- Процессинг антигенов

Содержание
- 2. T клетки не распознают нативные антигены Y Y Y Y Y Y Пролиферация и продукция антител
- 3. Антигены должны пройти процессинг, чтобы их распознали Т лимфоциты T клеточный ответ Нет Т-ответа Нет Т-
- 4. Для взаимодействия T клеток с макрофагами необходим катаболизм антигена Listeria Tклетки не связываются с АПК до
- 5. Только метаболически активные клетки могут осуществлять процессинг АГ Фиксация формальдегидом Инкубация с Listeria 60мин добавление Listeria
- 6. Достаточен ли только экзогенный процессинг? У большинства типов клеток нет такой развитой лизосомальной системы, как у
- 7. Y Места размножения патогенов определяют тип процессинга антигенов Цитозольный компартмент Процессинг эндогенных АГ (вирусные АГ) Везикулярный
- 8. Y Элиминируются с помощью: Киллинга инфицированных клеток с помощью CTL-цитотоксических Т лимфоитов Элиминируются с помощью: Антител
- 9. Стадии процессинга экзогенных и эндогенных АГ Захват Вовлечение нативных АГ и патогенов во внутриклеточные механизмы деградации
- 10. Изменения клеточной оболочки А - при экзоцитозе; Б - при эндоцитозе; В - при мерокринном типе
- 11. Эндоцитоз
- 12. АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
- 13. Y пиноцитоз фагоцитоз Ig –рецепторы на мембране связывают АГ Захват экзогенных антигенов: механизмы Рецепторы к компонентам
- 14. Протеазы образуют примерно 24- аминокислотные пептиды из АГ Экзогенный путь Белковые АГ в эндосоме катепсины B,
- 15. Протеазы образуют из АГ пептиды длиной ~24 аминокислот Лекарства, повышающие pH в эндосомах, ингибируют процессинг АГ
- 16. В молекулах MHC есть сайты связвания с пептидами, которые в незрелых молекулах МНС – гибкие, подвижные
- 17. Необходимо предотвратить связывание с незрелыми молекулами MHC синтезируемых ауто - белков Инвариантная цепь стабилизирует молекулы MHC
