- Системы трансдукции сигнала при ответе на феромоны и другие соединения

Содержание
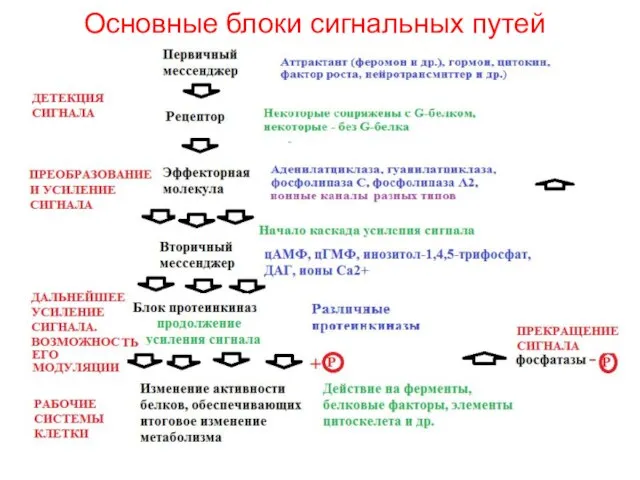
- 2. Основные блоки сигнальных путей
- 3. Первичные мессенджеры - природные экстраклеточные лиганды, которые взаимодействует с рецепторами и активируют их (феромоны и другие
- 4. Адреналиновый каскад. Аденилатциклазная система.
- 5. Для активации 1 молекулы ПКА необходимо 4 молекулы цАМФ
- 6. УСИЛЕНИЕ СИГНАЛА В желтых рамках – нижние границы реального увеличения числа молекул на каждой ступени каскада
- 7. Как остановить сигнал? 1) Дефосфорилировать (фосфатазы) 2) Устранить цАМФ (фосфодиэстераза)
- 8. Инозит(ол) и инозитолфосфатный путь регуляции метаболизма Инозит (биос I, раньше – вит.В8) – водорастворимый Мио-инозит Инозитол-1,4,5-трифосфат
- 9. Недостаточность инозита, источники У человека недостаточность - очень редко. У животных – плешивость, нарушения НС и
- 10. Действие фосфолипаз на глицерофосфолипиды
- 11. Инозитолфосфатный путь регуляции метаболизма по: Зинченко В.П., Долгачева Л.П. Внутриклеточная сигнализация. Пущино, 2003
- 12. Как остановить сигнал? 1) Дефосфорилировать 2) Откачать Са2+ наружу или обратно в ЭПР (Са2+-АТФаза плазматической мембраны,
- 13. Фосфоинозитидный цикл
- 14. Нобелевская премия 2004г. - за исследования «обонятельных ре-цепторов и организации системы органов обоняния» Ричард Аксел Линда
- 15. Ольфакторная (обонятельная) система человека Человек различает в среднем около 10 000 запахов Гены, отвечающие за обоняние,
- 16. Большинство наземных млекопитающих имеют в полости носа, кроме основного органа обоняния, ещё вомероназальный, или ЯКОБСОНОВ ОРГАН
- 17. Ольфакторные системы у млекопитающих У млекопитающих каждый ольфакторный нейрон экспрессирует ген единственного OR-белка Нужны ОR-белки (рецепторы
- 18. Потенциал покоя клетки отрицателен (обычно в пределах от –40 до –60 мВ). Под воздействием стимула он
- 19. Ольфакторные сигналы. У млекопитающих их передача требует G-белков
- 20. Передача сигнала в обонятельных сенсорных нейронах у разных биол. видов У насекомых “1” (рис.С) G-белков –
- 21. Отличия ольфакторных систем насекомых СХОДСТВО: белки OR насекомых – из 7 трансмембранных доменов, (как и в
- 22. Где расположены обонятельные нейроны у насекомых?
- 23. The Pherobase (http://www.pherobase.net/). Там 2 группы веществ: феромоны (внутривидовые) и “аллелохемики”- межвидовые. Комары и москиты: одорант-аттрактант
- 24. Растение ощущает вредителя по запаху? A. M. Helms, C. M. De Moraes, J. F. Tooker, M.
- 25. Литература Иванов В.Д. Феромоны насекомых //Соросовский образовательный журнал 1998.№6 Саловарова В. П., Приставка А. А., Берсенева
- 27. Скачать презентацию
Слайд 3Первичные мессенджеры - природные экстраклеточные лиганды, которые взаимодействует с рецепторами и активируют
Первичные мессенджеры - природные экстраклеточные лиганды, которые взаимодействует с рецепторами и активируют

