- Презентация на тему Днк Дезоксирибонуклеиновая кислота

Содержание
- 2. Дезоксирибонуклеиновая кислота ДНК –биологический полимер, состоящий из двух спирально закрученных цепочек.
- 3. История открытия. В 1869 г. Фридрих Мишер, швейцарский врач биохимик, выделил нуклеиновые кислоты из ядер клеток
- 4. История открытия. Альтман обнаружил ортофосфорную кислоту в составе аминокислот. Именно ее он поначалу называл нуклеиновой кислотой.
- 5. История открытия. В 1912 г. Леви обнаружил, что в состав нуклеиновых кислот входит углевод пентоза. В
- 6. История открытия. Однако, эта правильная точка зрения просуществовала не долго. При гистохимическом анализе кислот было обнаружено,
- 7. История открытия. В 1936 г. советский ученый Белозерский доказал что в проростках конского каштана содержится тимонуклеиновая
- 8. История открытия. 1953 г. американские биохимики Дж. Уотсон и Ф.Крик установили расположение частей молекулы ДНК
- 9. Первичная структура нуклеиновых кислот. Под первичной структурой нуклеиновых кислот понимают порядок, последовательность расположения мононуклеотидов в полинуклеотидной
- 10. Первичная структура нуклеиновых кислот. Установлено, что в образовании межнуклеотидной связи участвуют гидроксильные группы в 3'- и
- 11. Первичная структура нуклеиновых кислот. три варианта схемы нуклеотидной последовательности ДНК:
- 12. Строение ДНК. ДНК - полимер. Мономеры - нуклеотиды. Нуклеотид- химическое соединение остатков трех веществ: Строение нуклеотида
- 13. Конформации компонентов нуклеиновых кислот. Все 5 гетероциклических оснований, входящих в состав НК, имеют плоскую конформацию. Для
- 14. Син- и анти- конформации нуклеозидов В свободных нуклеозидах и нуклеотидах переход от C2`- эндо- к C3`
- 15. Макромолекулярная структура ДНК. В 1953 г. Дж.Уотсон и Ф.Крик предложили модель структуры ДНК. При постоении стуктуры
- 16. Макромолекулярная структура ДНК. - правильная правовинтовая спираль, состоящая из 2 полинуклеатидных цепей, которые закручены друг относительно
- 17. Полиморфизм двойной спирали. Правые спирали образуют 2 семейства: А-семейство (конформация сахара С3`- эндо-) и В-семейство (конформация
- 18. А – семейство ДНК. Розалинда Франклин получила эксперементальные свидетельства существования весьма упорядоченной структуры в ориентированных вытягиванием
- 19. А – семейство ДНК. С3`- эндоконформация сахара приводит к уменьшению расстояния между фосфатными группами и, следовательно,
- 20. А – семейство ДНК. Пары оснований в А-форме образуют с осью спирали угол около 20 градусов
- 21. А – семейство ДНК.
- 22. В – семейство ДНК. Для этого семейства характерно структурное разнообразие. ДНК со случайными последовательностями могут находиться
- 23. Z – форма ДНК. Левоспиральная конформация ДНК. Она была открыта в 1979 г. при исследовании структуры
- 24. Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах. 2 типа взаимодействия между гетероциклическими основаниями нуклеотидных остатков: взаимодействия
- 26. Скачать презентацию
Слайд 2Дезоксирибонуклеиновая кислота
ДНК –биологический полимер, состоящий из двух спирально закрученных цепочек.
Дезоксирибонуклеиновая кислота
ДНК –биологический полимер, состоящий из двух спирально закрученных цепочек.

Слайд 3История открытия.
В 1869 г. Фридрих Мишер, швейцарский врач биохимик, выделил нуклеиновые кислоты
История открытия.
В 1869 г. Фридрих Мишер, швейцарский врач биохимик, выделил нуклеиновые кислоты

Слайд 4История открытия.
Альтман обнаружил ортофосфорную кислоту в составе аминокислот. Именно ее он поначалу
История открытия.
Альтман обнаружил ортофосфорную кислоту в составе аминокислот. Именно ее он поначалу

Слайд 5История открытия.
В 1912 г. Леви обнаружил, что в состав нуклеиновых кислот входит
История открытия.
В 1912 г. Леви обнаружил, что в состав нуклеиновых кислот входит