- 18. Пептид инвариантной цепи блокирует сайт связывания в молекуле MHC Этот пептид называется CLass II associated Invariant
- 19. CLIP –пептид, связанный с инвариантной цепью молекулы МНС II класса (αβinv)3 комплексы двигаются к эндосоме ин-
- 20. Удаление CLIP ? Каким образом АГ- пептид связывается с сайтом связывания? Конкуренция между большим числом пептидов
- 21. HLA-DM HLA-DR HLA-DM помогает в удалении CLIP HLA-DM: кристаллизируется без пептида в полости –пространство в полости
- 22. HLA-DM HLA-DR Этот карман –загружен – он недоступен для АГ- пептида Множество карманов в полости свободны
- 23. В MIIC компартменте происходит сортировка комплексов MHC – антигенный пептид либо для поверхностной экспрессии либо для
- 24. Захват Антиген / патоген уже находится внутри клетки Деградация протеолитическая деградация в цитоплазме Формирование комплекса антиген-MHC
- 25. Разрушение в протеосоме Компоненты протеосом включают MECL-1, LMP2, LMP7 Эти компоненты индуцируются IFN-γ и становятся компонентами
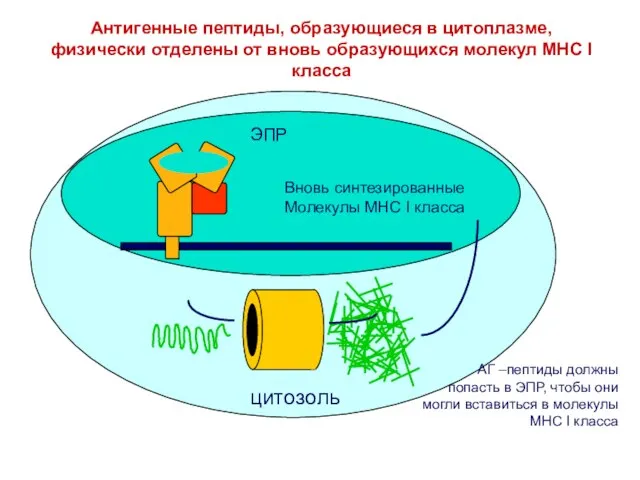
- 26. ЭПР цитозоль Антигенные пептиды, образующиеся в цитоплазме, физически отделены от вновь образующихся молекул MHC I класса
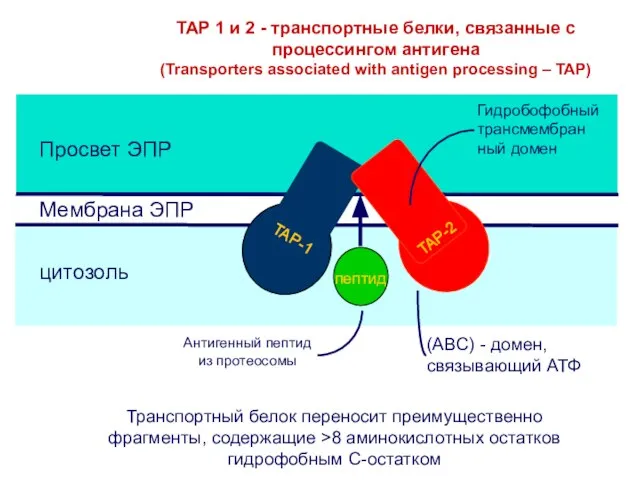
- 27. TAP 1 и 2 - транспортные белки, связанные с процессингом антигена (Transporters associated with antigen processing
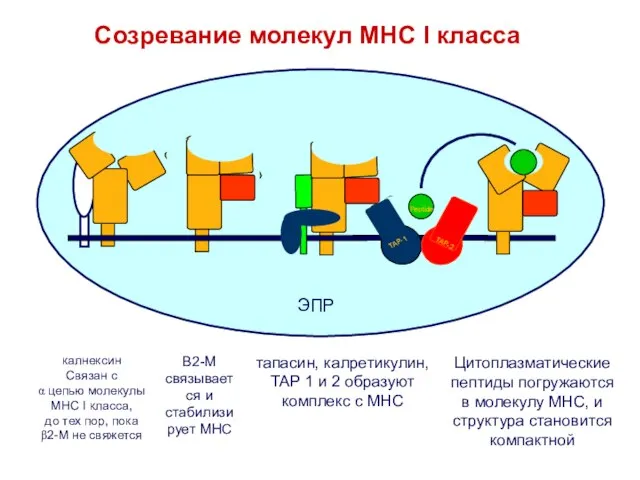
- 28. калнексин Связан с α цепью молекулы МНС I класса, до тех пор, пока β2-M не свяжется
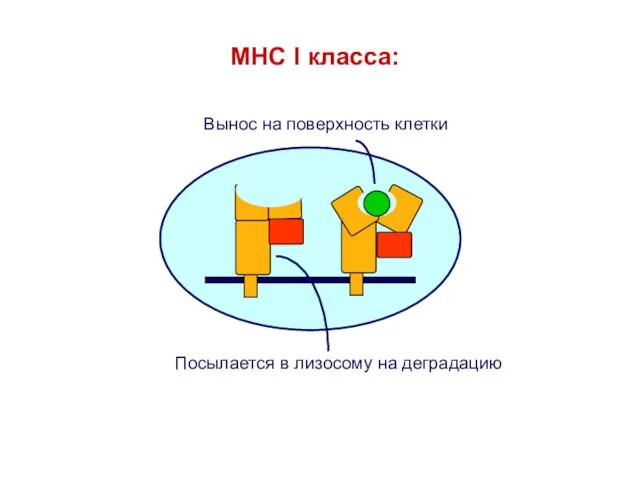
- 29. MHC I класса:
- 30. Нарушение иммунного ответа путем влияния на процессинг эндогенных антигенов
- 31. Нарушение иммунного ответа путем влияния на процессинг эндогенных антигенов
- 32. T и B клетки распознают антигены по - разному Перед тем, как Т–лимфоцит распознает АГ, АГ
- 33. В любой ядерной клетке антигены после фрагментирования на олигопептиды транспортируются в ЭПР, где они связываются с