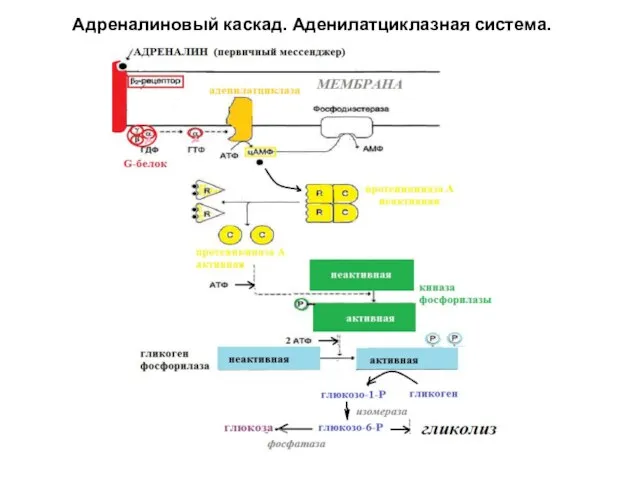
Слайд 4Адреналиновый каскад. Аденилатциклазная система.
Адреналиновый каскад. Аденилатциклазная система.
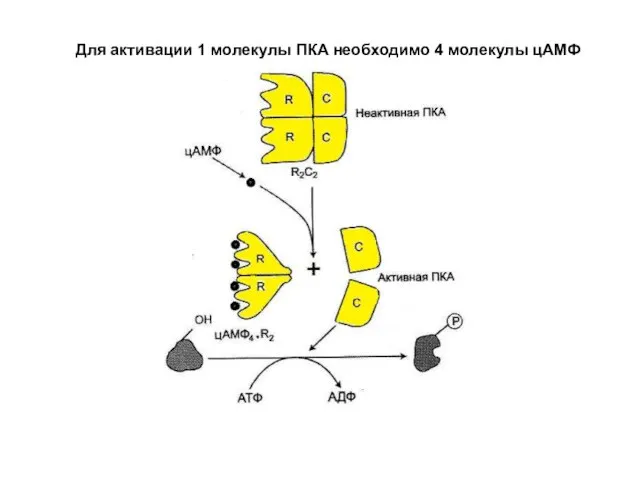
Слайд 5Для активации 1 молекулы ПКА необходимо 4 молекулы цАМФ
Для активации 1 молекулы ПКА необходимо 4 молекулы цАМФ
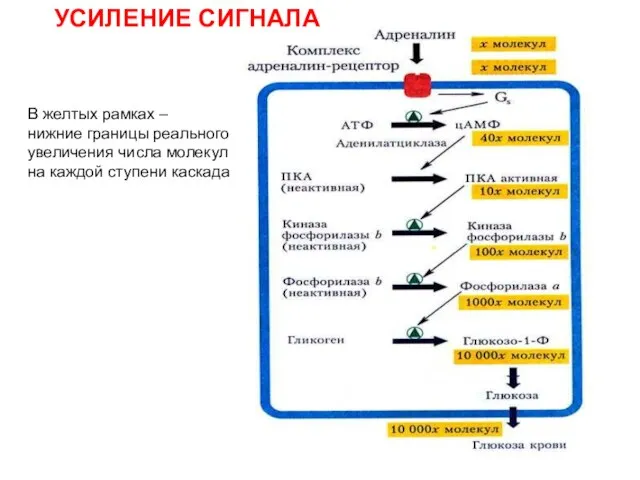
Слайд 6УСИЛЕНИЕ СИГНАЛА
В желтых рамках –
нижние границы реального
увеличения числа молекул
на каждой ступени каскада
УСИЛЕНИЕ СИГНАЛА
В желтых рамках –
нижние границы реального
увеличения числа молекул
на каждой ступени каскада
Слайд 7Как остановить сигнал?
1) Дефосфорилировать (фосфатазы)
2) Устранить цАМФ (фосфодиэстераза)
Как остановить сигнал?
1) Дефосфорилировать (фосфатазы)
2) Устранить цАМФ (фосфодиэстераза)

