- теломеры

Содержание
- 2. Существование специальных структур на концах хромосом было постулировано в 1938 году классиками генетики, лауреатами Нобелевской премии
- 3. Строение теломер В клетках человека теломеры обычно представлены одноцепочечной ДНК и состоят из несколько тысяч повторяющихся
- 4. Основная функция этих участков — поддержание целостности концов хромосом; Теломера содержит специальные последовательности ДНК, обеспечивающие точную
- 5. "КОНЦЕВАЯ НЕДОРЕПЛИКАЦИЯ ДНК" ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении от ее 3'-конца
- 6. Первым на проблему "концевой недорепликации ДНК" обратил внимание А.М. Оловников в 1971 году. Он высказал гипотезу
- 7. Теломераза Гипотеза А.М. Оловникова нашла убедительное подтверждение в последующие годы. Во-первых, было установлено, что теломеры нормальных
- 8. Механизм действия теломераз Повторное копирование матрицы, включающее этап элонгации, когда дезоксирибонуклеотиды последовательно добавляются к 3'-концу G-богатой
- 9. (1) обратная транскриптаза; (2) теломеразная РНК; (3) нуклеаза, отщепляет от 3'-конца теломерной ДНК один за другим
- 10. Не обходимо отметить, что теломераза синтезирует лишь небольшой участок теломеры, утрачиваемый вследствие концевой репликации. Основная же
- 11. Теломеразная активность Теломераза добавляет особые повторяющиеся последовательности ДНК к 3'-концу цепи ДНК на участках теломер. Длина
- 12. Теломеразная активность соматических клеток В подавляющем большинстве соматических клеток человека на стадии раннего эмбриогенеза происходит выключение
- 13. Теломеразная активность раковых клеток Клетки большинства раковых опухолей характеризуются достаточно высокой активностью теломеразы, которая поддерживает длину
- 14. Вывод Какие же практические выводы следуют из того, что на сегодняшний день удалось узнать о связи
- 16. Скачать презентацию
Слайд 2Существование специальных структур на концах хромосом было постулировано в 1938 году классиками
Существование специальных структур на концах хромосом было постулировано в 1938 году классиками

Слайд 3Строение теломер
В клетках человека теломеры обычно представлены одноцепочечной ДНК и состоят из
Строение теломер
В клетках человека теломеры обычно представлены одноцепочечной ДНК и состоят из

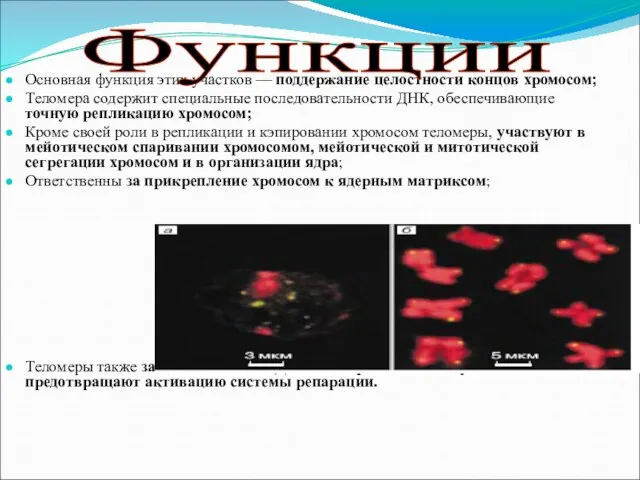
Слайд 4Основная функция этих участков — поддержание целостности концов хромосом;
Теломера содержит специальные последовательности
Основная функция этих участков — поддержание целостности концов хромосом;
Теломера содержит специальные последовательности
Слайд 5
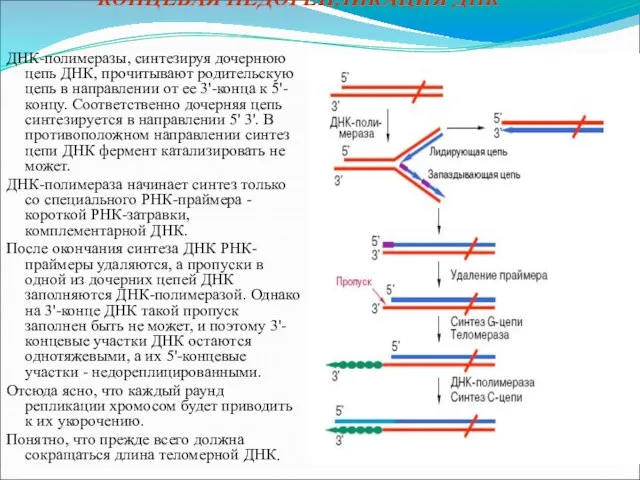
"КОНЦЕВАЯ НЕДОРЕПЛИКАЦИЯ ДНК"
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении
"КОНЦЕВАЯ НЕДОРЕПЛИКАЦИЯ ДНК"
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении
Слайд 6Первым на проблему "концевой недорепликации ДНК" обратил внимание А.М. Оловников в 1971