- 34. Субъединицы MHC-II молекул сразу после образования соединяются в ЭПР с инвариантной цепью (IC), препятствующей ассоциации MHC-II
- 35. Костимуляция Процесс презентации антигенов сопровождается костимуляцией в результате взаимодействия молекул и их лигандов, экспрессирующихся на поверхностных
- 37. Скачать презентацию
Слайд 2T клетки не распознают нативные антигены
Y
Y
Y
Y
Y
Y
Пролиферация и продукция антител
Нет пролиферации,нет
освобождения цитокинов
Перекрестное
T клетки не распознают нативные антигены
Y
Y
Y
Y
Y
Y
Пролиферация и продукция антител
Нет пролиферации,нет
освобождения цитокинов
Перекрестное
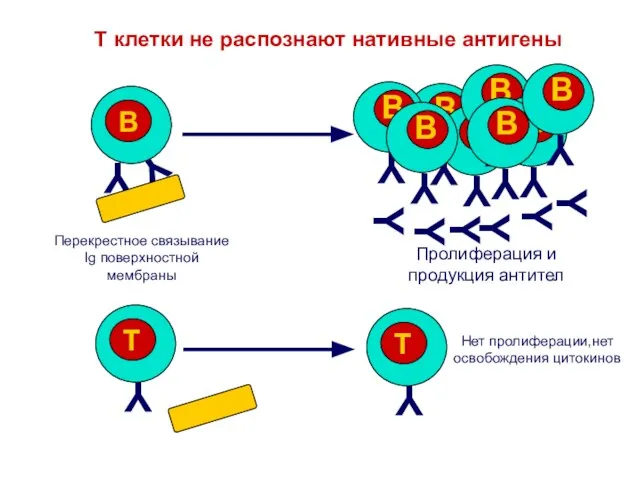
Слайд 3Антигены должны пройти процессинг,
чтобы их распознали Т лимфоциты
T клеточный
ответ
Нет
Антигены должны пройти процессинг,
чтобы их распознали Т лимфоциты
T клеточный
ответ
Нет
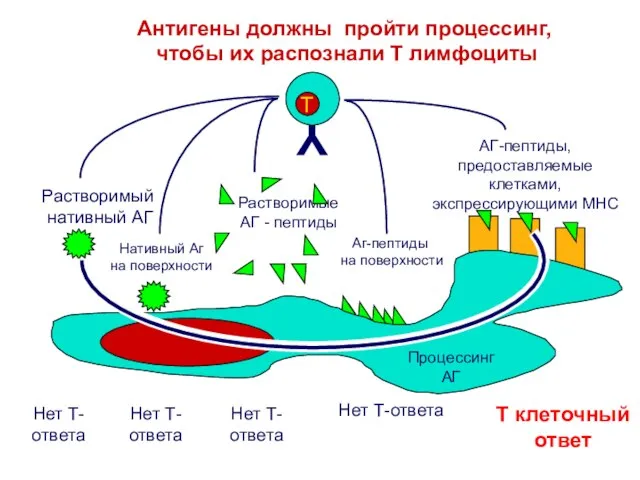
Слайд 4Для взаимодействия T клеток с макрофагами
необходим катаболизм антигена
Listeria
Tклетки не связываются с АПК
Для взаимодействия T клеток с макрофагами
необходим катаболизм антигена
Listeria
Tклетки не связываются с АПК
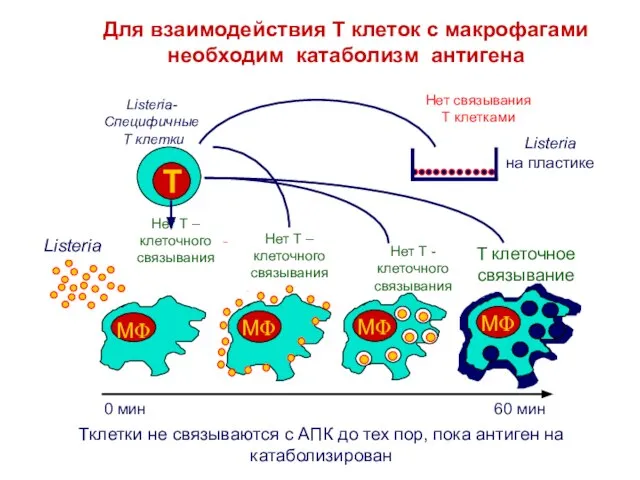
Слайд 5Только метаболически активные клетки могут осуществлять процессинг АГ
Фиксация формальдегидом
Инкубация с
Listeria 60мин
добавление Listeria
специфических
Только метаболически активные клетки могут осуществлять процессинг АГ
Фиксация формальдегидом
Инкубация с
Listeria 60мин
добавление Listeria
специфических
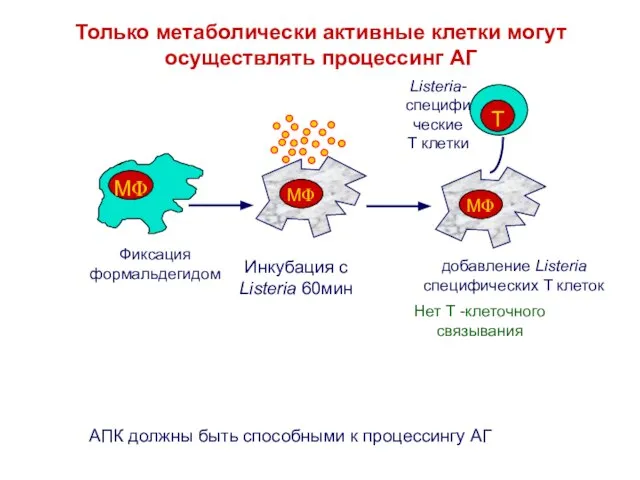
Слайд 6Достаточен ли только экзогенный процессинг?
У большинства типов клеток нет такой развитой лизосомальной
Достаточен ли только экзогенный процессинг?
У большинства типов клеток нет такой развитой лизосомальной

Слайд 7Y
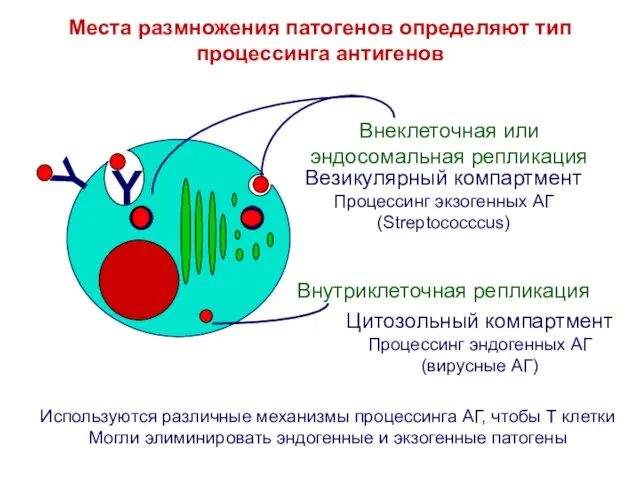
Места размножения патогенов определяют тип процессинга антигенов
Цитозольный компартмент
Процессинг эндогенных АГ