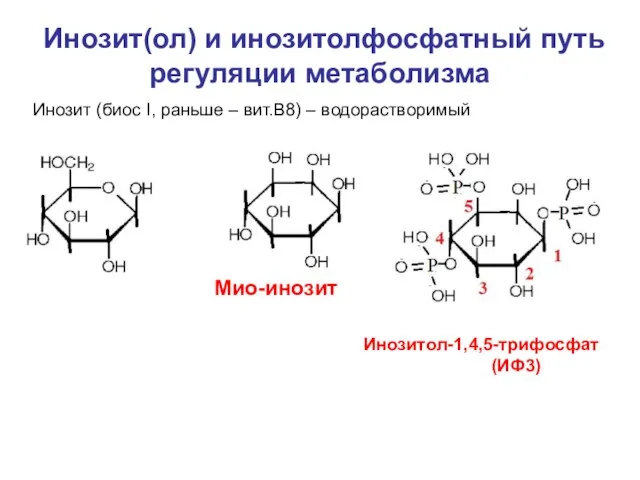
Слайд 8Инозит(ол) и инозитолфосфатный путь регуляции метаболизма
Инозит (биос I, раньше – вит.В8) –
Инозит(ол) и инозитолфосфатный путь регуляции метаболизма
Инозит (биос I, раньше – вит.В8) –
Слайд 9Недостаточность инозита, источники
У человека недостаточность - очень редко.
У животных – плешивость,
нарушения
Недостаточность инозита, источники
У человека недостаточность - очень редко.
У животных – плешивость,
нарушения

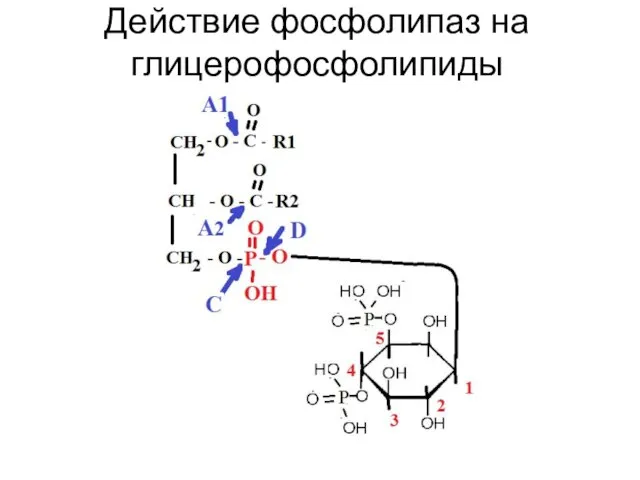
Слайд 10Действие фосфолипаз на глицерофосфолипиды
Действие фосфолипаз на глицерофосфолипиды
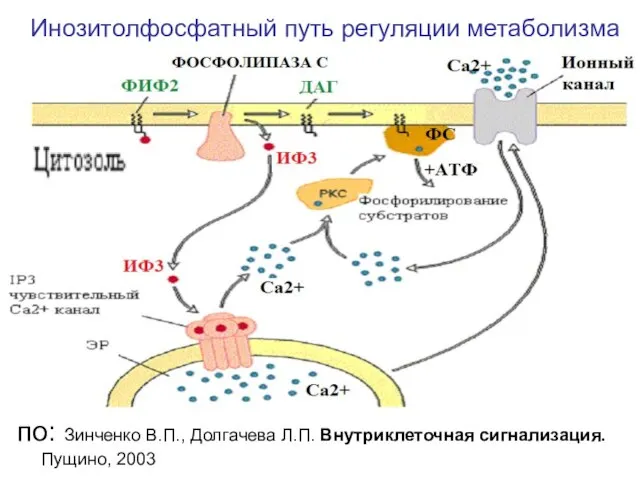
Слайд 11Инозитолфосфатный путь регуляции метаболизма
по: Зинченко В.П., Долгачева Л.П. Внутриклеточная сигнализация. Пущино, 2003
Инозитолфосфатный путь регуляции метаболизма
по: Зинченко В.П., Долгачева Л.П. Внутриклеточная сигнализация. Пущино, 2003
Слайд 12Как остановить сигнал?
1) Дефосфорилировать
2) Откачать Са2+ наружу или обратно в ЭПР (Са2+-АТФаза
Как остановить сигнал?
1) Дефосфорилировать
2) Откачать Са2+ наружу или обратно в ЭПР (Са2+-АТФаза