Слайд 7Теломераза
Гипотеза А.М. Оловникова нашла убедительное подтверждение в последующие годы.
Во-первых, было
Теломераза
Гипотеза А.М. Оловникова нашла убедительное подтверждение в последующие годы.
Во-первых, было

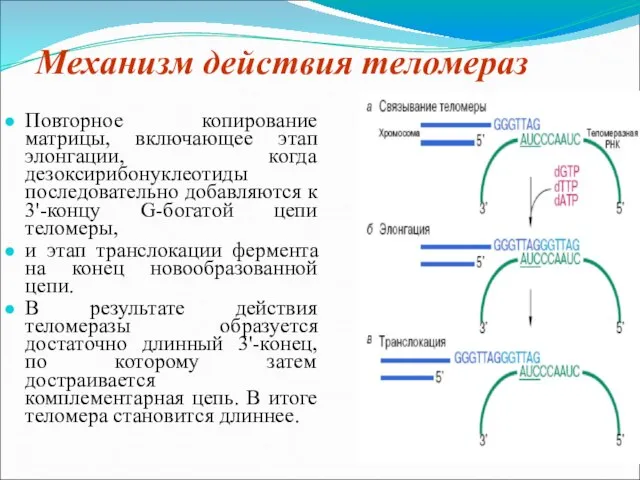
Слайд 8 Механизм действия теломераз
Повторное копирование матрицы, включающее этап элонгации, когда дезоксирибонуклеотиды
Механизм действия теломераз
Повторное копирование матрицы, включающее этап элонгации, когда дезоксирибонуклеотиды
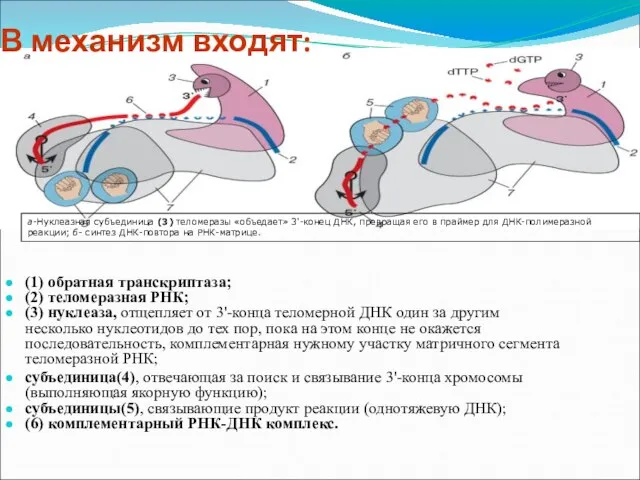
Слайд 9(1) обратная транскриптаза;
(2) теломеразная РНК;
(3) нуклеаза, отщепляет от 3'-конца теломерной ДНК
(1) обратная транскриптаза;
(2) теломеразная РНК;
(3) нуклеаза, отщепляет от 3'-конца теломерной ДНК
Слайд 10 Не обходимо отметить, что теломераза синтезирует лишь небольшой участок теломеры, утрачиваемый
Не обходимо отметить, что теломераза синтезирует лишь небольшой участок теломеры, утрачиваемый