Слайд 6История открытия.
Однако, эта правильная точка зрения просуществовала не долго. При гистохимическом анализе
История открытия.
Однако, эта правильная точка зрения просуществовала не долго. При гистохимическом анализе

Слайд 7История открытия.
В 1936 г. советский ученый Белозерский доказал что в проростках конского
История открытия.
В 1936 г. советский ученый Белозерский доказал что в проростках конского

Слайд 8История открытия.
1953 г. американские биохимики Дж. Уотсон и Ф.Крик установили расположение частей
История открытия.
1953 г. американские биохимики Дж. Уотсон и Ф.Крик установили расположение частей

Слайд 9Первичная структура нуклеиновых кислот.
Под первичной структурой нуклеиновых кислот понимают порядок, последовательность расположения
Первичная структура нуклеиновых кислот.
Под первичной структурой нуклеиновых кислот понимают порядок, последовательность расположения
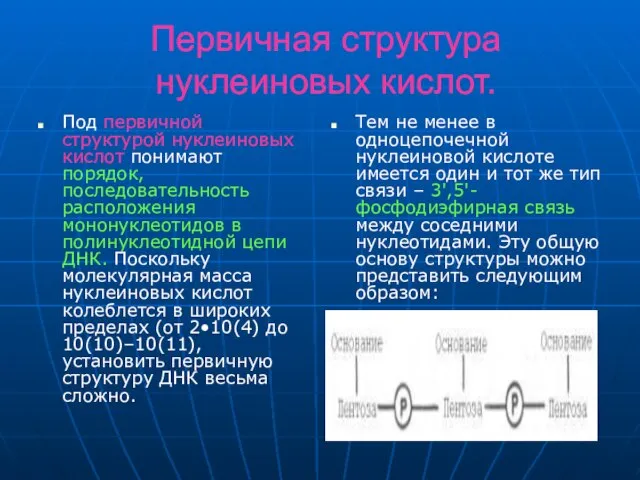
Слайд 10Первичная структура нуклеиновых кислот.
Установлено, что в образовании межнуклеотидной связи участвуют гидроксильные группы
Первичная структура нуклеиновых кислот.
Установлено, что в образовании межнуклеотидной связи участвуют гидроксильные группы