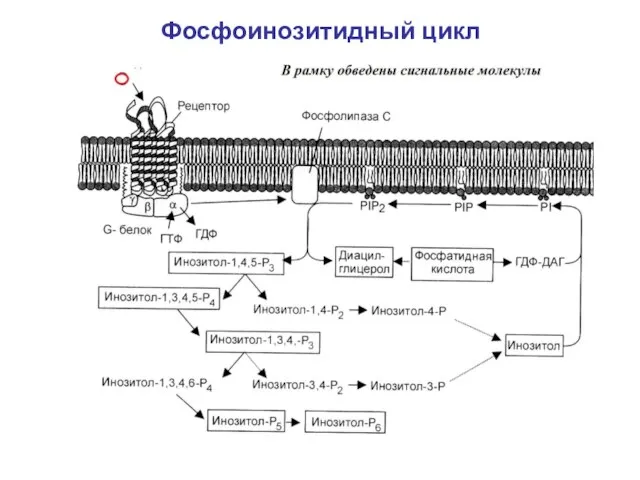
Слайд 13Фосфоинозитидный цикл
Фосфоинозитидный цикл
Слайд 14Нобелевская премия 2004г. - за исследования «обонятельных ре-цепторов и организации системы органов
Нобелевская премия 2004г. - за исследования «обонятельных ре-цепторов и организации системы органов

Слайд 15Ольфакторная (обонятельная) система человека
Человек различает в среднем около 10 000 запахов
Гены,
Ольфакторная (обонятельная) система человека
Человек различает в среднем около 10 000 запахов
Гены,

Слайд 16Большинство наземных млекопитающих имеют в полости носа, кроме основного органа обоняния, ещё
Большинство наземных млекопитающих имеют в полости носа, кроме основного органа обоняния, ещё

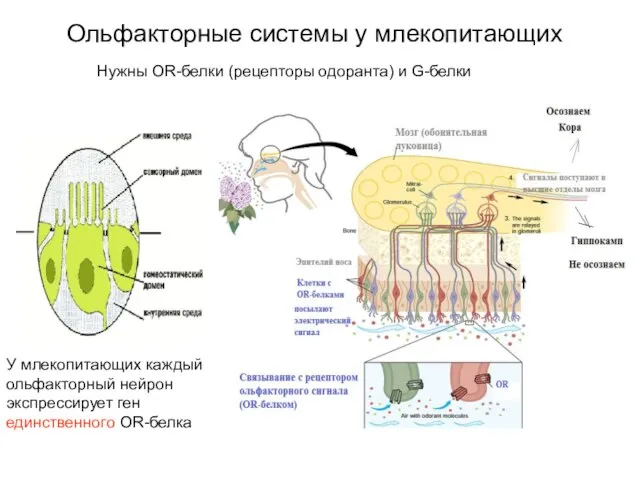
Слайд 17Ольфакторные системы у млекопитающих
У млекопитающих каждый ольфакторный нейрон экспрессирует ген единственного OR-белка
Нужны
Ольфакторные системы у млекопитающих
У млекопитающих каждый ольфакторный нейрон экспрессирует ген единственного OR-белка
Нужны
Слайд 18Потенциал покоя клетки отрицателен (обычно в пределах от –40 до –60 мВ).
Потенциал покоя клетки отрицателен (обычно в пределах от –40 до –60 мВ).