(вирусные АГ)
Везикулярный компартмент
Процессинг
Y
Места размножения патогенов определяют тип процессинга антигенов
Цитозольный компартмент
Процессинг эндогенных АГ
(вирусные АГ)
Везикулярный компартмент
Процессинг
Слайд 8Y
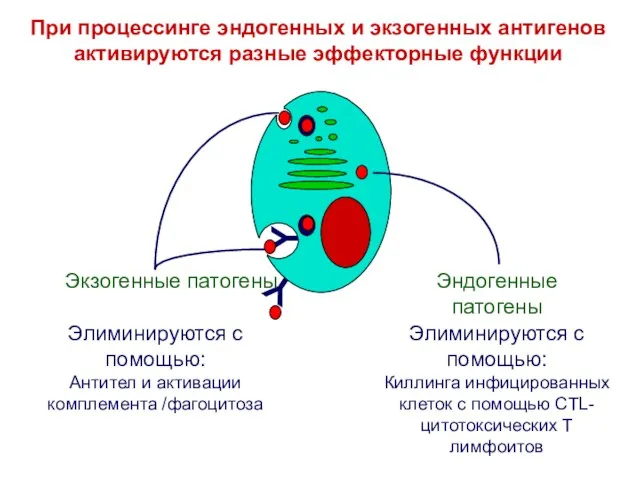
Элиминируются с помощью:
Киллинга инфицированных клеток с помощью CTL-цитотоксических Т лимфоитов
Элиминируются с помощью:
Антител
Y
Элиминируются с помощью:
Киллинга инфицированных клеток с помощью CTL-цитотоксических Т лимфоитов
Элиминируются с помощью:
Антител
Слайд 9Стадии процессинга экзогенных
и эндогенных АГ
Захват
Вовлечение нативных АГ и патогенов во
Стадии процессинга экзогенных
и эндогенных АГ
Захват
Вовлечение нативных АГ и патогенов во

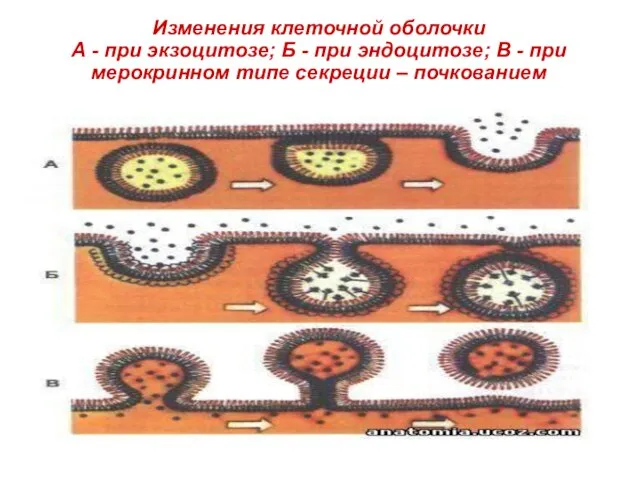
Слайд 10Изменения клеточной оболочки
А - при экзоцитозе; Б - при эндоцитозе; В
Изменения клеточной оболочки А - при экзоцитозе; Б - при эндоцитозе; В
Слайд 11Эндоцитоз
Эндоцитоз

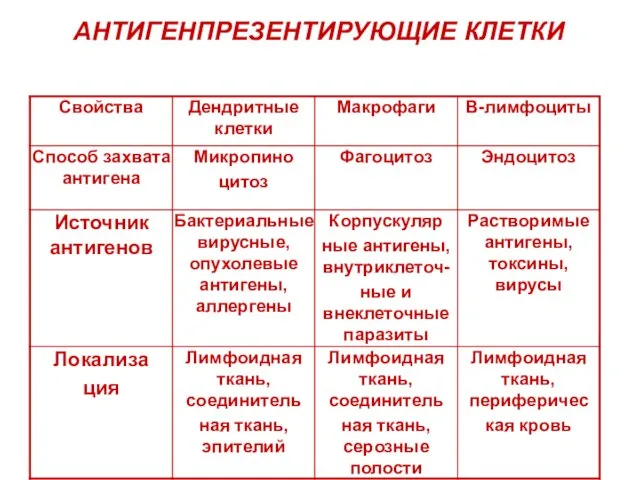
Слайд 12АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
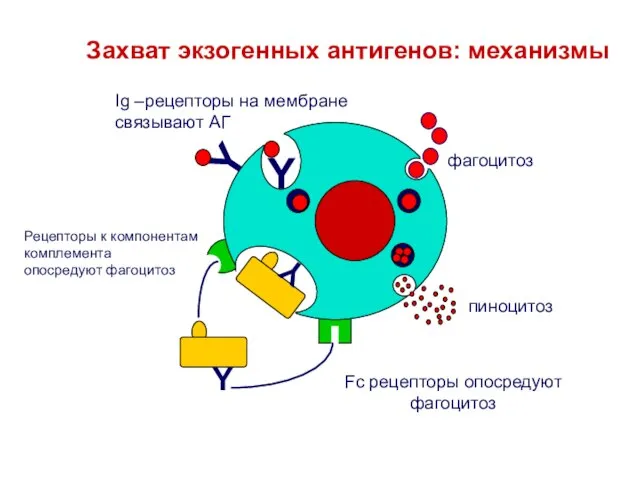
Слайд 13Y
пиноцитоз
фагоцитоз
Ig –рецепторы на мембране
связывают АГ
Захват экзогенных антигенов: механизмы
Рецепторы к компонентам
комплемента
опосредуют фагоцитоз
Fc
Y
пиноцитоз
фагоцитоз
Ig –рецепторы на мембране
связывают АГ
Захват экзогенных антигенов: механизмы
Рецепторы к компонентам
комплемента
опосредуют фагоцитоз
Fc
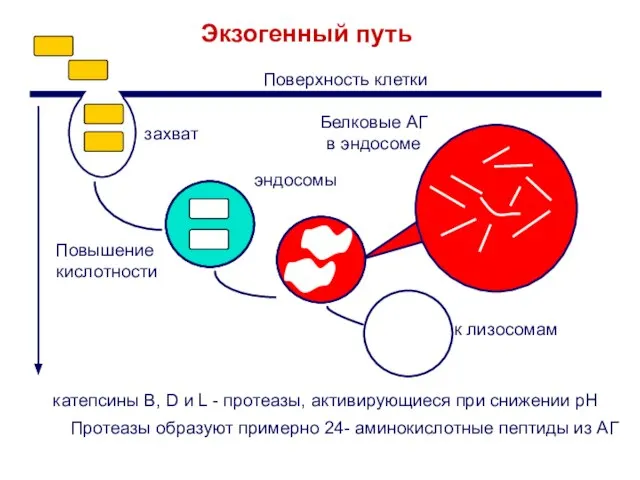
Слайд 14Протеазы образуют примерно 24- аминокислотные пептиды из АГ
Экзогенный путь
Белковые АГ
в эндосоме
катепсины B,
Протеазы образуют примерно 24- аминокислотные пептиды из АГ
Экзогенный путь
Белковые АГ
в эндосоме
катепсины B,
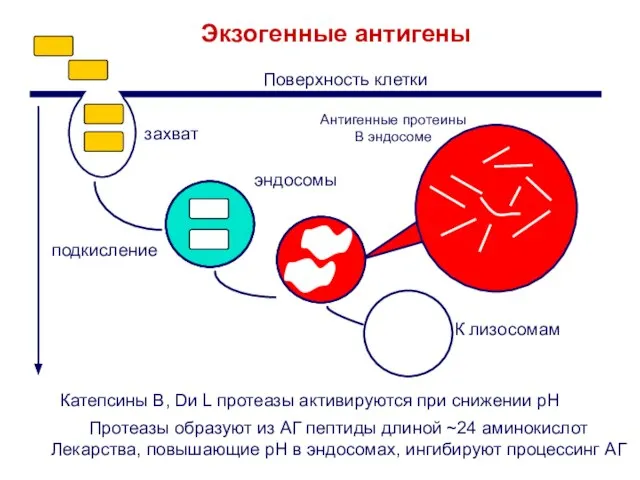
Слайд 15Протеазы образуют из АГ пептиды длиной ~24 аминокислот
Лекарства, повышающие pH в эндосомах,
Протеазы образуют из АГ пептиды длиной ~24 аминокислот
