический анализ транспортно-пересадочных узлов города Москвы
Стратегический анализ транспортно-пересадочных узлов города Москвы Вернуться из леса (2)
Вернуться из леса (2) Абсолютные и относительные величины
Абсолютные и относительные величины  Компьютерные презентации Мультимедийные технологии
Компьютерные презентации Мультимедийные технологии Mersedes-Vito
Mersedes-Vito Качество и уровень жизни. Бюджет прожиточного минимума. Потребительская корзина
Качество и уровень жизни. Бюджет прожиточного минимума. Потребительская корзина Техническое регулирование в строительстве. Системы стандартизации и добровольной оценки соответствия НОСТРОЙ.
Техническое регулирование в строительстве. Системы стандартизации и добровольной оценки соответствия НОСТРОЙ. Автоматика судовых энергетических установок и вспомогательных механизмов практика
Автоматика судовых энергетических установок и вспомогательных механизмов практика Классификация и оценка рисков
Классификация и оценка рисков Ранние половые связи подростков. Родительское собрание
Ранние половые связи подростков. Родительское собрание Как задавать домашнее задание?
Как задавать домашнее задание? Я0зык
Я0зык Собственные и нарицательные имена существительные
Собственные и нарицательные имена существительные Динамика точки
Динамика точки Модульный урок«Атомы химических элементов. Структура Периодической системы»
Модульный урок«Атомы химических элементов. Структура Периодической системы» B i l d e n d e K u n s t
B i l d e n d e K u n s t Орфографическая минутка
Орфографическая минутка Внутренние воды Южной Америки
Внутренние воды Южной Америки Презентация на тему Патриоты России (4 класс)
Презентация на тему Патриоты России (4 класс) Сказка о возвращенном времени. Кто они, герои картин В.И.Сурикова?
Сказка о возвращенном времени. Кто они, герои картин В.И.Сурикова? Реклама в мобильных приложениях. Коммерческое предложение для СМИ
Реклама в мобильных приложениях. Коммерческое предложение для СМИ Манипуляции в общении и переговорах Изобретательностью и умом побеждать лучше, чем сопротивлением
Манипуляции в общении и переговорах Изобретательностью и умом побеждать лучше, чем сопротивлением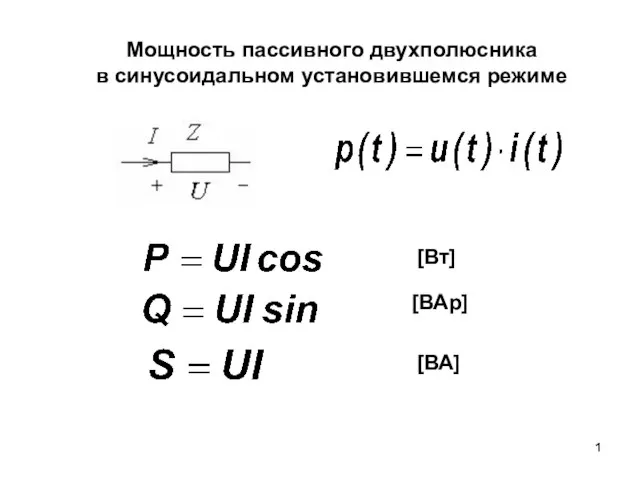 Мощность пассивного двухполюсника в синусоидальном установившемся режиме
Мощность пассивного двухполюсника в синусоидальном установившемся режиме Источники и институты классического средневекового права
Источники и институты классического средневекового права Солнечногорск
Солнечногорск Мы открыты для сотрудничества. АНО Начинающий фермер, Семейная животноводческая ферма
Мы открыты для сотрудничества. АНО Начинающий фермер, Семейная животноводческая ферма Pop-up формат. Временная розничная торговля
Pop-up формат. Временная розничная торговля