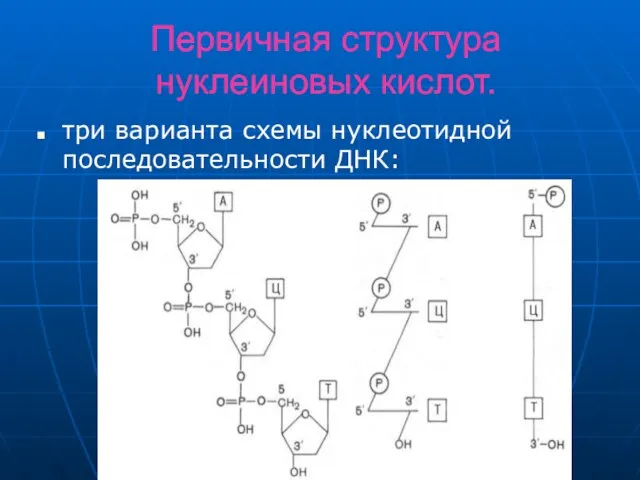
Слайд 11Первичная структура нуклеиновых кислот.
три варианта схемы нуклеотидной последовательности ДНК:
Первичная структура нуклеиновых кислот.
три варианта схемы нуклеотидной последовательности ДНК:
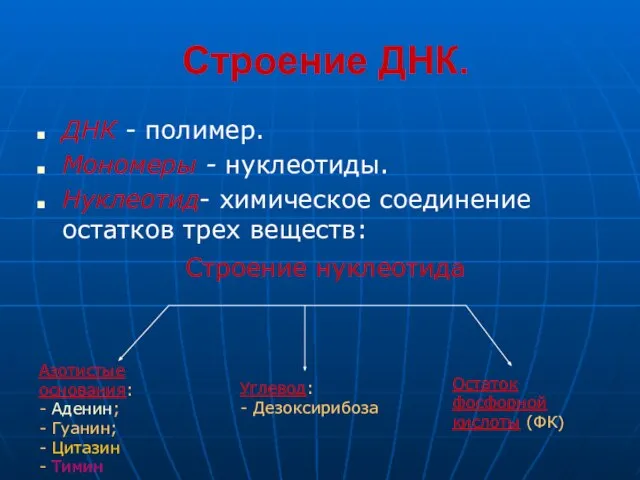
Слайд 12Строение ДНК.
ДНК - полимер.
Мономеры - нуклеотиды.
Нуклеотид- химическое соединение остатков трех веществ:
Строение нуклеотида
Азотистые
основания:
-
Строение ДНК.
ДНК - полимер.
Мономеры - нуклеотиды.
Нуклеотид- химическое соединение остатков трех веществ:
Строение нуклеотида
Азотистые
основания:
-
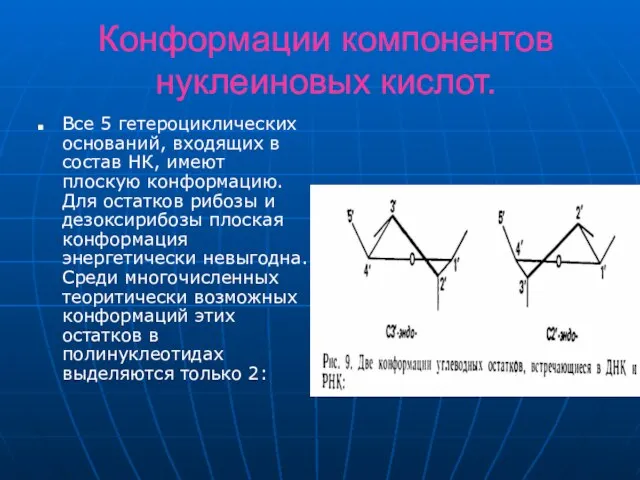
Слайд 13Конформации компонентов нуклеиновых кислот.
Все 5 гетероциклических оснований, входящих в состав НК, имеют
Конформации компонентов нуклеиновых кислот.
Все 5 гетероциклических оснований, входящих в состав НК, имеют
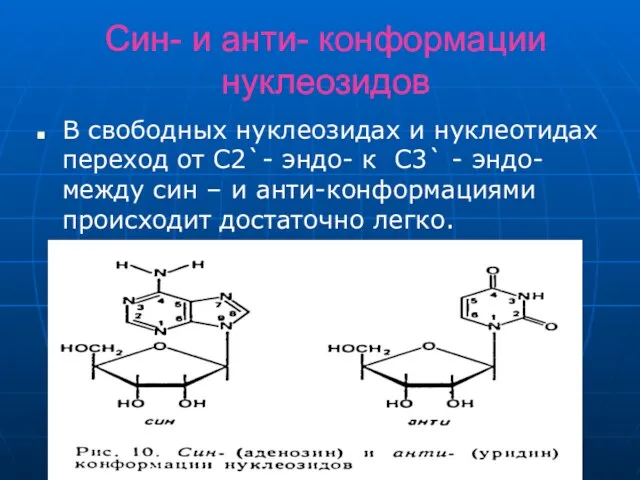
Слайд 14Син- и анти- конформации нуклеозидов
В свободных нуклеозидах и нуклеотидах переход от C2`-
Син- и анти- конформации нуклеозидов
В свободных нуклеозидах и нуклеотидах переход от C2`-
Слайд 15Макромолекулярная структура ДНК.
В 1953 г. Дж.Уотсон и Ф.Крик предложили модель структуры ДНК.
Макромолекулярная структура ДНК.
В 1953 г. Дж.Уотсон и Ф.Крик предложили модель структуры ДНК.

Слайд 16Макромолекулярная структура ДНК.
- правильная правовинтовая спираль, состоящая из 2 полинуклеатидных цепей, которые
Макромолекулярная структура ДНК.
- правильная правовинтовая спираль, состоящая из 2 полинуклеатидных цепей, которые

Слайд 17Полиморфизм двойной спирали.
Правые спирали образуют 2 семейства: А-семейство (конформация сахара С3`- эндо-)
Полиморфизм двойной спирали.
Правые спирали образуют 2 семейства: А-семейство (конформация сахара С3`- эндо-)