Лекарства, повышающие pH в эндосомах,
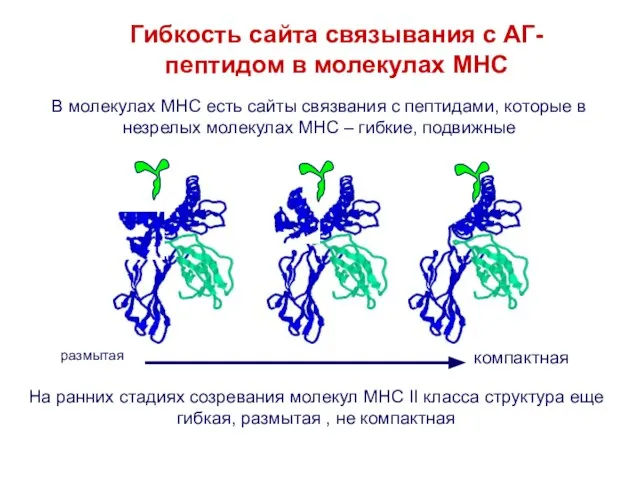
Слайд 16В молекулах MHC есть сайты связвания с пептидами, которые в незрелых молекулах
В молекулах MHC есть сайты связвания с пептидами, которые в незрелых молекулах
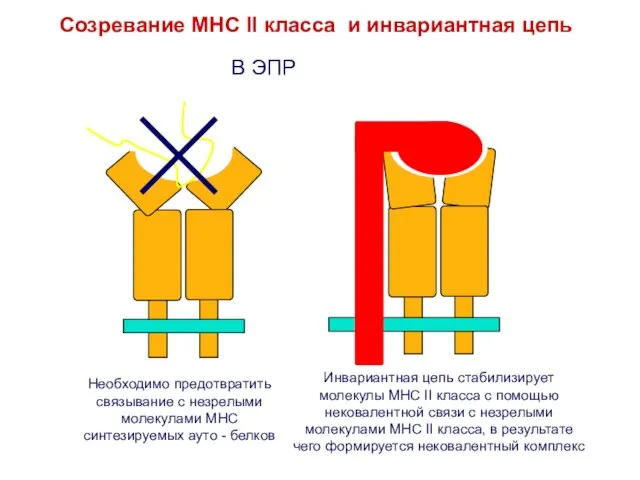
Слайд 17Необходимо предотвратить связывание с незрелыми молекулами MHC синтезируемых ауто - белков
Инвариантная цепь
Необходимо предотвратить связывание с незрелыми молекулами MHC синтезируемых ауто - белков
Инвариантная цепь
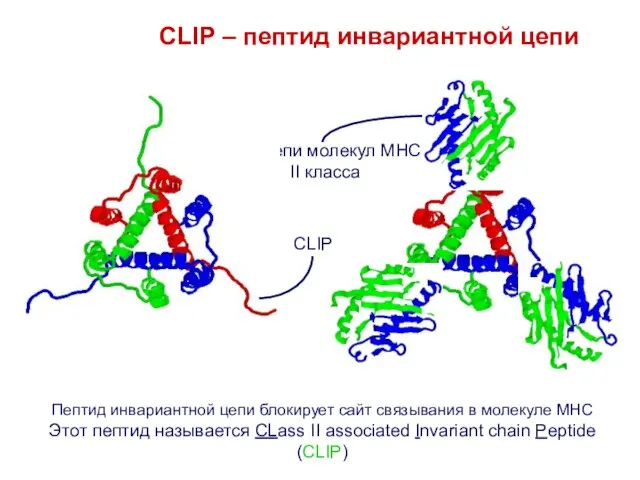
Слайд 18Пептид инвариантной цепи блокирует сайт связывания в молекуле MHC
Этот пептид называется CLass
Пептид инвариантной цепи блокирует сайт связывания в молекуле MHC
Этот пептид называется CLass
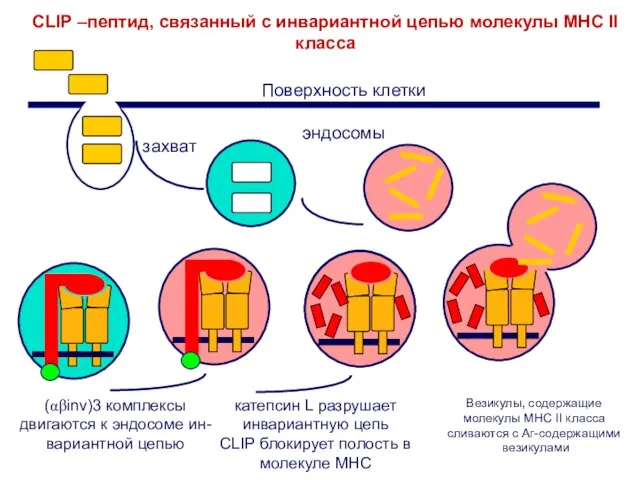
Слайд 19CLIP –пептид, связанный с инвариантной цепью молекулы МНС II класса
(αβinv)3 комплексы
двигаются к
CLIP –пептид, связанный с инвариантной цепью молекулы МНС II класса
(αβinv)3 комплексы
двигаются к
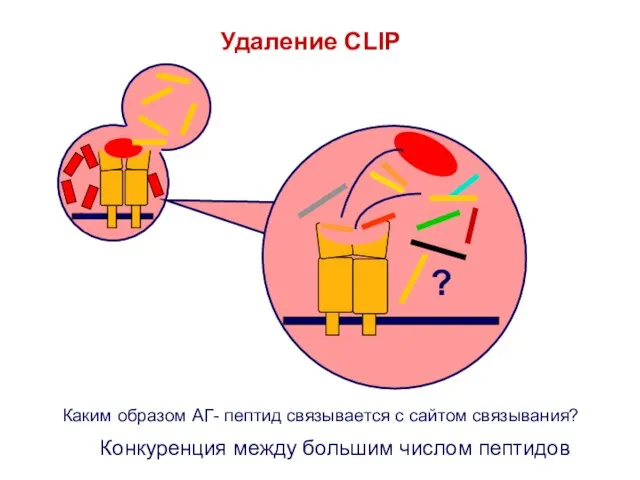
Слайд 20Удаление CLIP
?
Каким образом АГ- пептид связывается с сайтом связывания?
Конкуренция между большим числом
Удаление CLIP
?
Каким образом АГ- пептид связывается с сайтом связывания?
Конкуренция между большим числом
Слайд 21HLA-DM
HLA-DR
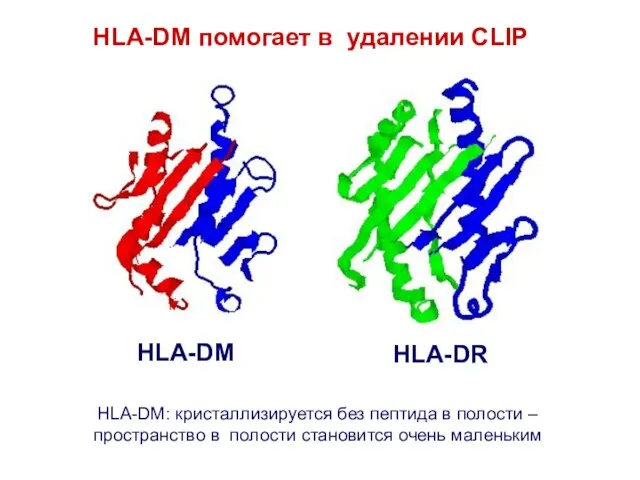
HLA-DM помогает в удалении CLIP
HLA-DM: кристаллизируется без пептида в полости –пространство в
HLA-DM
HLA-DR
HLA-DM помогает в удалении CLIP
HLA-DM: кристаллизируется без пептида в полости –пространство в
Слайд 22HLA-DM
HLA-DR
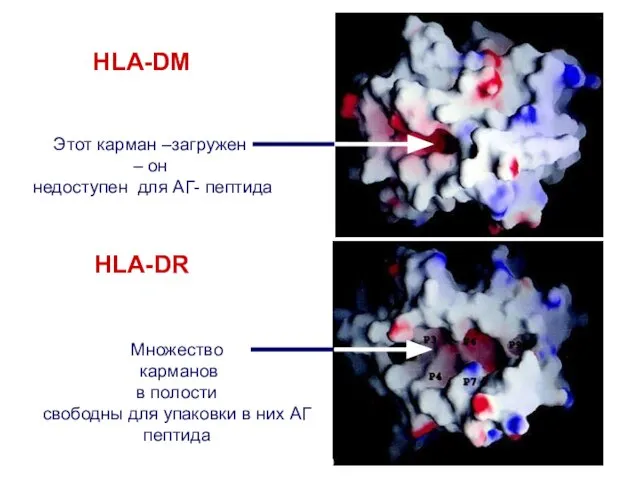
Этот карман –загружен
– он
недоступен для АГ- пептида
Множество
карманов
в полости
свободны
HLA-DM
HLA-DR
Этот карман –загружен
– он
недоступен для АГ- пептида
Множество
карманов
в полости
свободны
Слайд 23В MIIC компартменте происходит сортировка комплексов MHC – антигенный пептид