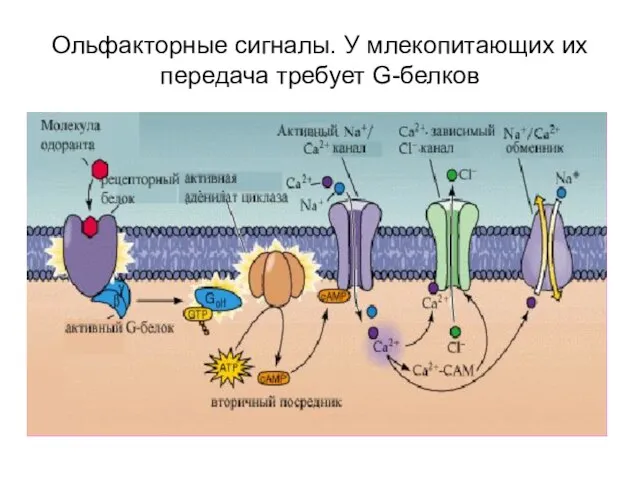
Слайд 19Ольфакторные сигналы. У млекопитающих их передача требует G-белков
Ольфакторные сигналы. У млекопитающих их передача требует G-белков
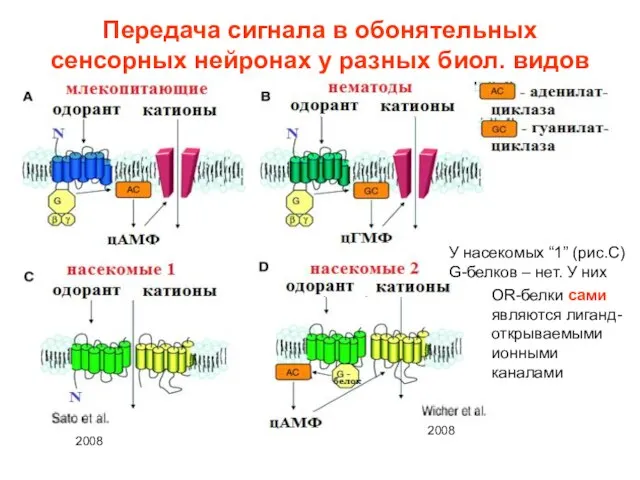
Слайд 20Передача сигнала в обонятельных сенсорных нейронах у разных биол. видов
У насекомых “1”
Передача сигнала в обонятельных сенсорных нейронах у разных биол. видов
У насекомых “1”
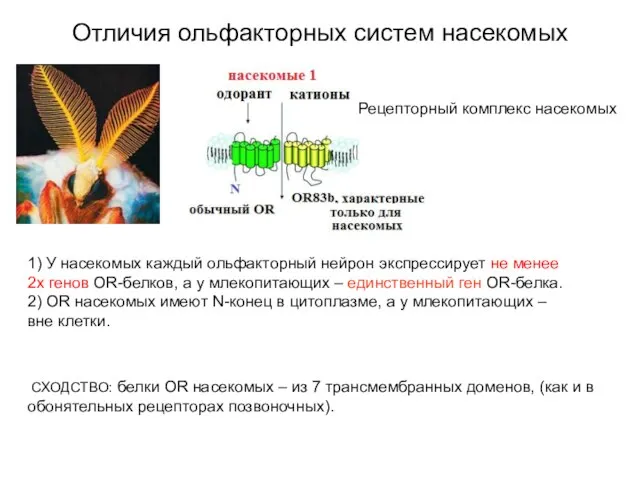
Слайд 21Отличия ольфакторных систем насекомых
СХОДСТВО: белки OR насекомых – из 7
Отличия ольфакторных систем насекомых
СХОДСТВО: белки OR насекомых – из 7
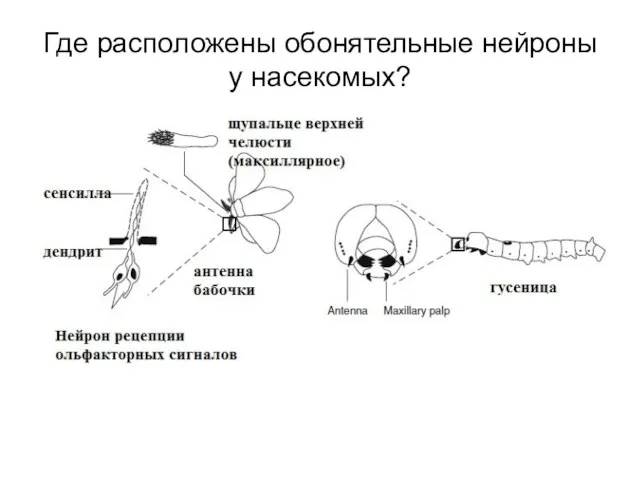
Слайд 22Где расположены обонятельные нейроны у насекомых?
Где расположены обонятельные нейроны у насекомых?
Слайд 23The Pherobase (http://www.pherobase.net/). Там 2 группы веществ: феромоны (внутривидовые) и “аллелохемики”- межвидовые.
The Pherobase (http://www.pherobase.net/). Там 2 группы веществ: феромоны (внутривидовые) и “аллелохемики”- межвидовые.