Слайд 11Теломеразная активность
Теломераза добавляет особые повторяющиеся последовательности ДНК к 3'-концу цепи ДНК на
Теломеразная активность
Теломераза добавляет особые повторяющиеся последовательности ДНК к 3'-концу цепи ДНК на

Слайд 12Теломеразная активность соматических клеток
В подавляющем большинстве соматических клеток человека на стадии раннего
Теломеразная активность соматических клеток
В подавляющем большинстве соматических клеток человека на стадии раннего

Слайд 13Теломеразная активность раковых клеток
Клетки большинства раковых опухолей характеризуются достаточно высокой активностью теломеразы,
Теломеразная активность раковых клеток
Клетки большинства раковых опухолей характеризуются достаточно высокой активностью теломеразы,

Слайд 14Вывод
Какие же практические выводы следуют из того, что на сегодняшний день удалось
Вывод
Какие же практические выводы следуют из того, что на сегодняшний день удалось

 Леонардо да Винчи (8 класс)
Леонардо да Винчи (8 класс) Let’s Learn About Russia
Let’s Learn About Russia 03.09.20122 Берестов Валентин Дмитриевич родился 1 апреля 1928 года в с. Мещовск, Калужской области. По образованию археолог, окончил истфа
03.09.20122 Берестов Валентин Дмитриевич родился 1 апреля 1928 года в с. Мещовск, Калужской области. По образованию археолог, окончил истфа Презентация на тему ТИПЫ БЕЗРАБОТИЦЫ Экономика 11 класс
Презентация на тему ТИПЫ БЕЗРАБОТИЦЫ Экономика 11 класс  Организация и управление юридической деятельностью
Организация и управление юридической деятельностью Притяжение Земли
Притяжение Земли Научное мероприятие
Научное мероприятие Химия и окружающая среда
Химия и окружающая среда Отчет о проведении недели истории
Отчет о проведении недели истории Рождество в России
Рождество в России Атмосферные фронты
Атмосферные фронты Понятие и предмет маркетингового анализа
Понятие и предмет маркетингового анализа Натуральные растительные волокна
Натуральные растительные волокна Презентация на тему Основы безопасности жизнедеятельности 7 класс
Презентация на тему Основы безопасности жизнедеятельности 7 класс  Части речи 3 класс
Части речи 3 класс Физические и химические явления. Химические уравнения
Физические и химические явления. Химические уравнения Булочка с творогом
Булочка с творогом ТЕМА: «Сложение и вычитание десятичных дробей»
ТЕМА: «Сложение и вычитание десятичных дробей» Пуск дизеля тепловоза ТЭМ-14
Пуск дизеля тепловоза ТЭМ-14 Презентация на тему "Чем люди живы" 11 класс
Презентация на тему "Чем люди живы" 11 класс ИСПОЛЬЗОВАНИЕ VISUAL BASIC FOR APPLICATIONS В ИНТЕРАКТИВНЫХ ПЕДАГОГИЧЕСКИХ МУЛЬТИМЕДИЙНЫХ МАСТЕР-ШАБЛОНАХ
ИСПОЛЬЗОВАНИЕ VISUAL BASIC FOR APPLICATIONS В ИНТЕРАКТИВНЫХ ПЕДАГОГИЧЕСКИХ МУЛЬТИМЕДИЙНЫХ МАСТЕР-ШАБЛОНАХ Индивид. Психологические теории личности
Индивид. Психологические теории личности Ознакомление детей старшего дошкольного возраста с декоративно-прикладным искусством Узбекистана
Ознакомление детей старшего дошкольного возраста с декоративно-прикладным искусством Узбекистана История масленицы
История масленицы Реестр должностей государственной службы
Реестр должностей государственной службы Гигиена девушки. Урок технологии
Гигиена девушки. Урок технологии Подвиги Геракла
Подвиги Геракла Экопоселения. Шаблон
Экопоселения. Шаблон