либо для поверхностной
В MIIC компартменте происходит сортировка комплексов MHC – антигенный пептид
либо для поверхностной
Слайд 24Захват
Антиген / патоген уже находится внутри клетки
Деградация
протеолитическая деградация в цитоплазме
Формирование комплекса антиген-MHC
Упаковка
Захват
Антиген / патоген уже находится внутри клетки
Деградация
протеолитическая деградация в цитоплазме
Формирование комплекса антиген-MHC
Упаковка

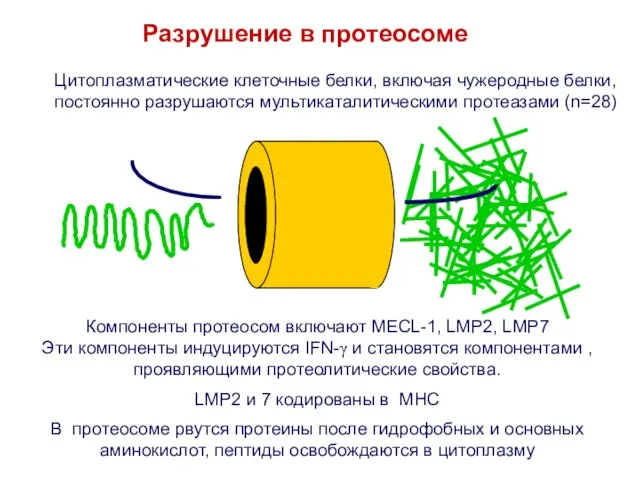
Слайд 25Разрушение в протеосоме
Компоненты протеосом включают MECL-1, LMP2, LMP7
Эти компоненты индуцируются IFN-γ и
Разрушение в протеосоме
Компоненты протеосом включают MECL-1, LMP2, LMP7
Эти компоненты индуцируются IFN-γ и
Слайд 26ЭПР
цитозоль
Антигенные пептиды, образующиеся в цитоплазме, физически отделены от вновь образующихся молекул MHC
ЭПР
цитозоль
Антигенные пептиды, образующиеся в цитоплазме, физически отделены от вновь образующихся молекул MHC
Слайд 27TAP 1 и 2 - транспортные белки, связанные с процессингом антигена
(Transporters
TAP 1 и 2 - транспортные белки, связанные с процессингом антигена
(Transporters
Слайд 28калнексин
Связан с
α цепью молекулы
МНС I класса,
до тех пор, пока
β2-M не свяжется
B2-M
калнексин
Связан с
α цепью молекулы
МНС I класса,
до тех пор, пока
β2-M не свяжется
B2-M
Слайд 29MHC I класса:
MHC I класса:
Слайд 30Нарушение иммунного ответа путем влияния на процессинг эндогенных антигенов
Нарушение иммунного ответа путем влияния на процессинг эндогенных антигенов
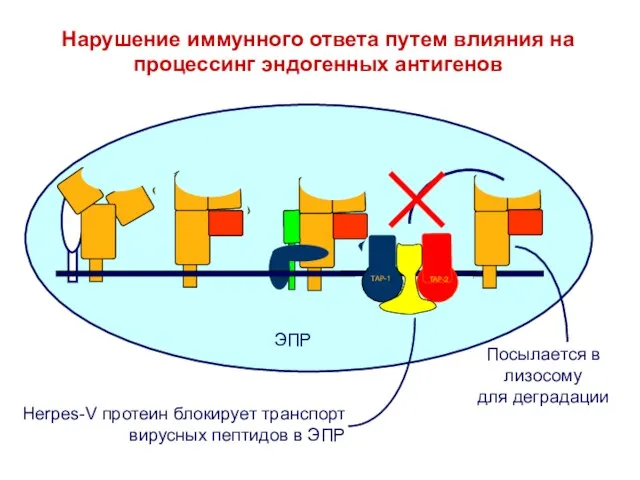
Слайд 31Нарушение иммунного ответа путем
влияния на процессинг эндогенных антигенов
Нарушение иммунного ответа путем
влияния на процессинг эндогенных антигенов
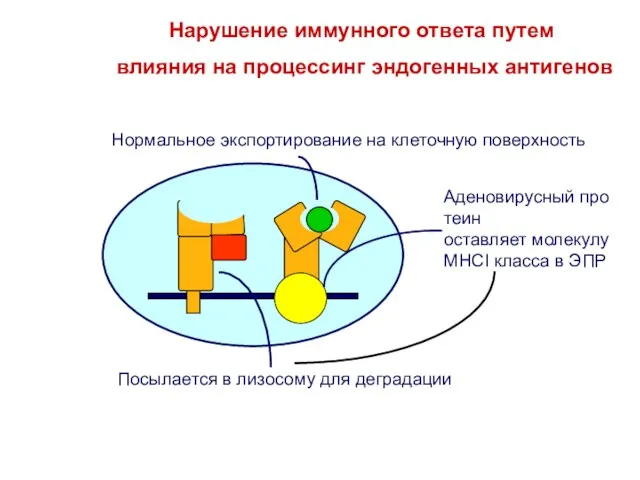
Слайд 32T и B клетки распознают антигены по - разному
Перед тем, как Т–лимфоцит
T и B клетки распознают антигены по - разному
Перед тем, как Т–лимфоцит