Слайд 24Растение ощущает
вредителя по запаху?
A. M. Helms, C. M. De Moraes, J. F. Tooker, M. C. Mescher. Exposure of
Растение ощущает вредителя по запаху? A. M. Helms, C. M. De Moraes, J. F. Tooker, M. C. Mescher. Exposure of

Слайд 25Литература
Иванов В.Д. Феромоны насекомых //Соросовский образовательный журнал 1998.№6
Саловарова В. П., Приставка А.
Литература
Иванов В.Д. Феромоны насекомых //Соросовский образовательный журнал 1998.№6
Саловарова В. П., Приставка А.

 Правовая политика Республики Казахстан и ее направления
Правовая политика Республики Казахстан и ее направления Компания Medical Icons
Компания Medical Icons Общее понятие контроля
Общее понятие контроля Формы записи алгоритмов
Формы записи алгоритмов Посадка под школу биохакинга
Посадка под школу биохакинга Автоматизация звука Р в середине слова
Автоматизация звука Р в середине слова В библиотеке Мурманского государственного технического университета с 15.11.2011 по 15.11.2012 открыт доступ к Электронно-библиотечной
В библиотеке Мурманского государственного технического университета с 15.11.2011 по 15.11.2012 открыт доступ к Электронно-библиотечной  Стратегический план развития
Стратегический план развития Правописание окончаний имен существительных 1-ого и 3-го склонения в Р.п., Д.п.
Правописание окончаний имен существительных 1-ого и 3-го склонения в Р.п., Д.п. ПЛАН 2022 2023
ПЛАН 2022 2023 Александр Иванович Куприн 11 класс
Александр Иванович Куприн 11 класс otnositelnost_dvizheniya
otnositelnost_dvizheniya Татьянин день
Татьянин день Производная – это сложно, но прожить без нее не возможно?
Производная – это сложно, но прожить без нее не возможно? Давньогрецький театр та його особливості
Давньогрецький театр та його особливості Современные VPD технологии MSC.Software для университетов
Современные VPD технологии MSC.Software для университетов Excel
Excel Применение Клеточной Хемилюминесценции
Применение Клеточной Хемилюминесценции Economic Justice
Economic Justice Фотоальбом
Фотоальбом Упорство и талант
Упорство и талант Путешествие в Царство Плутона
Путешествие в Царство Плутона простое решение с большими возможностями!
простое решение с большими возможностями! Эскалация_14.10
Эскалация_14.10 Гроза и молния
Гроза и молния Влияние добычи и переработки полезных ископаемых на окружающую среду
Влияние добычи и переработки полезных ископаемых на окружающую среду Алгоритмы канального уровня
Алгоритмы канального уровня Козловская Диана. Грамоты
Козловская Диана. Грамоты