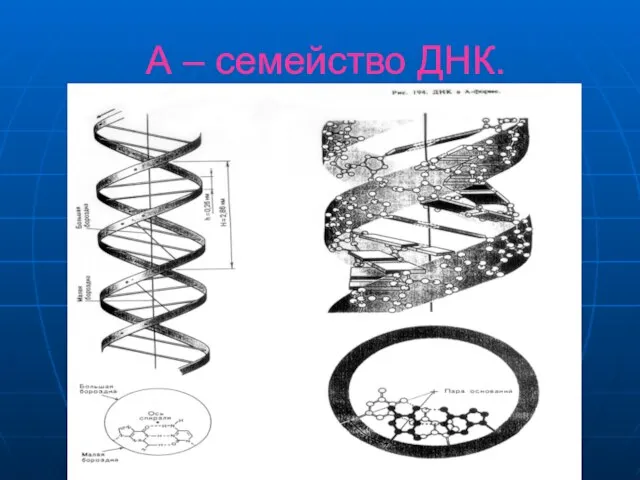
Слайд 18А – семейство ДНК.
Розалинда Франклин получила эксперементальные свидетельства существования весьма упорядоченной структуры
А – семейство ДНК.
Розалинда Франклин получила эксперементальные свидетельства существования весьма упорядоченной структуры

Слайд 19А – семейство ДНК.
С3`- эндоконформация сахара приводит к уменьшению расстояния между фосфатными
А – семейство ДНК.
С3`- эндоконформация сахара приводит к уменьшению расстояния между фосфатными

Слайд 20А – семейство ДНК.
Пары оснований в А-форме образуют с осью спирали угол
А – семейство ДНК.
Пары оснований в А-форме образуют с осью спирали угол

Слайд 21А – семейство ДНК.
А – семейство ДНК.
Слайд 22В – семейство ДНК.
Для этого семейства характерно структурное разнообразие. ДНК со случайными
В – семейство ДНК.
Для этого семейства характерно структурное разнообразие. ДНК со случайными

Слайд 23Z – форма ДНК.
Левоспиральная конформация ДНК. Она была открыта в 1979 г.
Z – форма ДНК.
Левоспиральная конформация ДНК. Она была открыта в 1979 г.

Слайд 24Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах.
2 типа взаимодействия между гетероциклическими основаниями
Взаимодействия между гетероциклическими основаниями в нуклеиновых кислотах.
2 типа взаимодействия между гетероциклическими основаниями

 Презентация на тему Новые педагогические технологии
Презентация на тему Новые педагогические технологии Обязанности таможенной финансовой службы
Обязанности таможенной финансовой службы Мы за здоровый образ жизни
Мы за здоровый образ жизни Озера авт. Дудаева Л.В.
Озера авт. Дудаева Л.В. Моя история. Вера в успех
Моя история. Вера в успех Екологічні рішення від YASNO. Проектування та встановлення сонячної електростанції
Екологічні рішення від YASNO. Проектування та встановлення сонячної електростанції ДОЛ Пионер - лагерь нового поколения!
ДОЛ Пионер - лагерь нового поколения! Текстовые документы и технологии их создания
Текстовые документы и технологии их создания Портрет - Поиск
Портрет - Поиск Таможенное оформление поставок запчастей
Таможенное оформление поставок запчастей История экономической мысли 2
История экономической мысли 2 Храни меня, мой талисман
Храни меня, мой талисман Свинец, плюсы и минусы
Свинец, плюсы и минусы Сохранение психологического и физического здоровья педагога
Сохранение психологического и физического здоровья педагога Золотая осень
Золотая осень Последствия для потребителей решений Правительства РФ в части оплаты электрической энергии и мощности
Последствия для потребителей решений Правительства РФ в части оплаты электрической энергии и мощности Виды рекламы
Виды рекламы Презентация по основам безопасности жизнедеятельности на тему _Социальная роль женщины в современном обществе_
Презентация по основам безопасности жизнедеятельности на тему _Социальная роль женщины в современном обществе_ Идеал красоты в разные эпохи
Идеал красоты в разные эпохи Ветвления
Ветвления Перевод чисел из одной системы счисления в другую
Перевод чисел из одной системы счисления в другую Презентация на тему Самоуправление
Презентация на тему Самоуправление  Налоговая система унитарных государств
Налоговая система унитарных государств Особенности ГИА-11 в 2020 году
Особенности ГИА-11 в 2020 году ТВОРЧЕСКИЙ ОТЧЕТ
ТВОРЧЕСКИЙ ОТЧЕТ Sport in Russia
Sport in Russia Моторайд.рус - сообщество любителей езды на мотоциклах
Моторайд.рус - сообщество любителей езды на мотоциклах ребования к проектно-сметной документации в рамках кредитного продукта Ипотечный кредит с льготной процентной ставкой
ребования к проектно-сметной документации в рамках кредитного продукта Ипотечный кредит с льготной процентной ставкой