Слайд 33В любой ядерной клетке антигены после фрагментирования на олигопептиды транспортируются в ЭПР,
В любой ядерной клетке антигены после фрагментирования на олигопептиды транспортируются в ЭПР,
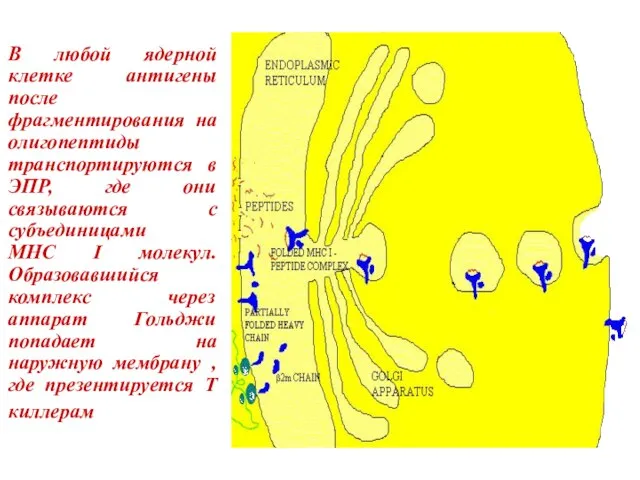
Слайд 34Субъединицы MHC-II молекул сразу после образования соединяются в ЭПР с инвариантной цепью
Субъединицы MHC-II молекул сразу после образования соединяются в ЭПР с инвариантной цепью
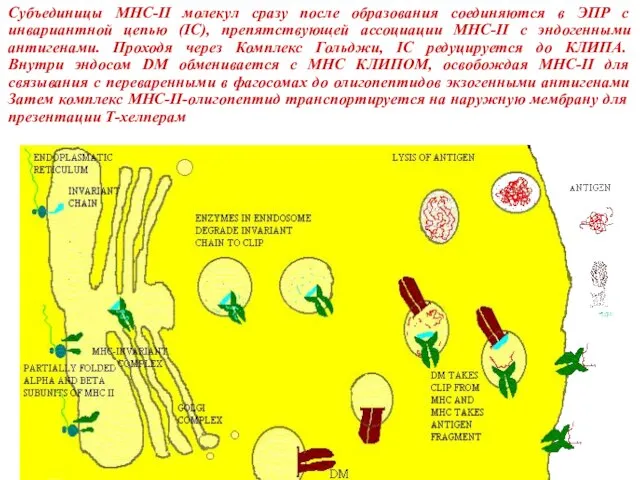
Слайд 35Костимуляция
Процесс презентации антигенов сопровождается костимуляцией в результате взаимодействия молекул и их лигандов,
Костимуляция
Процесс презентации антигенов сопровождается костимуляцией в результате взаимодействия молекул и их лигандов,

 Петр Михайлович Врангель
Петр Михайлович Врангель Витраж. Тематика витражей
Витраж. Тематика витражей МегаФон – Генеральный партнер XI Паралимпийских зимних игр 2014 2 МегаФон получил официальный статус Генерального партнера XI Парали
МегаФон – Генеральный партнер XI Паралимпийских зимних игр 2014 2 МегаФон получил официальный статус Генерального партнера XI Парали Карнавал Марди Гра в Новом Орлеане
Карнавал Марди Гра в Новом Орлеане Система дистанционного обучения Moodle. Работа с интерфейсом системы - общие возможности
Система дистанционного обучения Moodle. Работа с интерфейсом системы - общие возможности Церемония награждения конкурса чтецов
Церемония награждения конкурса чтецов Динамика количественных показателей муниципальной системы образования
Динамика количественных показателей муниципальной системы образования 20140216_dolgosrochnyy_proekt_slyuzova
20140216_dolgosrochnyy_proekt_slyuzova Проектная деятельность как фактор формирования познавательной деятельности учащихся
Проектная деятельность как фактор формирования познавательной деятельности учащихся Язык не может быть плохим или хорошим... Ведь язык - это только зеркало. То самое зеркало, на которое глупо пенять. Се
Язык не может быть плохим или хорошим... Ведь язык - это только зеркало. То самое зеркало, на которое глупо пенять. Се Презентація фокус-групового дослідження “Молодіжна політика в Донецькій області” Компонент: «Молодіжна політика і НГО»
Презентація фокус-групового дослідження “Молодіжна політика в Донецькій області” Компонент: «Молодіжна політика і НГО» Нравственные идеалы. Музыкальное произведение А. П. Бородина
Нравственные идеалы. Музыкальное произведение А. П. Бородина Презентация на тему Международный День Матери
Презентация на тему Международный День Матери Влияние рок-музыки на детей
Влияние рок-музыки на детей Сложение и вычитание трёхзначных чисел в столбик
Сложение и вычитание трёхзначных чисел в столбик Презентация на тему Социально-личностное развитие в ДОУ
Презентация на тему Социально-личностное развитие в ДОУ Практические рекомендации для учителей и родителей
Практические рекомендации для учителей и родителей Конституционное право России. Основы теории конституционализма. Тема № 3
Конституционное право России. Основы теории конституционализма. Тема № 3 Новая система оформления заказов в Орифлейм
Новая система оформления заказов в Орифлейм Презентация на тему Эфиры и жиры
Презентация на тему Эфиры и жиры Present Perfect
Present Perfect  Внутреннее строение насекомых
Внутреннее строение насекомых Механизмы психологической защиты личности.НИГИЛИЗМ и КАТАРСИС
Механизмы психологической защиты личности.НИГИЛИЗМ и КАТАРСИС Britain is my native country
Britain is my native country Невидимые нити в осеннем лесу
Невидимые нити в осеннем лесу Гласный звук [у], буквы У у, Юю
Гласный звук [у], буквы У у, Юю Интернет мощный маркетинговый инструмент в работе салона красоты.
Интернет мощный маркетинговый инструмент в работе салона красоты. Презентация на тему на почте At the post office
Презентация на тему на почте At the post office